Sphingomyelin Deacylase, the Enzyme Involved in the Pathogenesis of Atopic Dermatitis, Is Identical to the β-Subunit of Acid Ceramidase


Abstract
:1. Introduction
2. Results
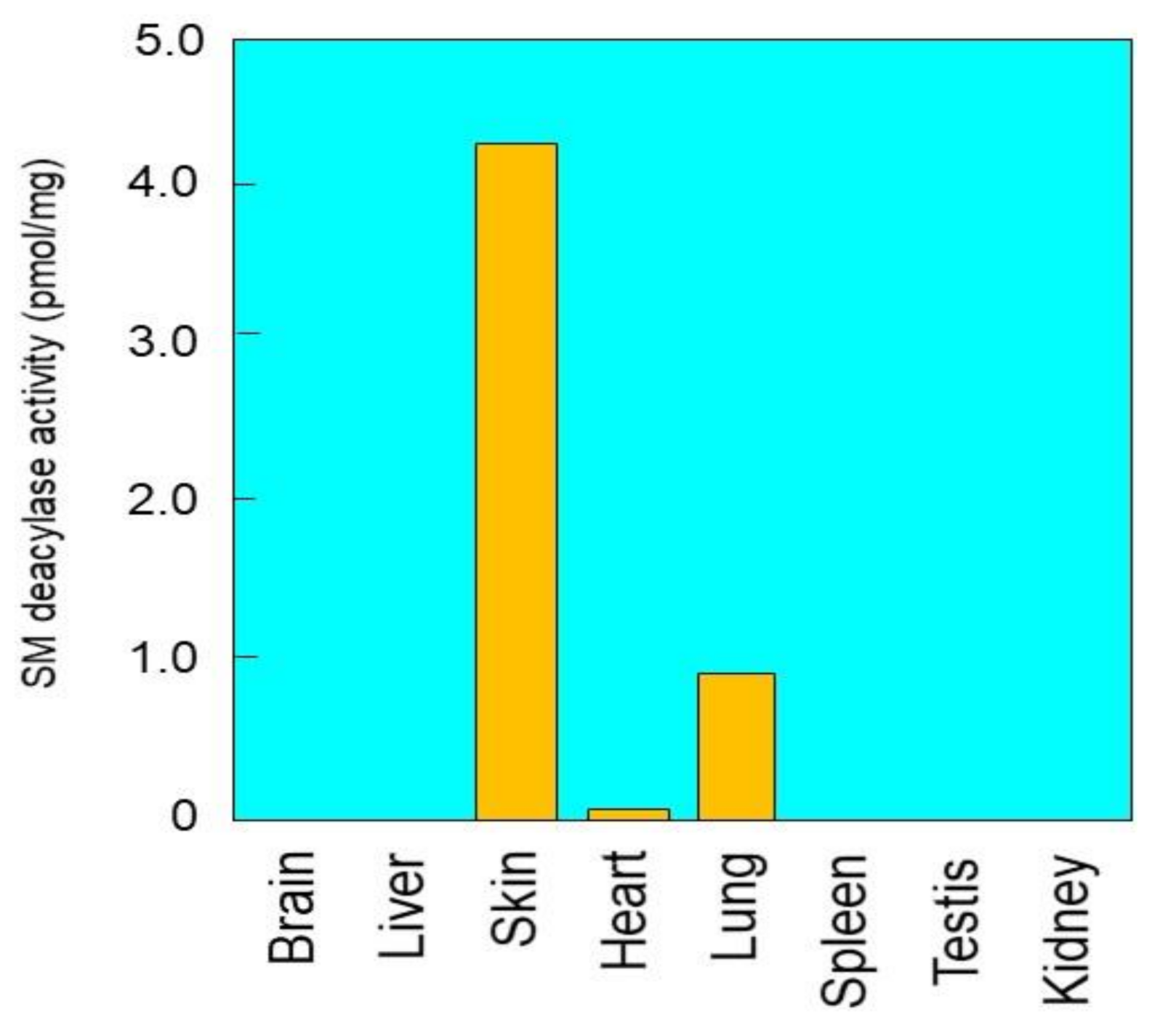
2.1. Tissue Distribution of SM Deacylase Activity
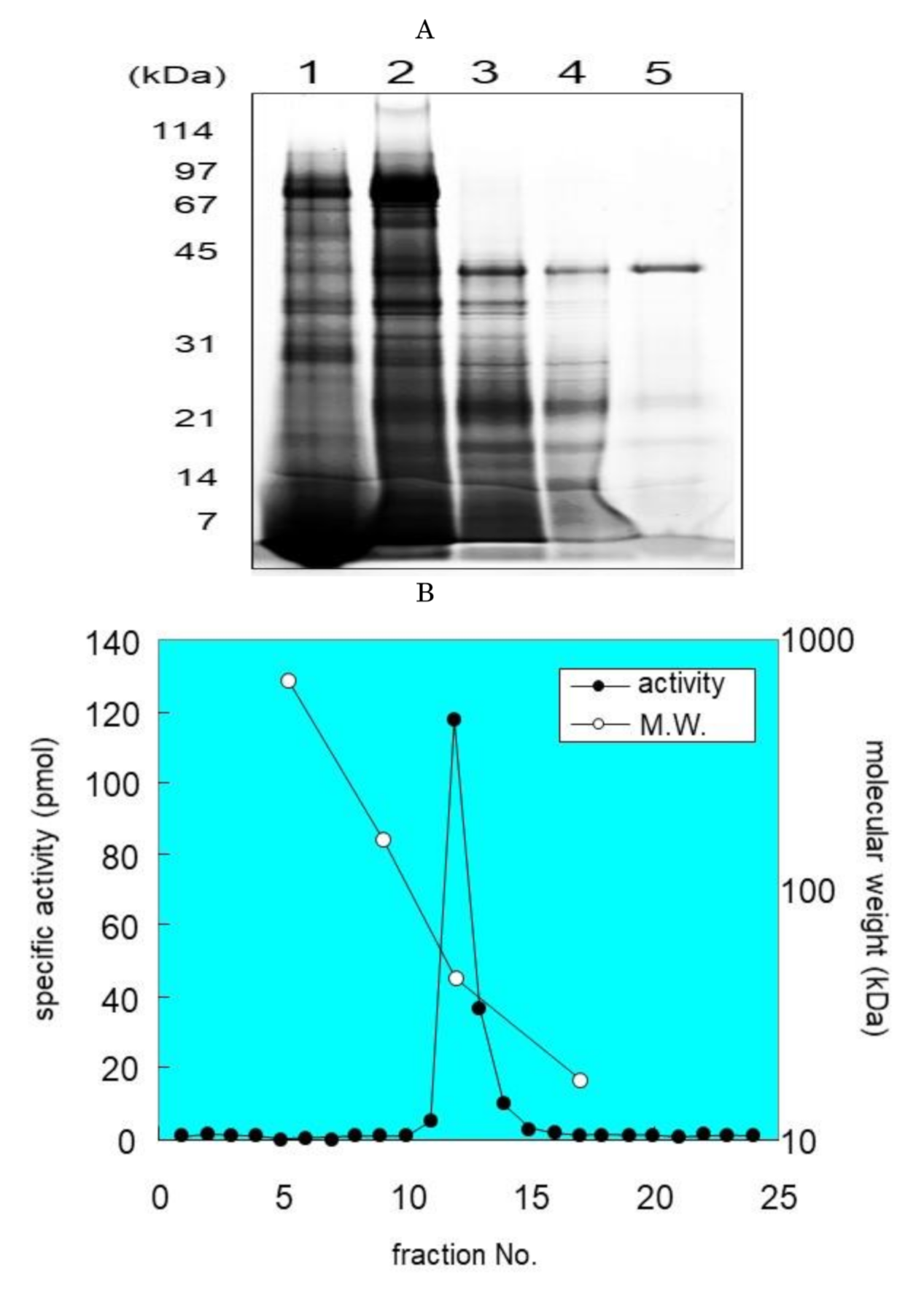
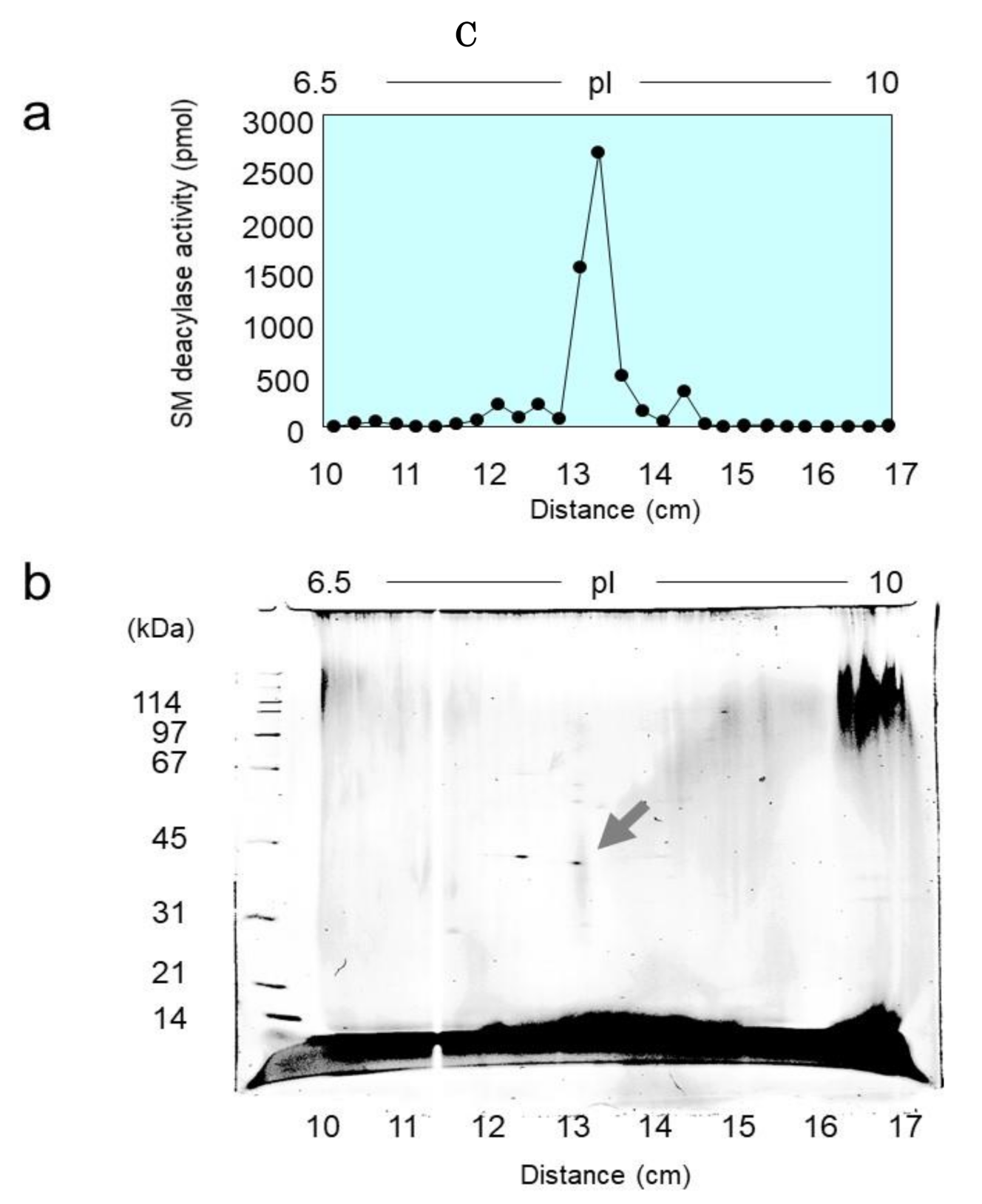
2.2. Purification of SM Deacylase from Rat Skin
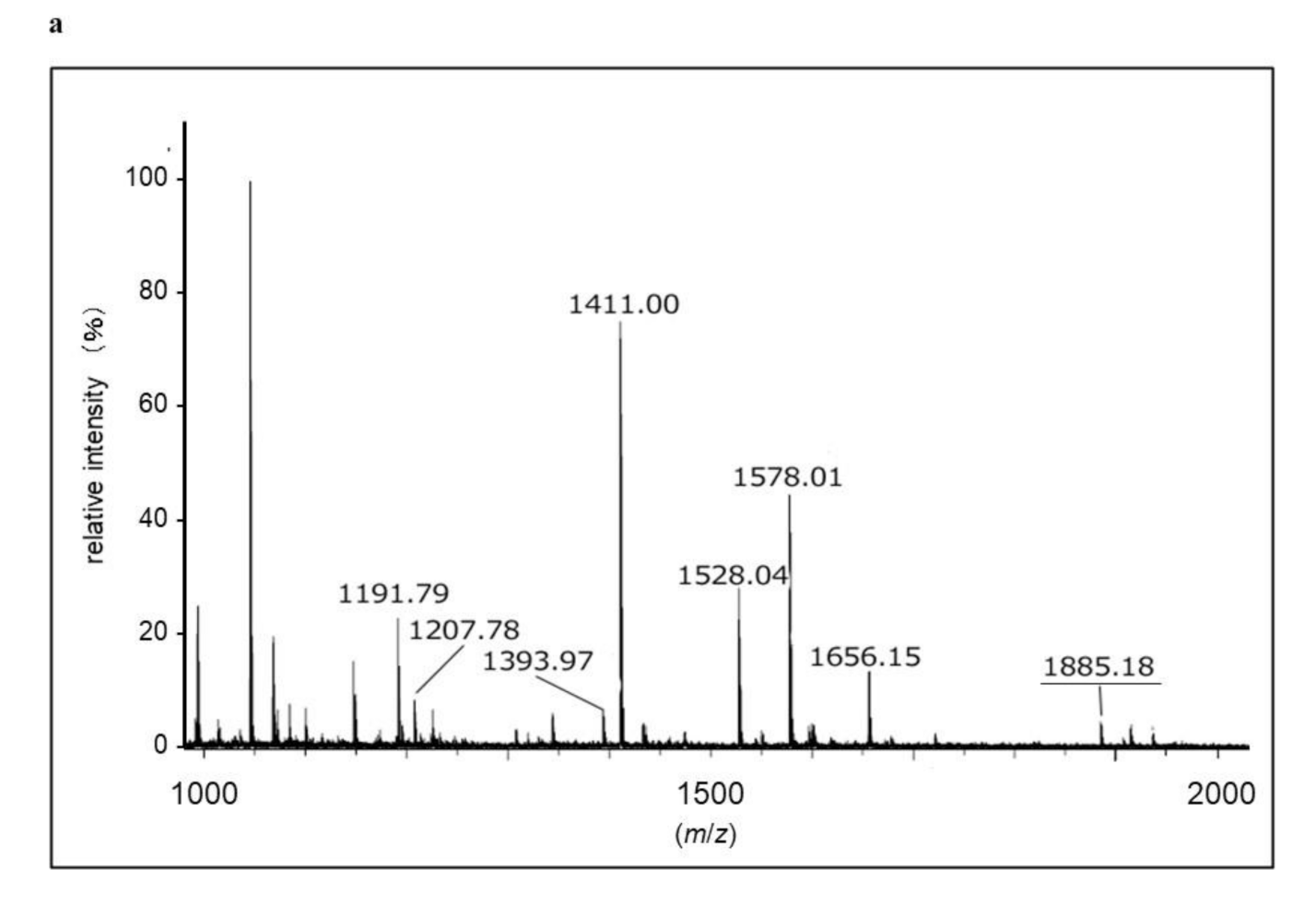
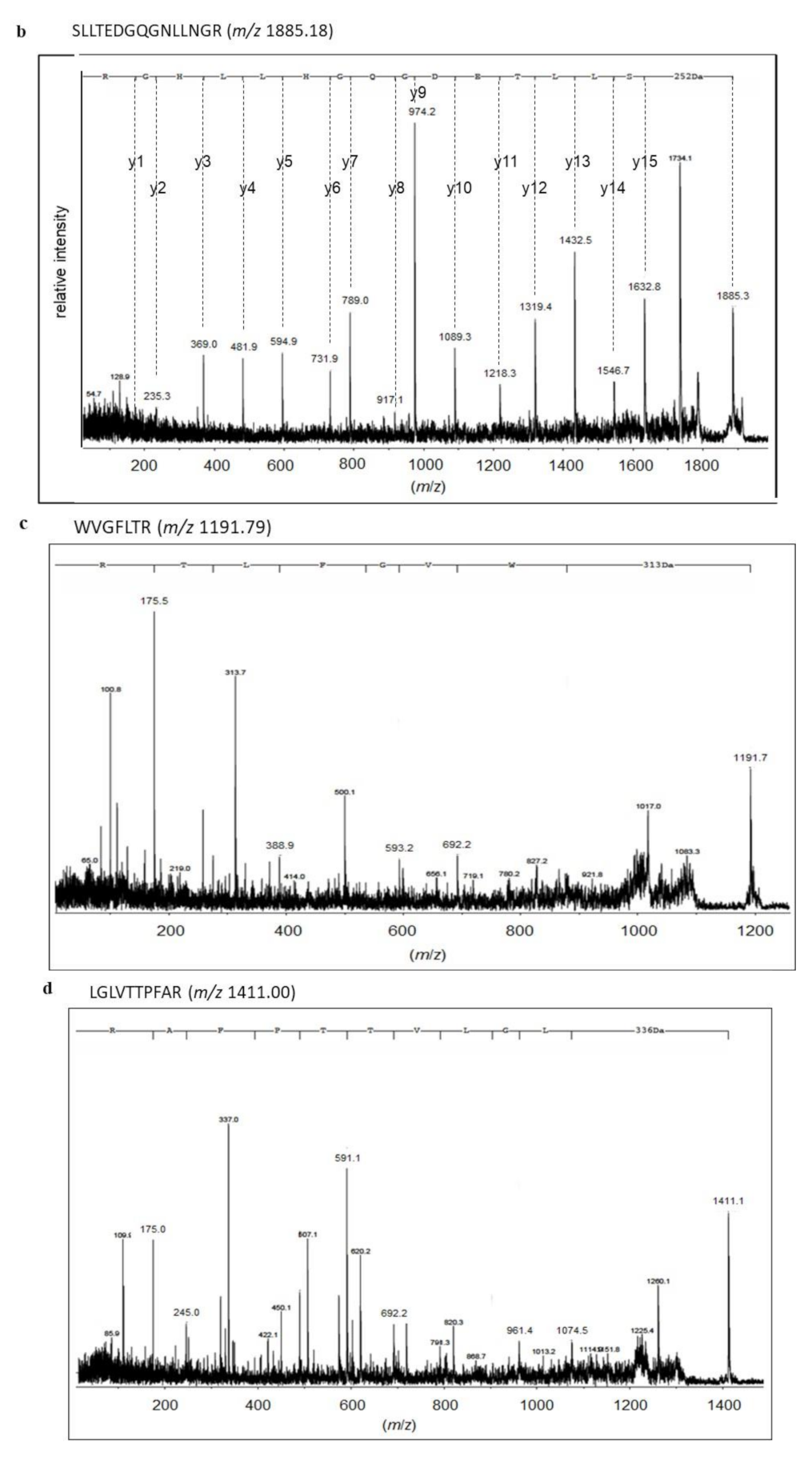
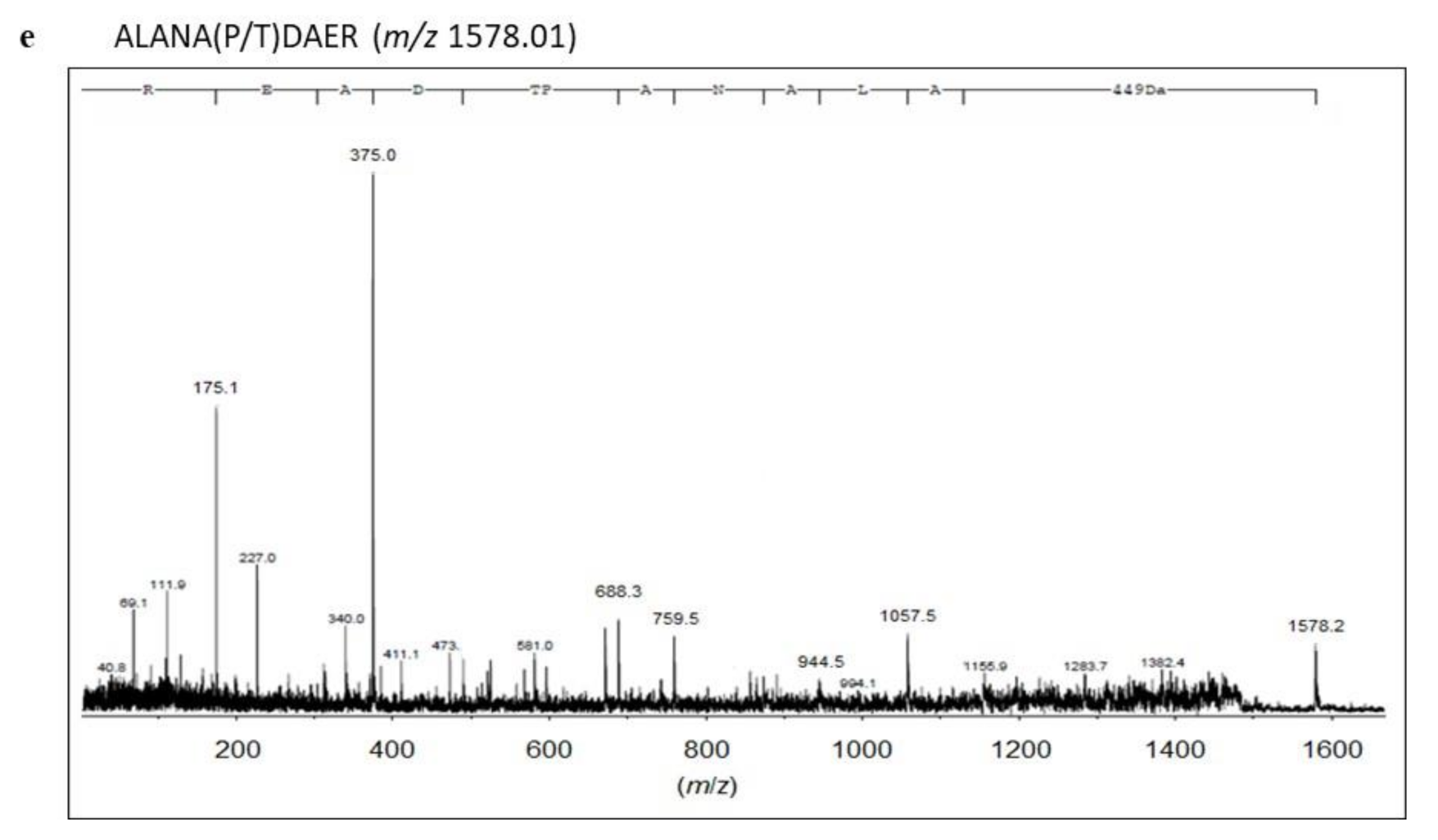
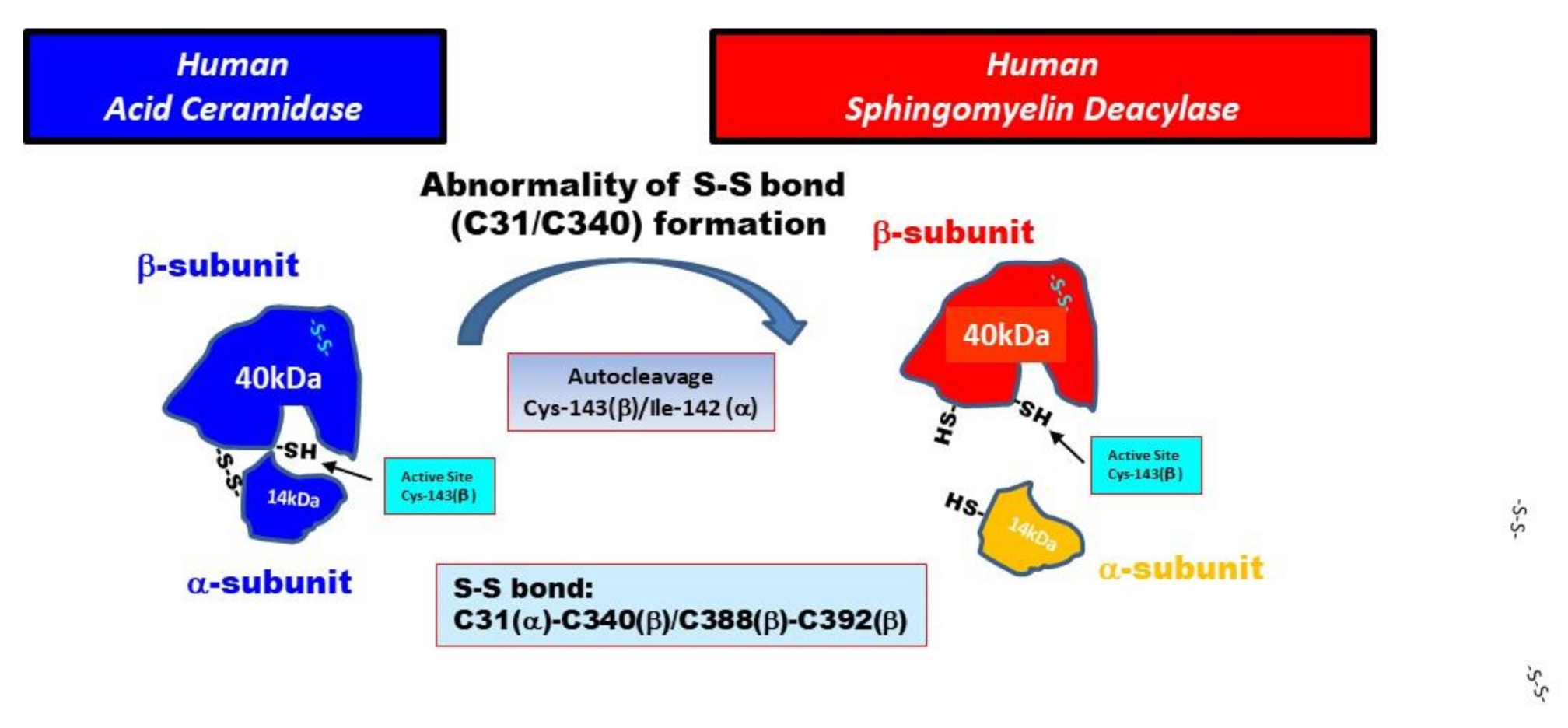
2.3. Identification of Proteins with SM Deacylase Activity
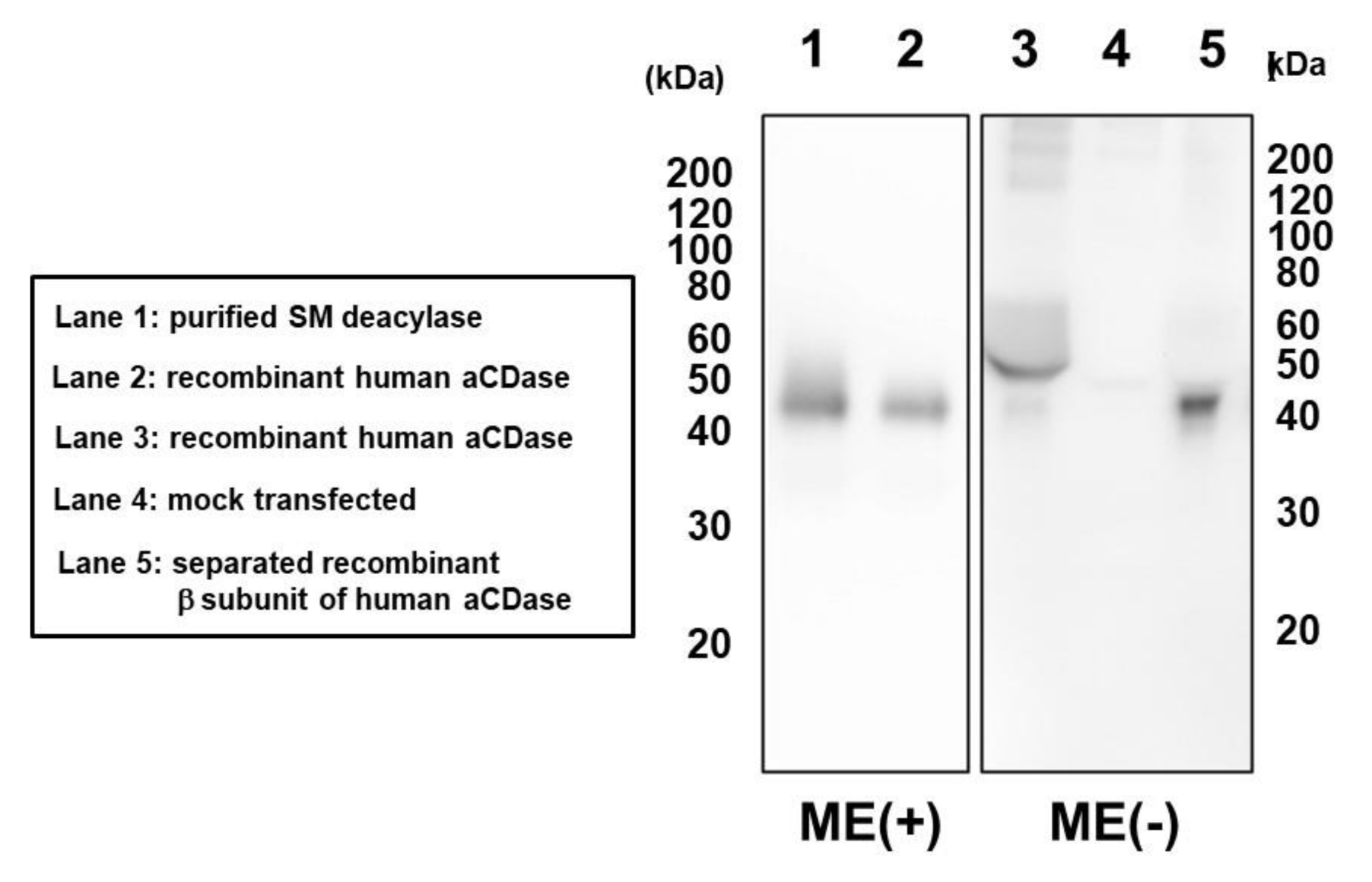
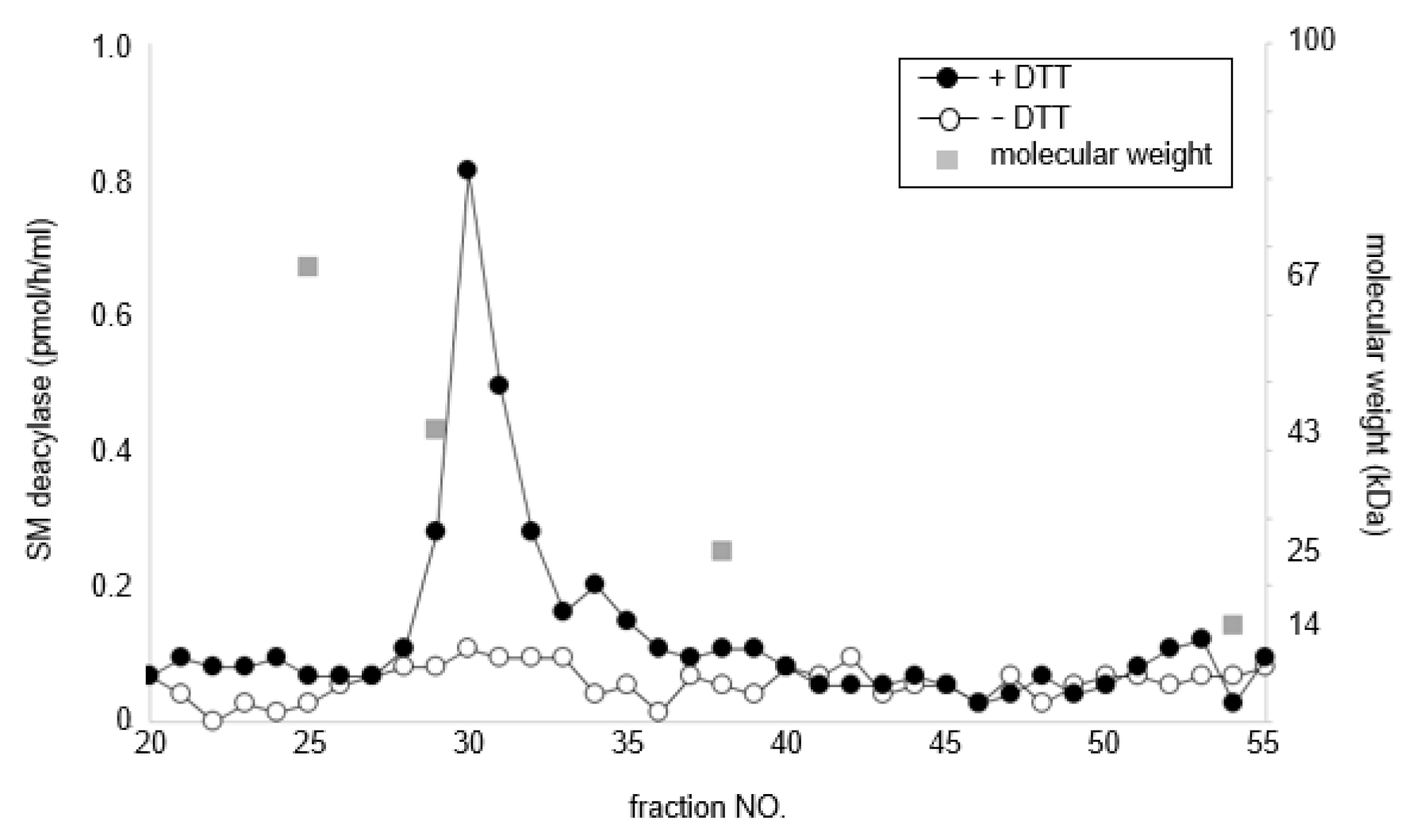
2.4. Treatment with DTT Separates SM Deacylase from Recombinant Human aCDase
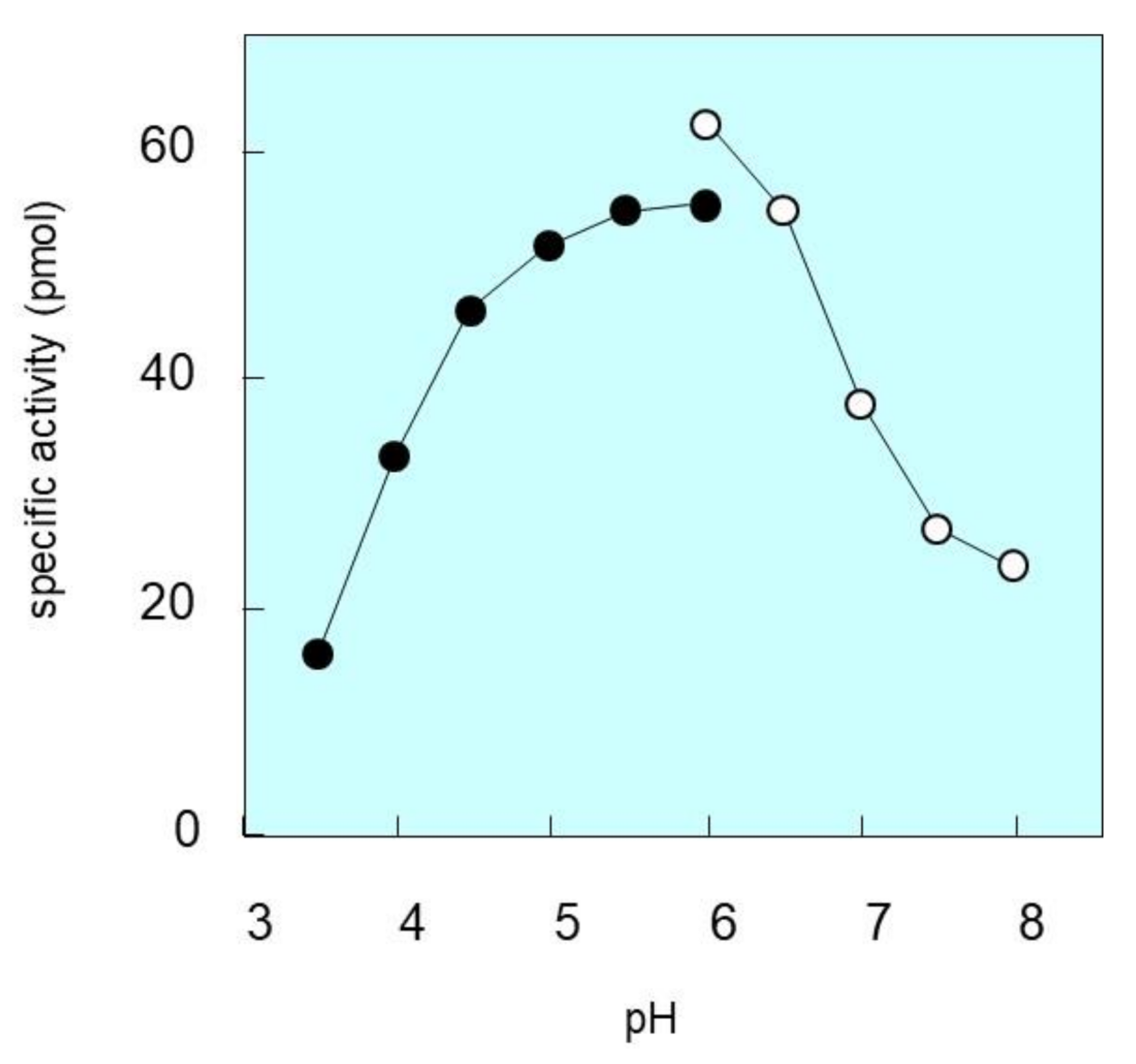
2.5. pH Dependence of SM Deacylase Activity
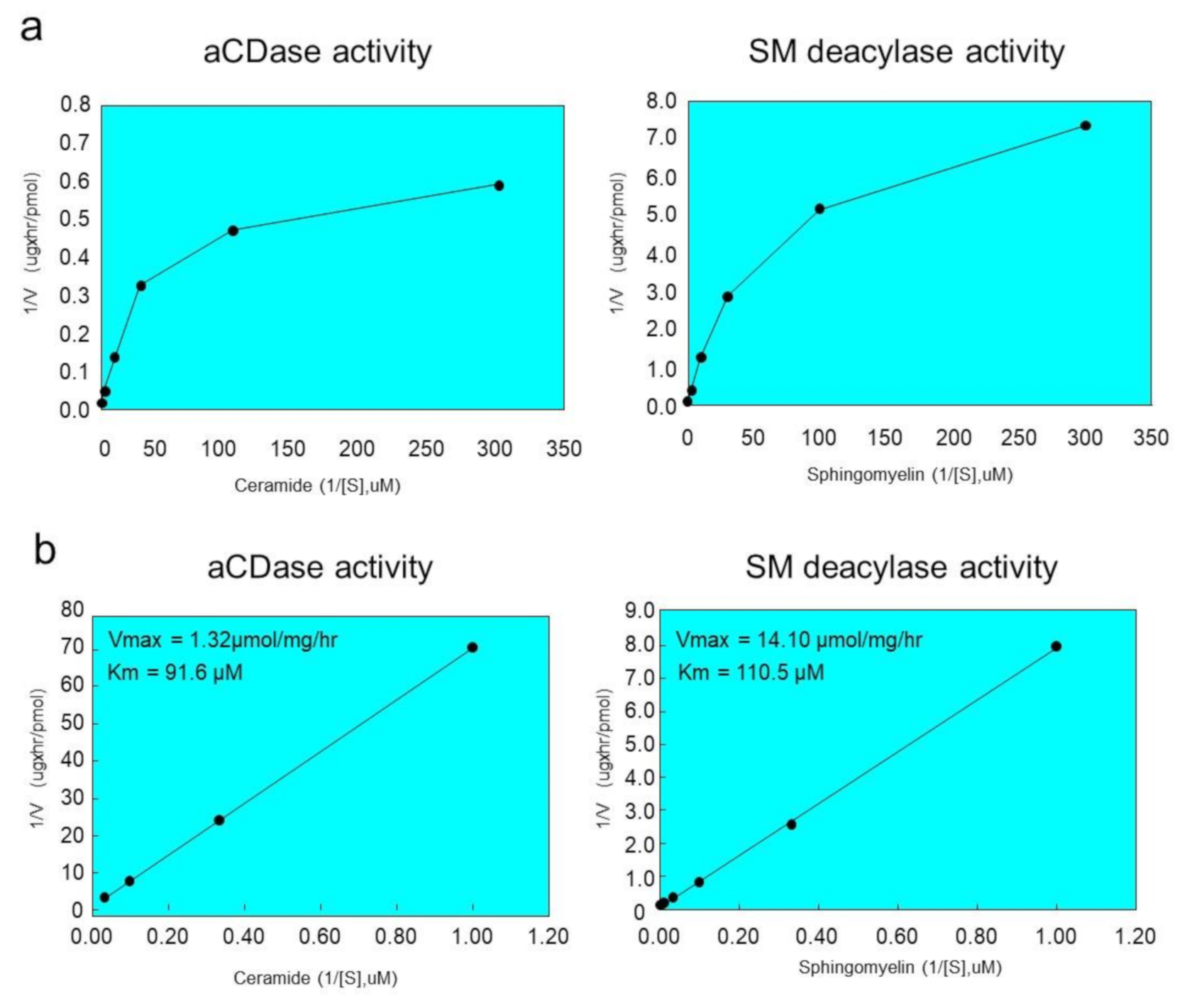
2.6. Ceramide and SM Hydrolyzing Activity of Purified SM Deacylase
3. Discussion
4. Materials and Methods
4.1. Materials and Antibodies
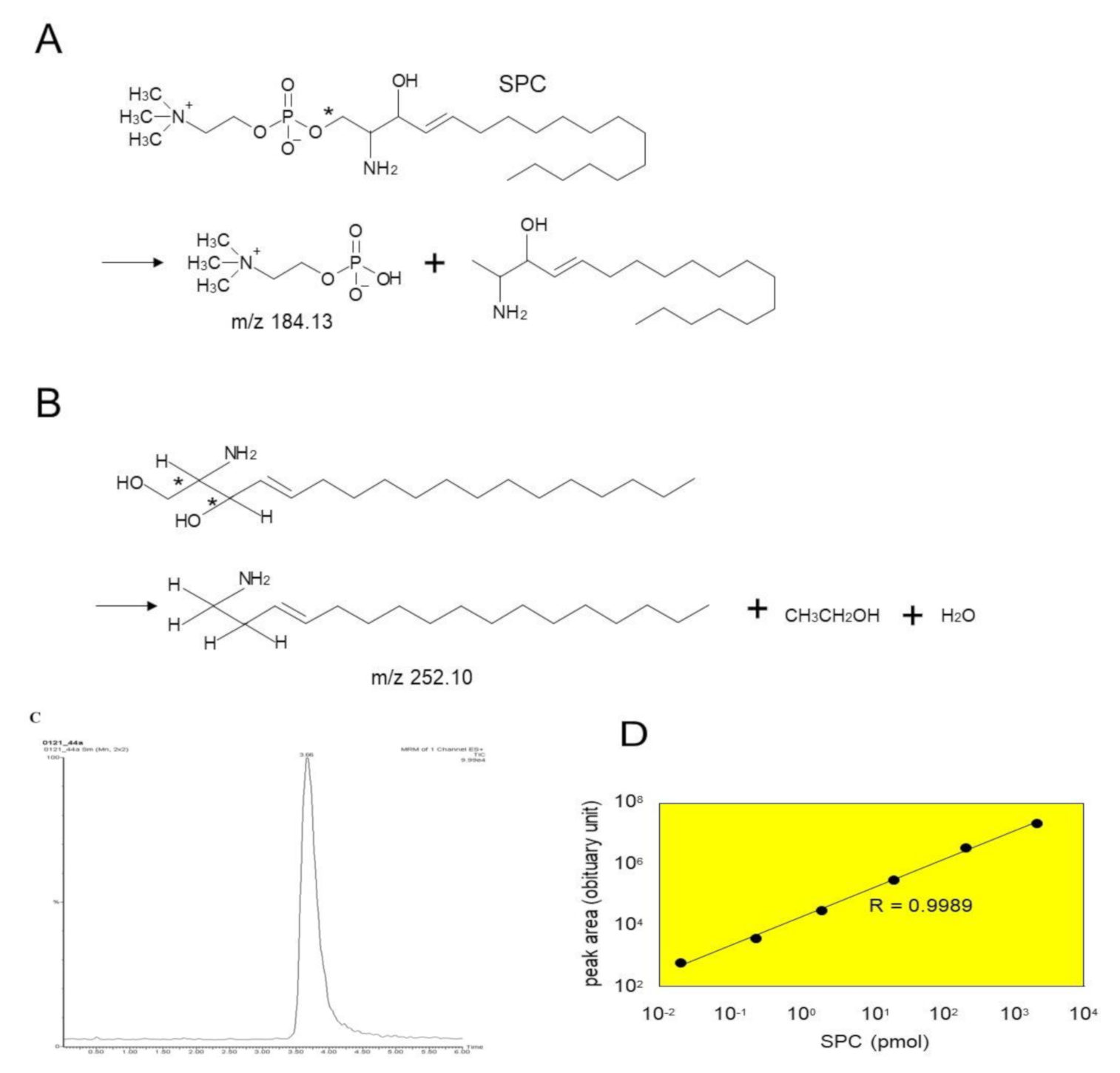
4.2. Assays for SM Deacylase and aCDase Activities
4.3. Purification of SM deacylase
4.3.1. Preparation of Skin Extracts from Rat Skin
4.3.2. Purification of Skin Extracts
Isoelectric Focusing
Ion Exchange Chromatography
Gel Filtration Chromatography
Chromatography with a C20 Column (Similar to Affinity Chromatography)
4.4. In-gel Protein Digestion and Protein Identification by MALDI-TOF-MS
4.5. Expression and Preparation of Recombinant Human aCDase
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | atopic dermatitis |
| aCDase | acid ceramidase |
| SM | sphingomyelin |
| SPC | sphingosylphosphorylcholine |
| PC | phosphorylcholine |
| SPH | sphingosine |
| IEF | isoelectric focusing |
| SC | stratum corneum |
| pCer | synthetic pseudo-ceramide |
| DTT | dithiothreitol |
| BGCase | β-glucocerebrosidase |
| aSMase | acid sphingomyelinase |
| LGs | lamellar granules |
| GCer | glucosylceramide |
| SPT | serine-palmitoyl transferase |
| SCD | stearoyl CoA desaturase |
| GCERS | GCer synthase |
| SMS | GCer synthase |
| GS | glucosylsphingosine |
| HCs | healthy controls |
References
- Hata, M.; Tokura, Y.; Takigawa, M.; Sato, M.; Shioya, Y.; Fujikura, Y.; Imokawa, G. Assessment of epidermal barrier function by photoacoustic spectrometry in relation to its importance in the pathogenesis of atopic dermatitis. Lab. Investig. 2002, 82, 1451–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imokawa, G.; Ishida, K. Role of ceramide in the barrier function of the stratum corneum, Implications for the pathogenesis of atopic dermatitis. J. Clin. Exp. Dermatol. Res. 2014, 5, 206–218. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased level of ceramides in stratum corneum of atopic dermatitis: An etiologic factor in atopic dry skin? J. Investig. Dermatol. 1991, 96, 523–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Nardo, A.; Wertz, P.; Giannetti, A.; Seidenari, S. Ceramide and cholesterol composition of the skin of patients with atopic dermatitis. Acta Derm. Venereol. 1998, 78, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Arikawa, J.; Ishibashi, M.; Kawashima, M.; Takagi, Y.; Ichikawa, Y.; Imokawa, G. Decreased levels of sphingosine, a natural antimicrobial agent, may be associated with vulnerability of the stratum corneum from patients with atopic dermatitis to colonization by Staphylococcus aureus. J. Investig. Dermatol. 2002, 119, 433–439. [Google Scholar] [CrossRef]
- Ishibashi, M.; Arikawa, J.; Okamoto, R.; Kawashima, M.; Takagi, Y.; Ohguchi, K.; Imokawa, G. Abnormal expression of the novel epidermal enzyme, glucosylceramide deacylase, and the accumulation of its enzymatic reaction product, glucosylsphingosine, in the skin of patients with atopic dermatitis. Lab. Investig. 2003, 83, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, R.; Arikawa, J.; Ishibashi, M.; Kawashima, M.; Takagi, Y.; Imokawa, G. Sphingosylphosphorylcholine is upregulated in the stratum corneum of patients with atopic dermatitis. J. Lipid Res. 2003, 44, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, J.; Narita, H.; Kondo, N.; Hotta, M.; Takagi, Y.; Masukawa, Y.; Kitahara, T.; Takema, Y.; Koyano, S.; Yamazaki, S.; et al. Changes in the ceramide profile of atopic dermatitis patients. J. Investig. Dermatol. 2010, 130, 2511–2514. [Google Scholar] [CrossRef] [Green Version]
- Jungersted, J.M.; Scheer, H.; Mempel, M.; Baurecht, H.; Cifuentes, L.; Høgh, J.K.; Hellgren, L.I.; Jemec, G.B.E.; Agner, T.; Weidinger, S. Stratum corneum lipids, skin barrier function and filaggrin mutations in patients with atopic eczema. Allergy 2010, 65, 911–918. [Google Scholar] [CrossRef]
- Sugiura, A.; Nomura, T.; Mizuno, A.; Imokawa, G. Reevaluation of the non-lesional dry skin in atopic dermatitis by acute barrier disruption: An abnormal permeability barrier homeostasis with defective processing to generate ceramide. Arch. Dermatol. Res. 2014, 306, 427–440. [Google Scholar] [CrossRef]
- Imokawa, G.; Kuno, H.; Kawai, M. Stratum corneum lipids serve as a bound-water modulator. J. Investig. Dermatol. 1991, 96, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Holleran, W.M.; Feingold, K.R.; Man, M.Q.; Gao, W.N.; Lee, J.M.; Elias, P.M. Regulation of epidermal sphingolipid synthesis by permeability barrier function. J. Lipid Res. 1991, 32, 1151–1158. [Google Scholar] [PubMed]
- Holleran, W.M.; Man, M.Q.; Gao, W.N.; Menon, G.K.; Elias, P.M.; Feingold, K.R. Sphingolipids are required for mammalian epidermal barrier function. Inhibition of sphingolipid synthesis delays barrier recovery after acute perturbation. J. Clin. Investig. 1991, 88, 1338–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holleran, W.M.; Takagi, Y.; Menon, G.K.; Legler, G.; Feingold, K.R.; Elias, P.M. Processing of epidermal glucosylceramides is required for optimal mammalian cutaneous permeability barrier function. J. Clin. Investig. 1993, 91, 1656–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holleran, W.M.; Ginns, E.I.; Menon, G.K.; Grundmann, J.U.; Fartasch, M.; McKinney, C.E.; Elias, P.M.; Sidransky, E. Consequences of beta-glucocerebrosidase deficiency in epidermis. Ultrastructure and permeability barrier alterations in Gaucher disease. J. Clin. Investig. 1994, 93, 1756–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holleran, W.M.; Takagi, Y.; Menon, G.K.; Jackson, S.M.; Lee, J.M.; Feingold, K.R.; Elias, P.M. Permeability barrier requirements regulate epidermal beta-glucocerebrosidase. J. Lipid Res. 1994, 35, 905–912. [Google Scholar] [PubMed]
- Janssens, M.; van Smeden, J.; Gooris, G.S.; Bras, W.; Portale, G.; Caspers, P.J.; Vreeken, R.J.; Kezic, S.; Lavrijsen, A.P.; Bouwstra, J.A. Lamellar lipid organization and ceramide composition in the stratum corneum of patients with atopic eczema. J. Investig. Dermatol. 2011, 131, 2136–2138. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Takahashi, A.; Bito, K.; Draelos, Z.; Imokawa, G. Treatment with Synthetic Pseudoceramide Improves Atopic Skin, Switching the Ceramide Profile to a Healthy Skin Phenotype. J. Investig. Dermatol. 2020, 140, 1762–1770. [Google Scholar] [CrossRef]
- Higuchi, K.; Hara, J.; Okamoto, R.; Kawashima, M.; Imokawa, G. The skin of atopic dermatitis patients contains a novel enzyme, glucosylceramide sphingomyelin deacylase, which cleaves the N-acyl linkage of sphingomyelin and glucosylceramide. Biochem. J. 2000, 350 Pt 3, 747–756. [Google Scholar] [CrossRef]
- Jin, K.; Higaki, Y.; Takagi, Y.; Higuchi, K.; Yada, Y.; Kawashima, M.; Imokawa, G. Analysis of beta-glucocerebrosidase and ceramidase activities in atopic and aged dry skin. Acta Derm. Venereol. 1994, 74, 337–340. [Google Scholar]
- Kusuda, S.; Cui, C.Y.; Takahashi, M.; Tezuka, T. Localization of sphingomyelinase in lesional skin of atopic dermatitis patients. J. Investig. Dermatol. 1998, 111, 733–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danso, M.; Boiten, W.; Van Drongelen, V.; Meijling, K.G.; Gooris, G.; El Ghalbzouri, A.; Absalah, S.; Vreeken, R.; Kezic, S.; Van Smeden, J.; et al. Altered expression of epidermal lipid bio-synthesis enzymes in atopic dermatitis skin is accompanied by changes in stratum corneum lipid composition. J. Dermatol. Sci. 2017, 88, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, J.M.; Forl, M.; Winoto-Morbach, S.; Seite, S.; Schunck, M.; Proksch, E.; Schutze, S. Acid and neutral sphingomyelinase, ceramide synthase, and acid ceramidase activities in cutaneous aging. Exp. Dermatol. 2005, 14, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Ogata, J.; Higaki, Y.; Kawashima, M.; Yada, Y.; Higuchi, K.; Tsuchiya, T.; Kawainami, S.; Imokawa, G. Abnormal expression of sphingomyelin acylase in atopic dermatitis: An etiologic factor for ceramide deficiency? J. Investig. Dermatol. 1996, 106, 1242–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, J.; Higuchi, K.; Okamoto, R.; Kawashima, M.; Imokawa, G. High-expression of sphingomyelin deacylase is an important determinant of ceramide deficiency leading to barrier disruption in atopic dermatitis. J. Investig. Dermatol. 2000, 115, 406–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imokawa, G. A possible mechanism underlying the ceramide deficiency in atopic dermatitis: Expression of a deacylase enzyme that cleaves the N-acyl linkage of sphingomyelin and glucosylceramide. J. Dermatol. Sci. 2009, 55, 1–9. [Google Scholar] [CrossRef]
- Higuchi, K.; Kawashima, M.; Ichikawa, Y.; Imokawa, G. Sphingosylphosphorylcholine is a Melanogenic Stimulator for Human Melanocytes. Pigment. Cell Res. 2003, 16, 670–678. [Google Scholar] [CrossRef]
- Imokawa, G.; Takagi, Y.; Higuchi, K.; Kondo, H.; Yada, Y. Sphingosylphosphorylcholine is a potent inducer of intercellular adhesion molecule-1 expression in human keratinocytes. J. Investig. Dermatol. 1999, 112, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, K.; Kawashima, M.; Takagi, Y.; Kondo, H.; Yada, Y.; Ichikawa, Y.; Imokawa, G. Sphingosylphosphorylcholine is an activator of transglutaminase activity in human keratinocytes. J. Lipid Res. 2001, 42, 1562–1570. [Google Scholar]
- Colver, G.B.; Mortimer, P.S.; Millard, P.R.; Dawber, R.P.R.; Ryan, T.J. The ‘Dirty Neck’-a reticulate pigmentation in atopics. Clin. Exp. Dermatol. 1987, 12, 1–4. [Google Scholar] [CrossRef]
- He, X.; Okino, N.; Dhami, R.; Dagan, A.; Gatt, S.; Schulze, H.; Sandhoff, K.; Schuchman, E.H. Purification and characterization of recombinant, human acid ceramidase. Catalytic reactions and interactions with acid sphingomyelinase. J. Biol. Chem. 2003, 278, 32978–32986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardo, K.; Hurwitz, R.; Zenk, T.; Desnick, R.J.; Ferlinz, K.; Schuchman, E.H.; Sandhoff, K. Purification, characterization, and biosynthesis of human acid ceramidase. J. Biol. Chem. 1995, 270, 11098–11102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linke, T.; Wilkening, G.; Sadeghlar, F.; Mozcall, H.; Bernardo, K.; Schuchman, E.; Sandhoff, K. Interfacial regulation of acid ceramidase activity. Stimulation of ceramide degradation by lysosomal lipids and sphingolipid activator proteins. J. Biol. Chem. 2001, 276, 5760–5768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, J.; Gartner, S.; Li, C.M.; Quintern, L.E.; Bernardo, K.; Levran, O.; Schnabel, D.; Desnick, R.J.; Schuchman, E.H.; Sandhoff, K. Molecular cloning and characterization of a full-length complementary DNA encoding human acid ceramidase. Identification Of the first molecular lesion causing Farber disease. J. Biol. Chem. 1996, 271, 33110–33115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferlinz, K.; Kopal, G.; Bernardo, K.; Linke, T.; Bar, J.; Breiden, B.; Neumann, U.; Lang, F.; Schuchman, E.H.; Sandhoff, K. Human acid ceramidase: Processing, glycosylation, and lysosomal targeting. J. Biol. Chem. 2001, 276, 35352–35360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebai, A.; Gorelik, A.; Li, Z.; Illes, K.; Nagar, B. Structural basis for the activation of acid ceramidase. Nat. Commun. 2018, 9, 1621. [Google Scholar] [CrossRef]
- Schulze, H.; Schepers, U.; Sandhoff, K. Overexpression and mass spectrometry analysis of mature human acid ceramidase. Biol. Chem. 2007, 388, 1333–1343. [Google Scholar] [CrossRef]
- Oizumi, A.; Nakayama, H.; Okino, N.; Iwahara, C.; Kina, K.; Matsumoto, R.; Ogawa, H.; Takamori, K.; Ito, M.; Suga, Y.; et al. Pseudomonas-derived ceramidase induces production of inflammatory mediators from human keratinocytes via sphingosine-1-phosphate. PLoS ONE 2014, 9, e89402. [Google Scholar] [CrossRef] [Green Version]
- Shtraizent, N.; Eliyahu, E.; Park, J.H.; He, X.; Shalgi, R.; Schuchman, E.H. Autoproteolytic cleavage and activation of human acid ceramidase. J. Biol. Chem. 2008, 283, 11253–11259. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total Protein | Total Activity | Specific Activity | Enrichment | Yield |
|---|---|---|---|---|---|
| mg | pmol | pmol/mg | fold | % | |
| homogenate | 2447 | 6.3 × 104 | 26 | 1 | 100 |
| Phenyl-5PW | 293 | 2.2 × 104 | 76 | 3 | 35 |
| Rotofor | 55 | 8.8 × 105 | 16089 | 622 | 1398 |
| SP-Sepharose | 27 | 7.5 × 105 | 27735 | 1072 | 1183 |
| Superdex 200 | 2 | 7.6 × 105 | 363919 | 14067 | 1207 |
| Shodex RP18-415 | - | 2.4 × 105 | - | - | 379 |

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teranishi, Y.; Kuwahara, H.; Ueda, M.; Takemura, T.; Kusumoto, M.; Nakamura, K.; Sakai, J.; Kimura, T.; Furutani, Y.; Kawashima, M.; et al. Sphingomyelin Deacylase, the Enzyme Involved in the Pathogenesis of Atopic Dermatitis, Is Identical to the β-Subunit of Acid Ceramidase. Int. J. Mol. Sci. 2020, 21, 8789. https://doi.org/10.3390/ijms21228789
Teranishi Y, Kuwahara H, Ueda M, Takemura T, Kusumoto M, Nakamura K, Sakai J, Kimura T, Furutani Y, Kawashima M, et al. Sphingomyelin Deacylase, the Enzyme Involved in the Pathogenesis of Atopic Dermatitis, Is Identical to the β-Subunit of Acid Ceramidase. International Journal of Molecular Sciences. 2020; 21(22):8789. https://doi.org/10.3390/ijms21228789
Chicago/Turabian StyleTeranishi, Yasuhiro, Hiroshi Kuwahara, Masaru Ueda, Tadashi Takemura, Masanori Kusumoto, Keiji Nakamura, Jun Sakai, Toru Kimura, Yasuji Furutani, Makoto Kawashima, and et al. 2020. "Sphingomyelin Deacylase, the Enzyme Involved in the Pathogenesis of Atopic Dermatitis, Is Identical to the β-Subunit of Acid Ceramidase" International Journal of Molecular Sciences 21, no. 22: 8789. https://doi.org/10.3390/ijms21228789
APA StyleTeranishi, Y., Kuwahara, H., Ueda, M., Takemura, T., Kusumoto, M., Nakamura, K., Sakai, J., Kimura, T., Furutani, Y., Kawashima, M., Imokawa, G., & Nogami-Itoh, M. (2020). Sphingomyelin Deacylase, the Enzyme Involved in the Pathogenesis of Atopic Dermatitis, Is Identical to the β-Subunit of Acid Ceramidase. International Journal of Molecular Sciences, 21(22), 8789. https://doi.org/10.3390/ijms21228789