
Cutting Edge of the Pathogenesis of Atopic Dermatitis: Sphingomyelin Deacylase, the Enzyme Involved in Its Ceramide Deficiency, Plays a Pivotal Role


Abstract
:1. Skin Characteristics and Barrier/Water Reservoir Functions in the Stratum Corneum of Patients with Atopic Dermatitis
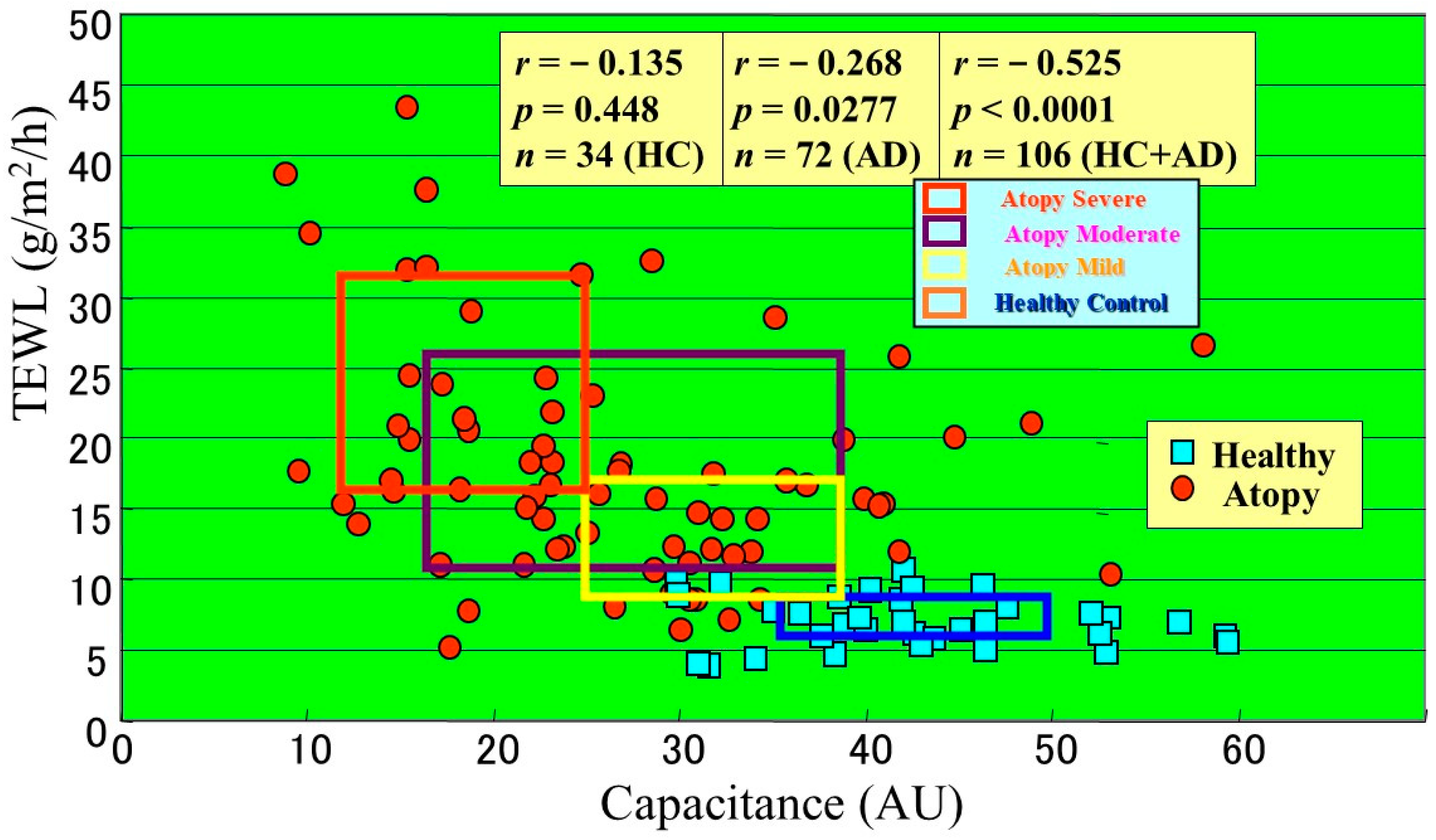
2. Abnormality in Percutaneous Permeability Barrier Function
3. Is Barrier Disruption a Cause or a Result of Dermatitis?
4. A Ceramide Deficiency Is Responsible for the Disrupted SC Function
5. Significance of Ceramides in SC Functions
6. Significance of the Ceramide Profile in SC Functions
7. Is the Barrier Disruption and Its Associated Ceramide Deficiency Inherent or Not?
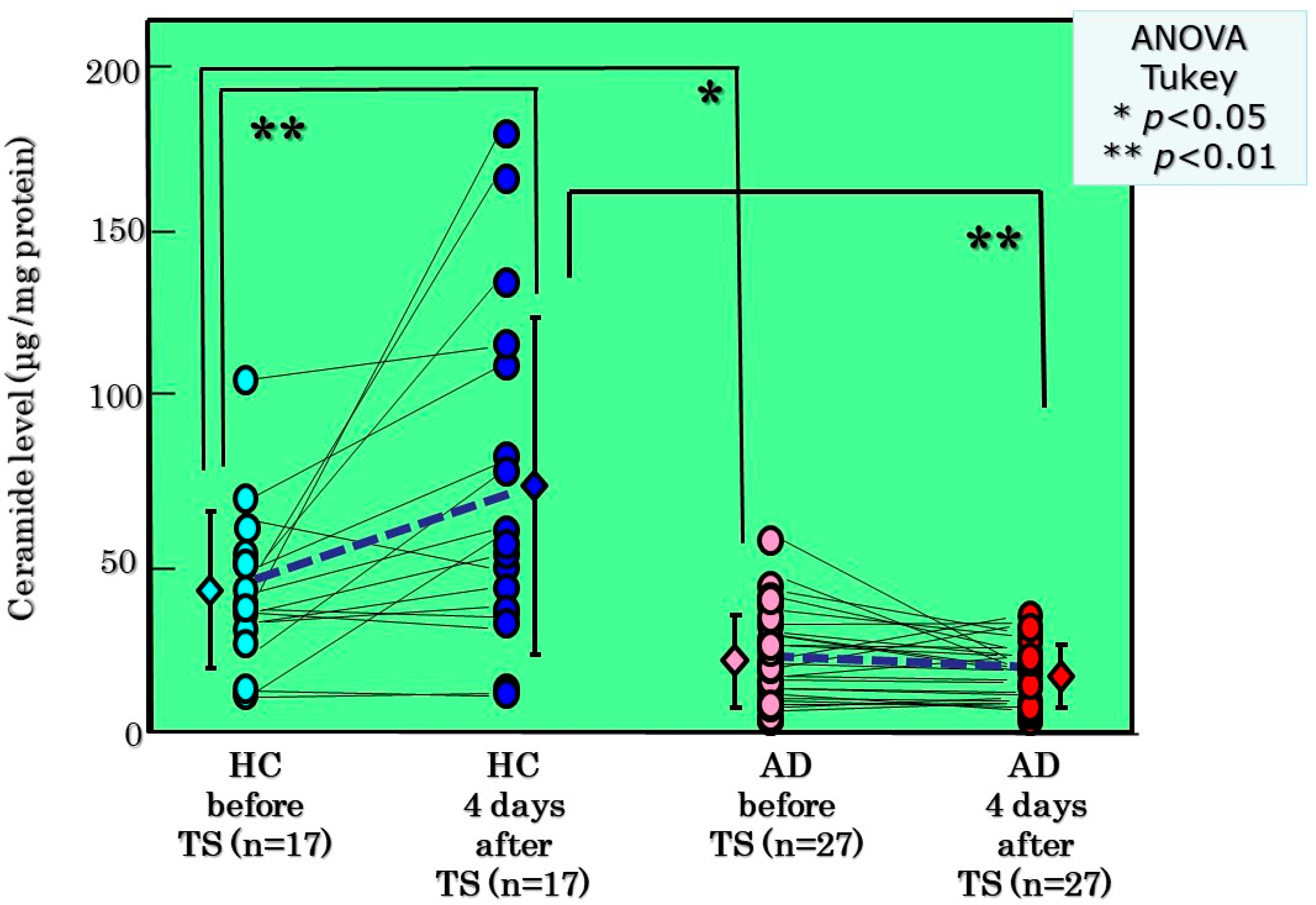
8. Clinical Evidence for the Impaired Homeostasis of the Ceramide-Generating Process
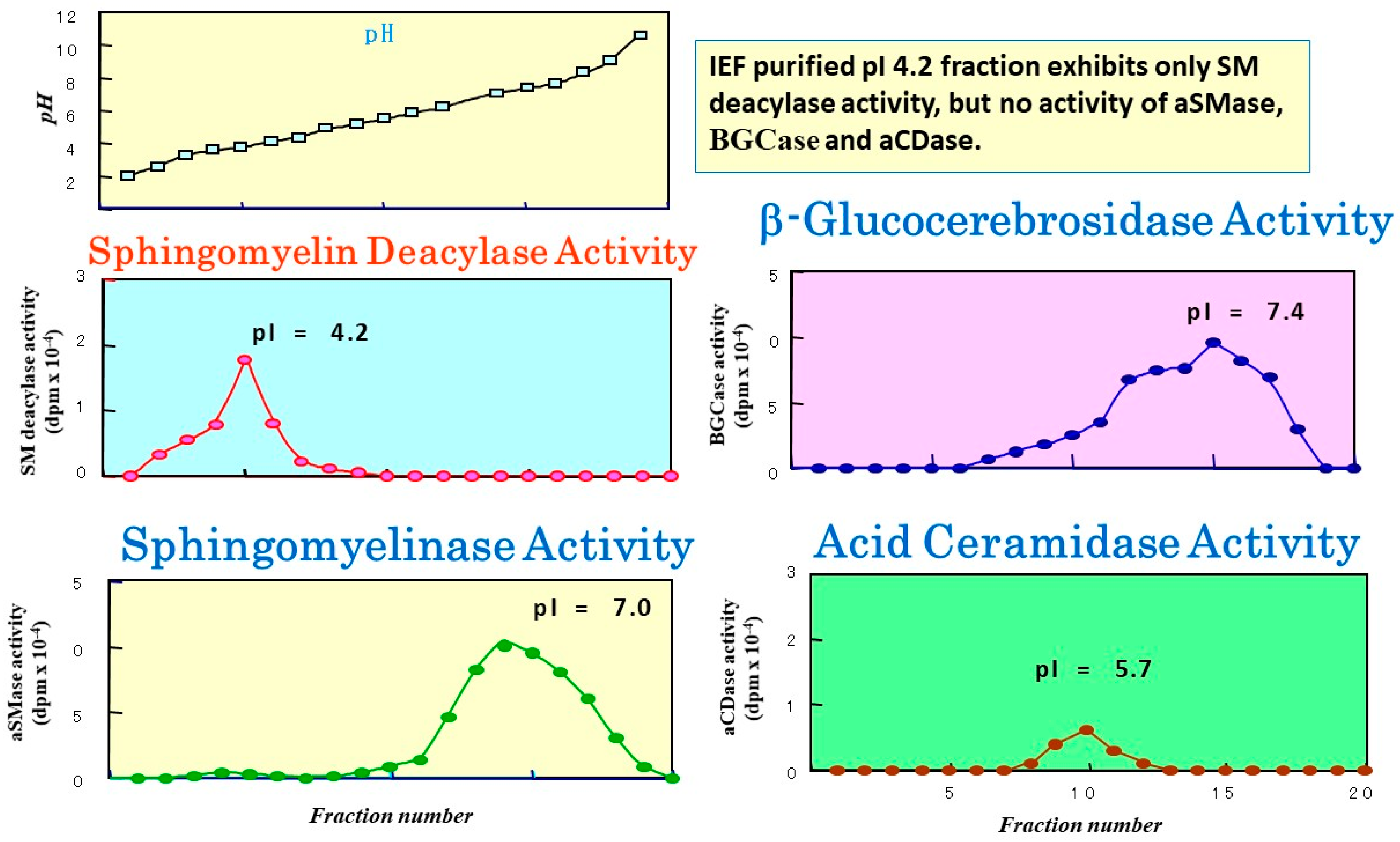
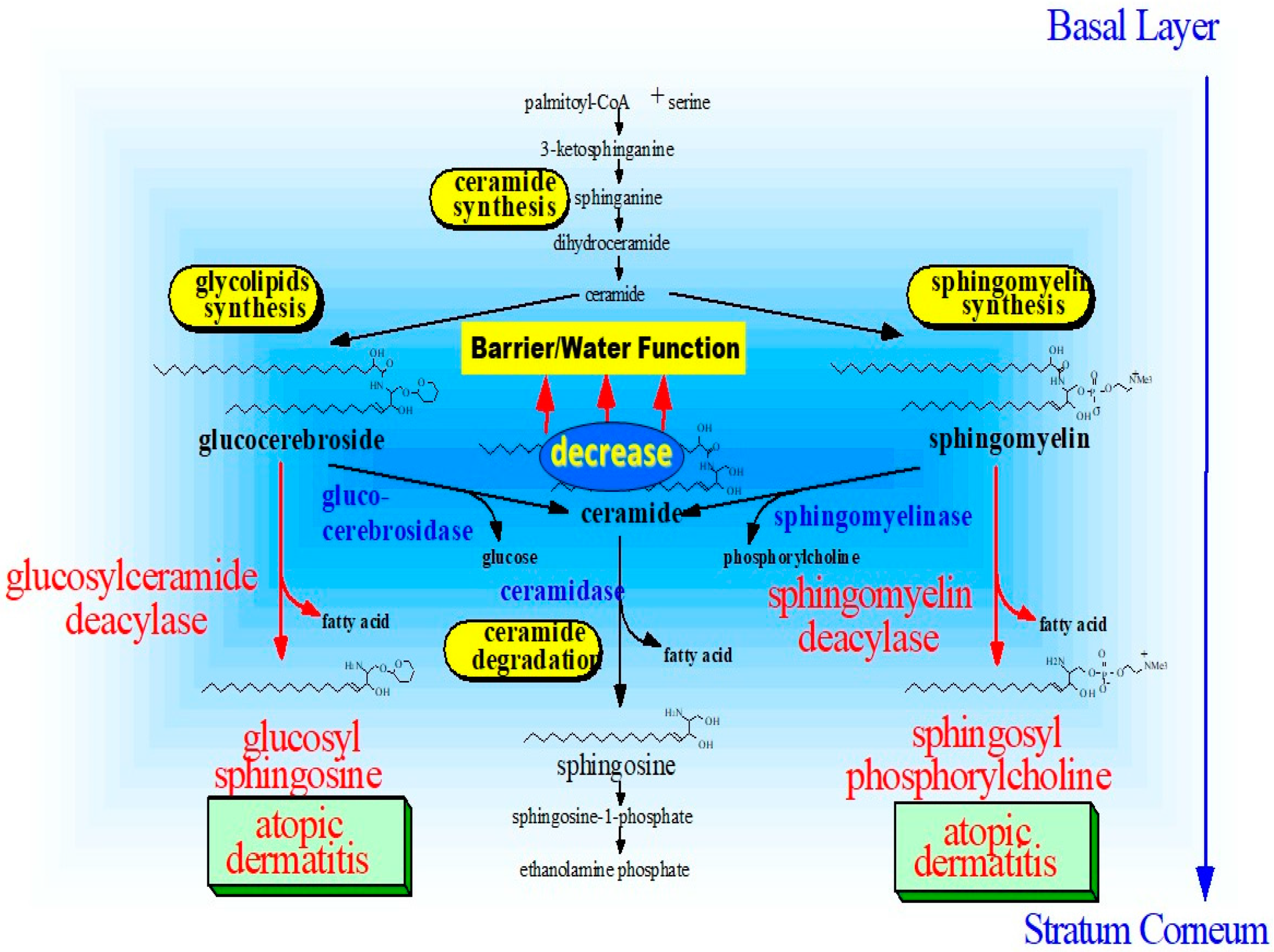
9. Discovery of Sphingomyelin Deacylase
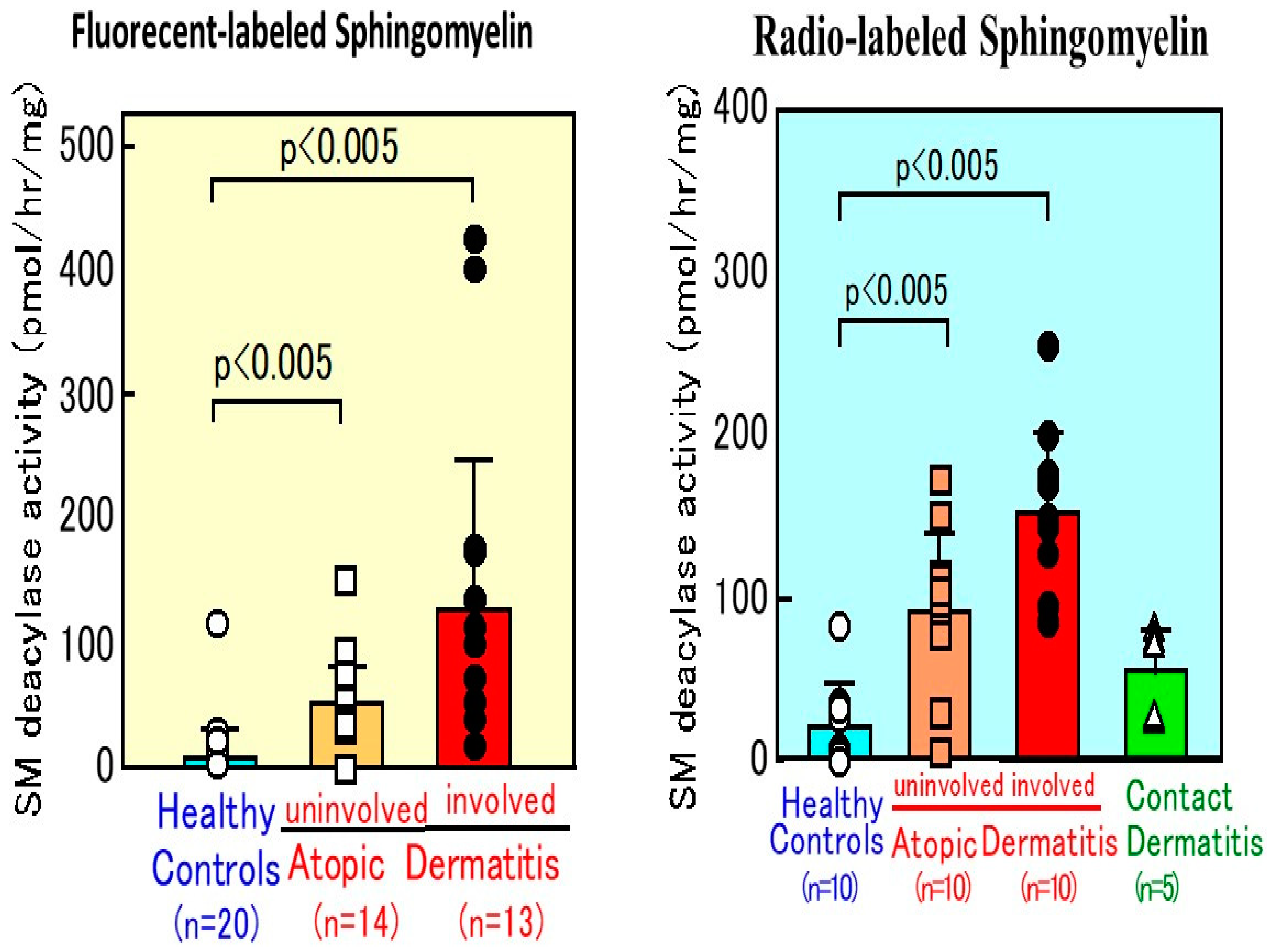
10. SM Deacylase Activity in AD Skin
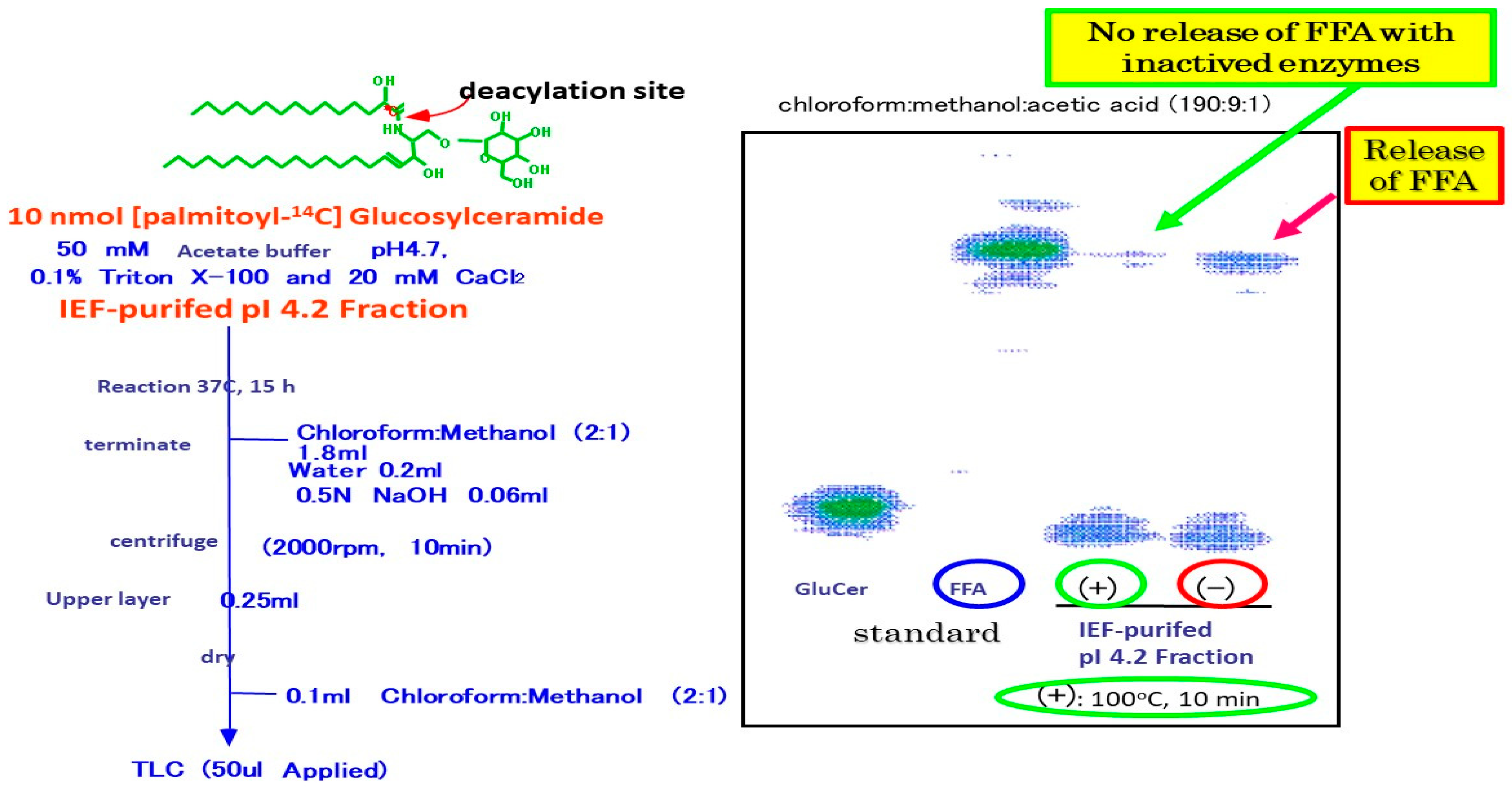
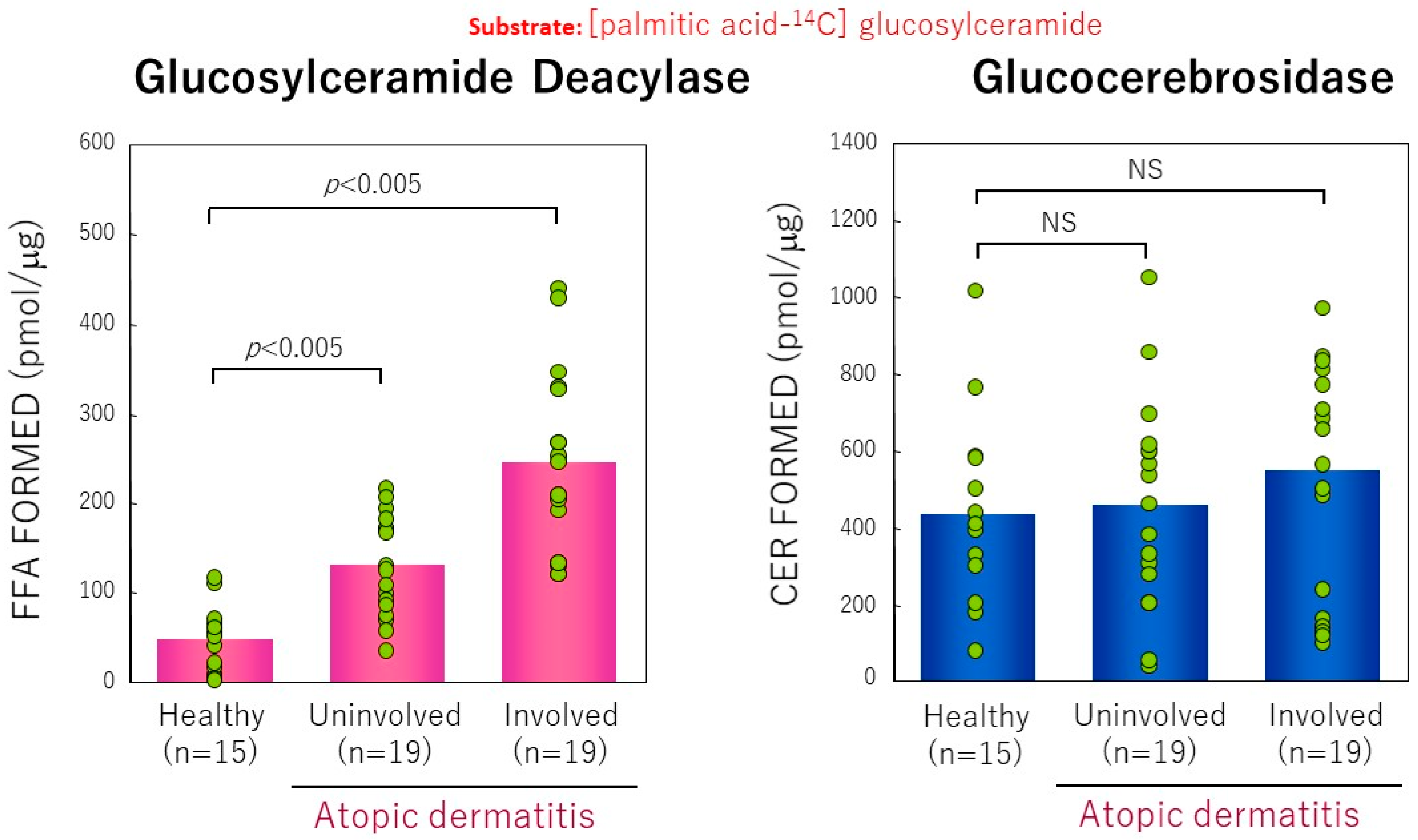
11. GCer Deacylase Activity in AD Skin
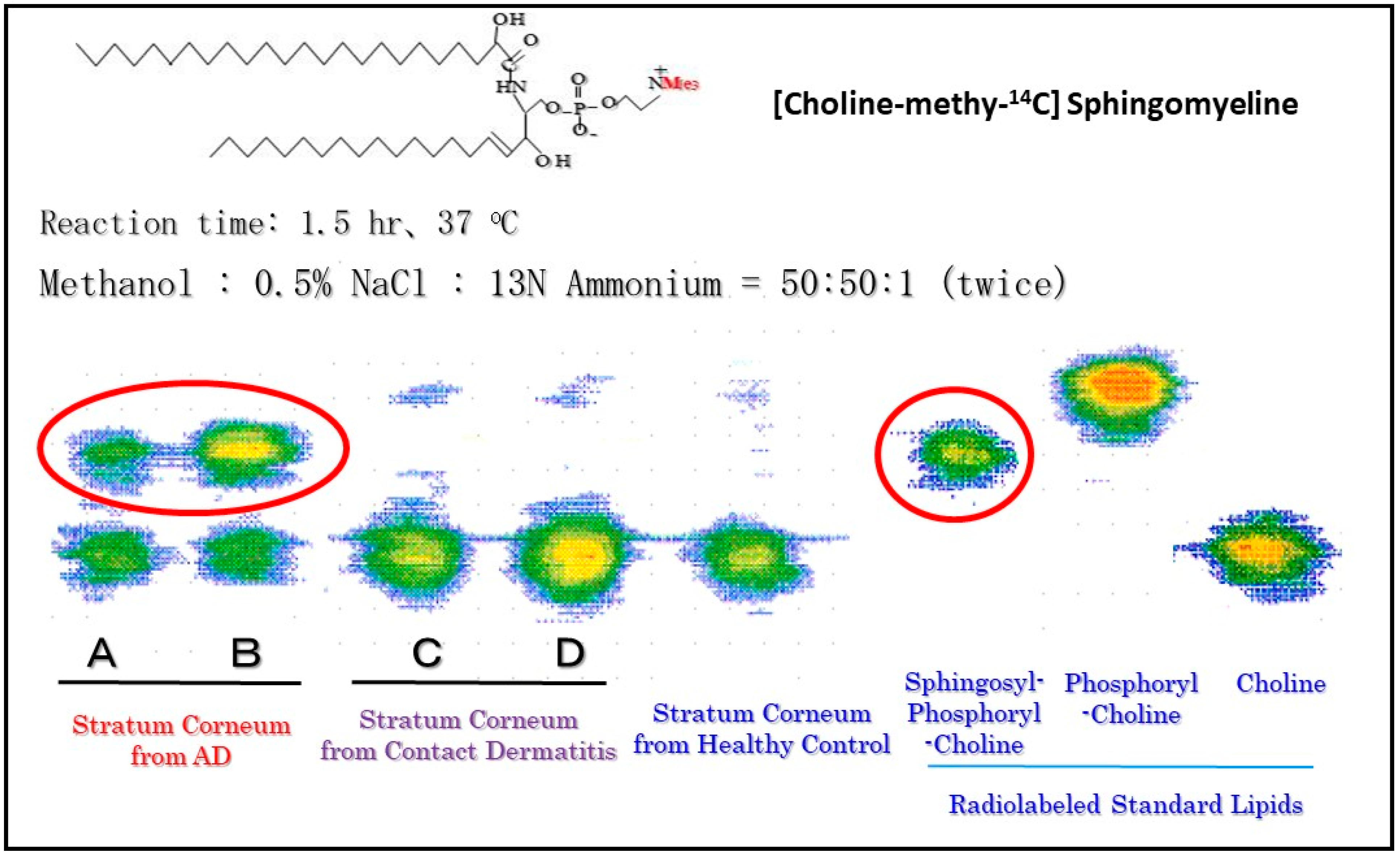
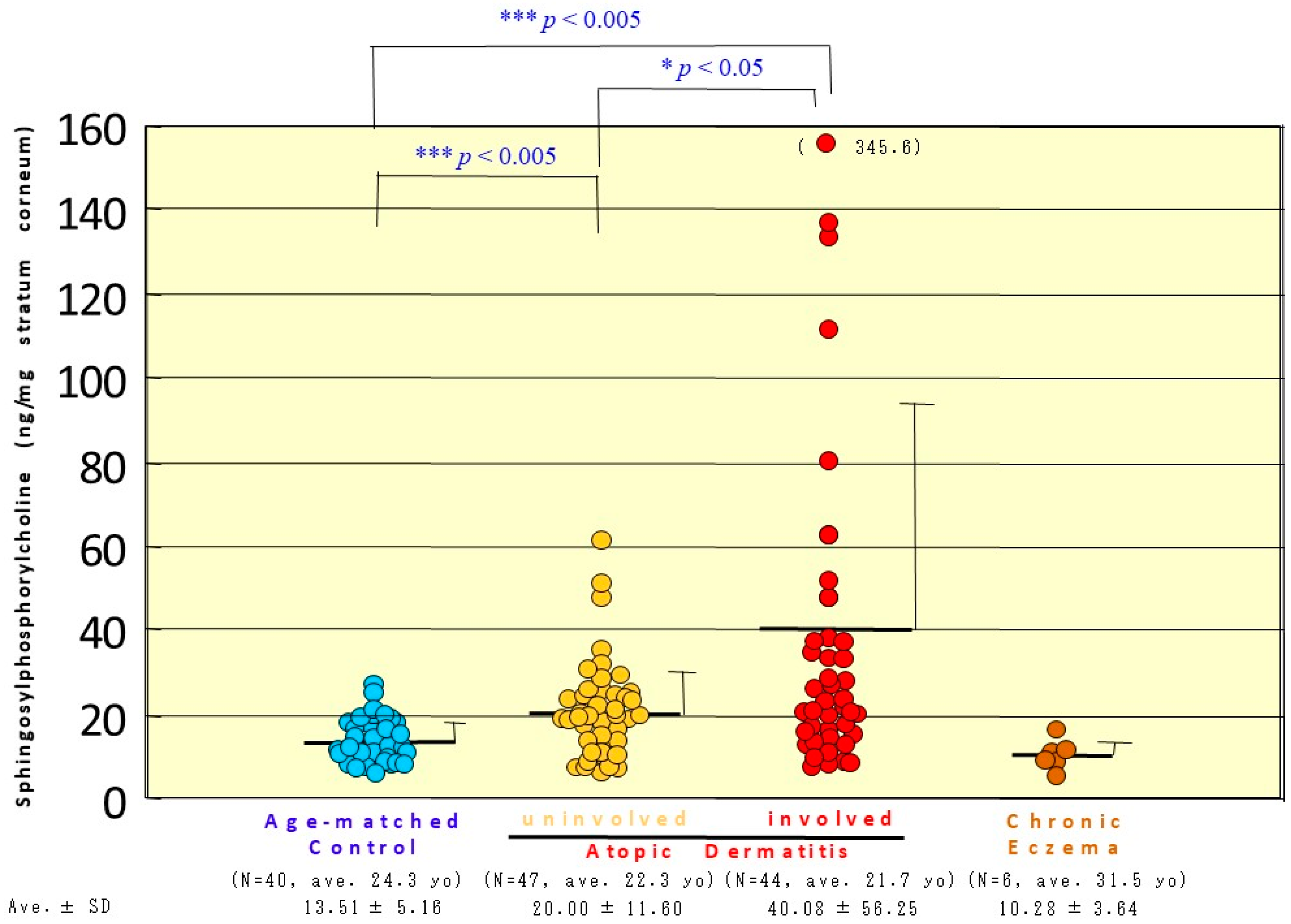
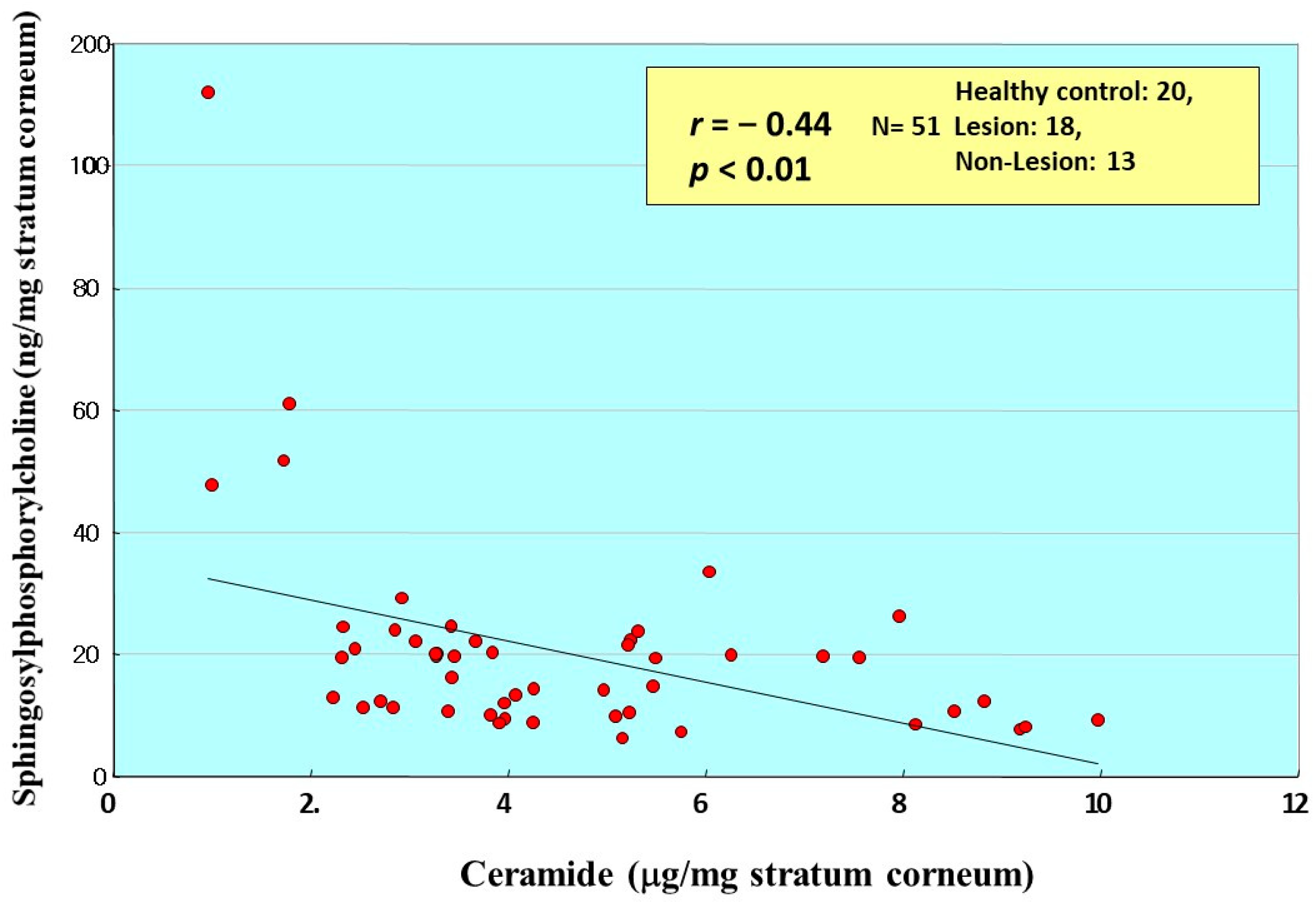
12. Accumulation of SPC as Evidence for Functional SM Deacylase
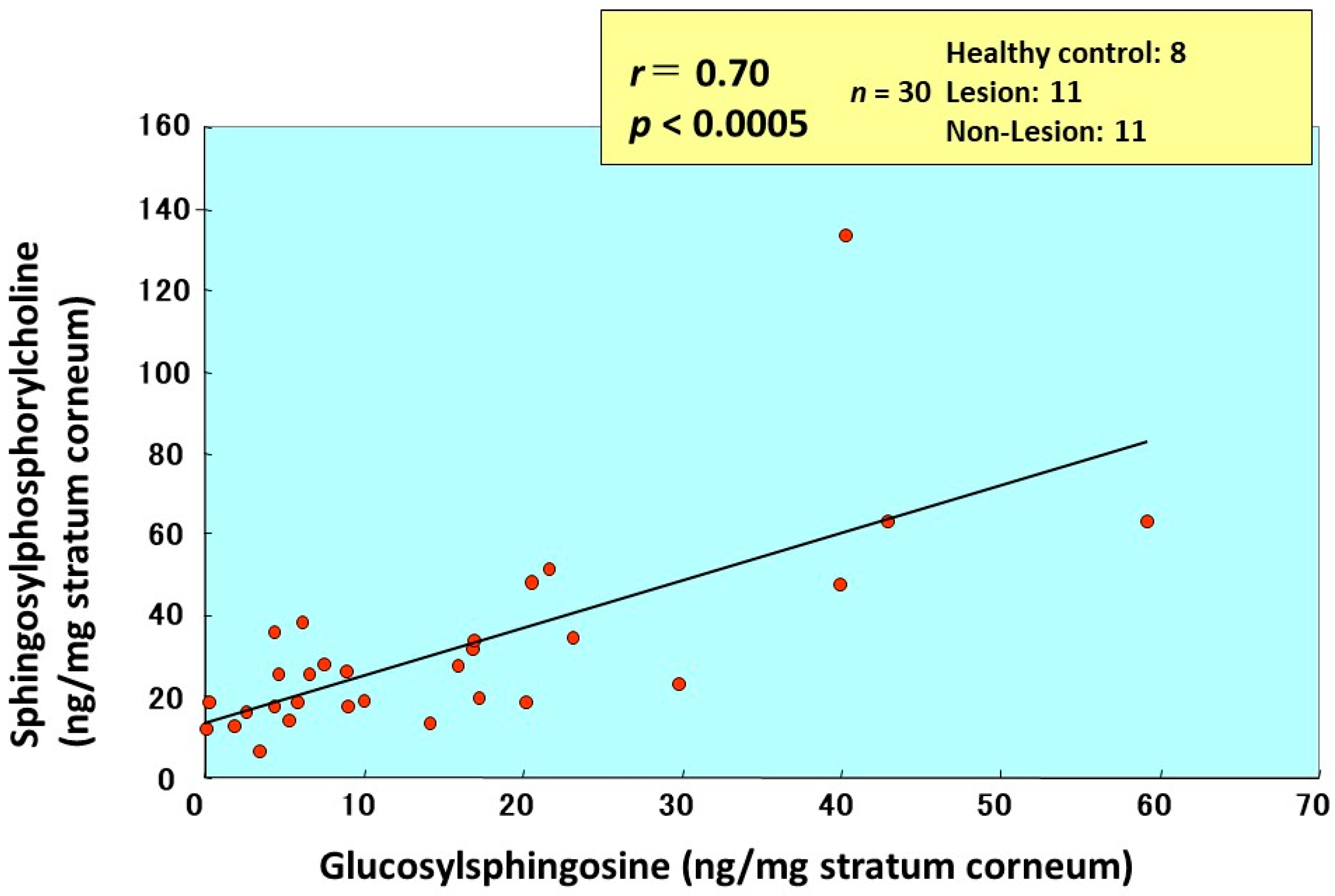
13. Accumulation of GSP as Evidence for Functional GCer Deacylase
14. Altered Sphingolipid Metabolism May Contribute to Atopic Skin Phenotypes (Inflammation, Roughened Skin and Hyperpigmentation)
15. Identification of SM Deacylase at the Gene and Protein Levels
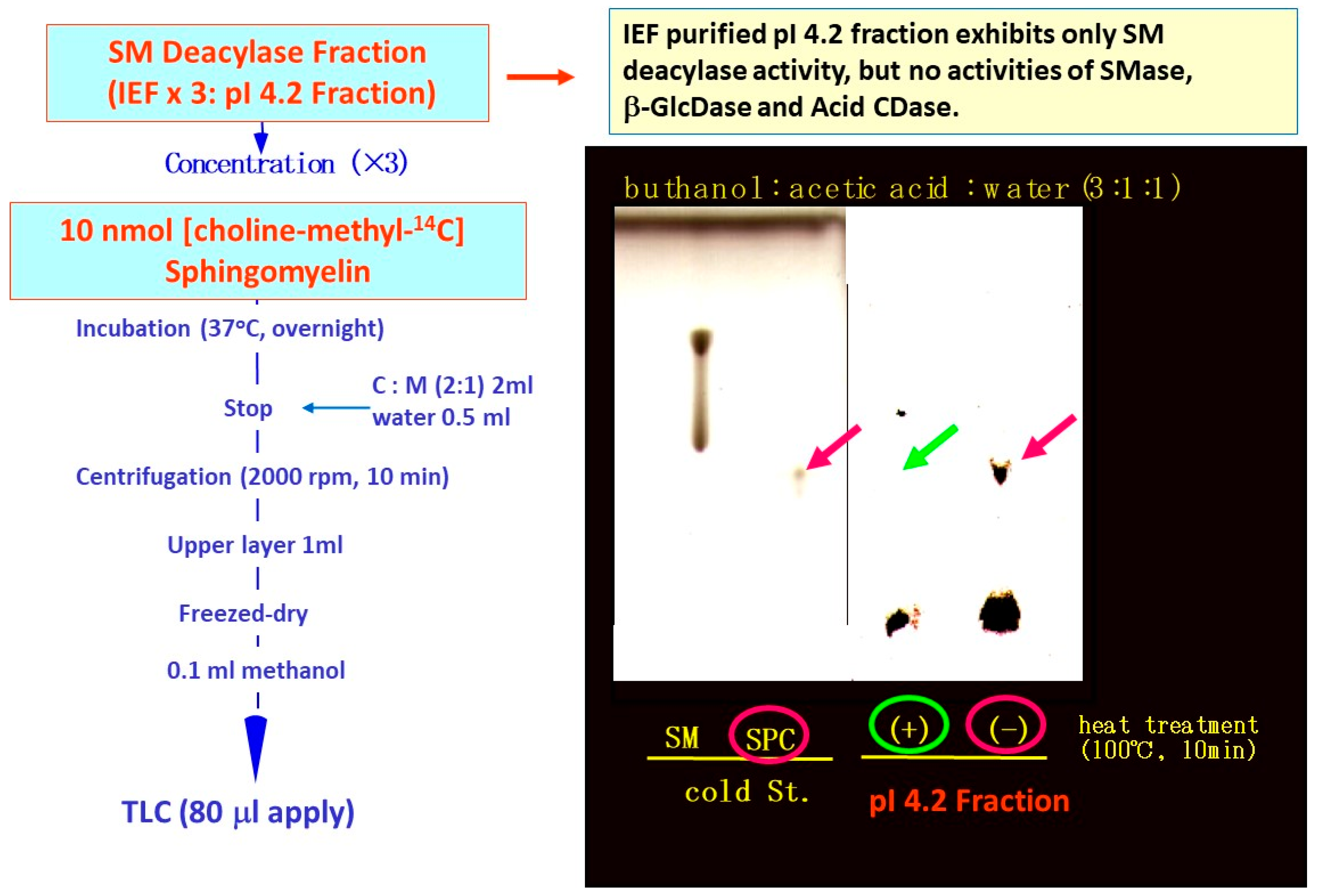
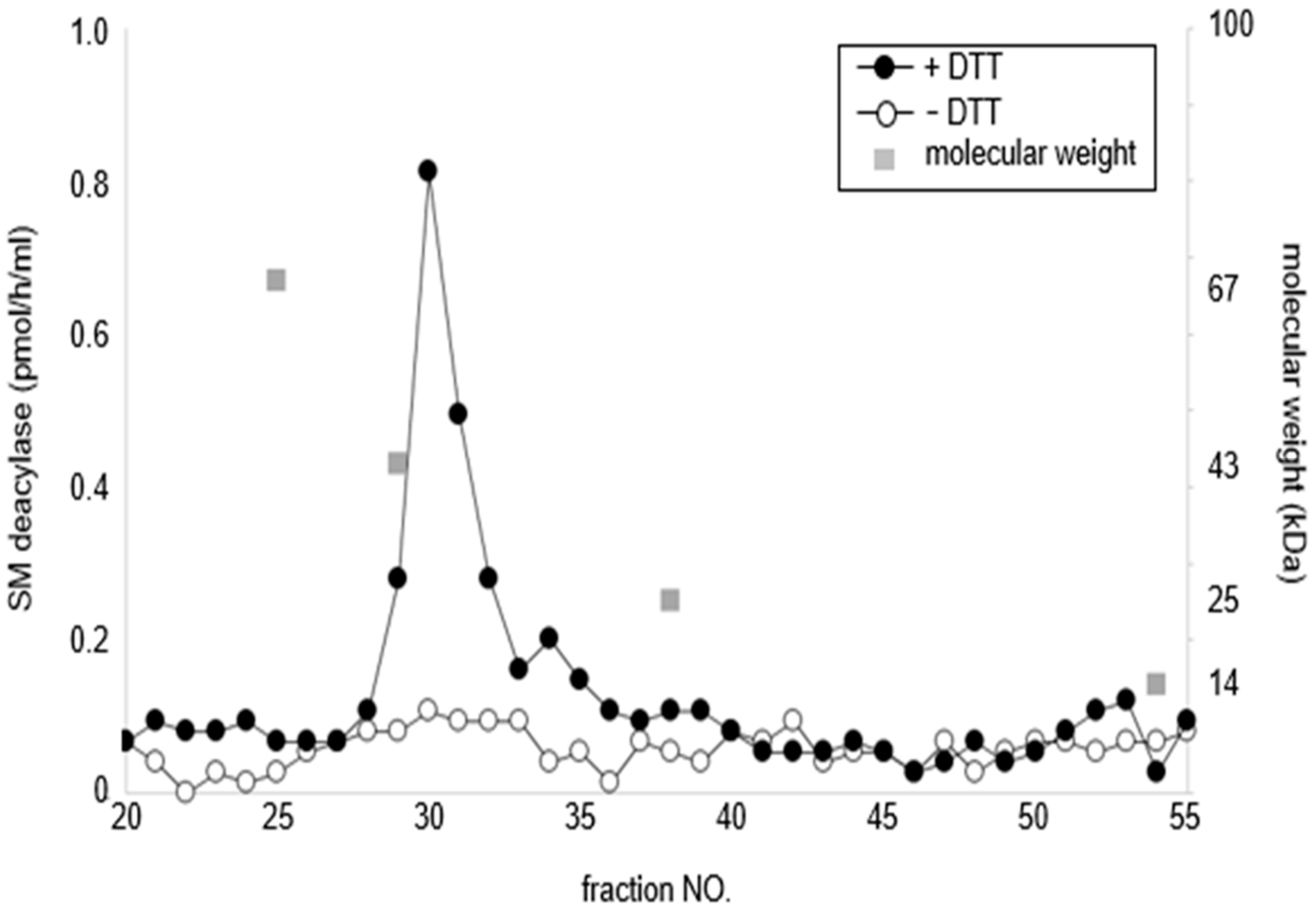
16. Purification of SM Deacylase
17. Enzymatic Properties of Purified SM Deacylase
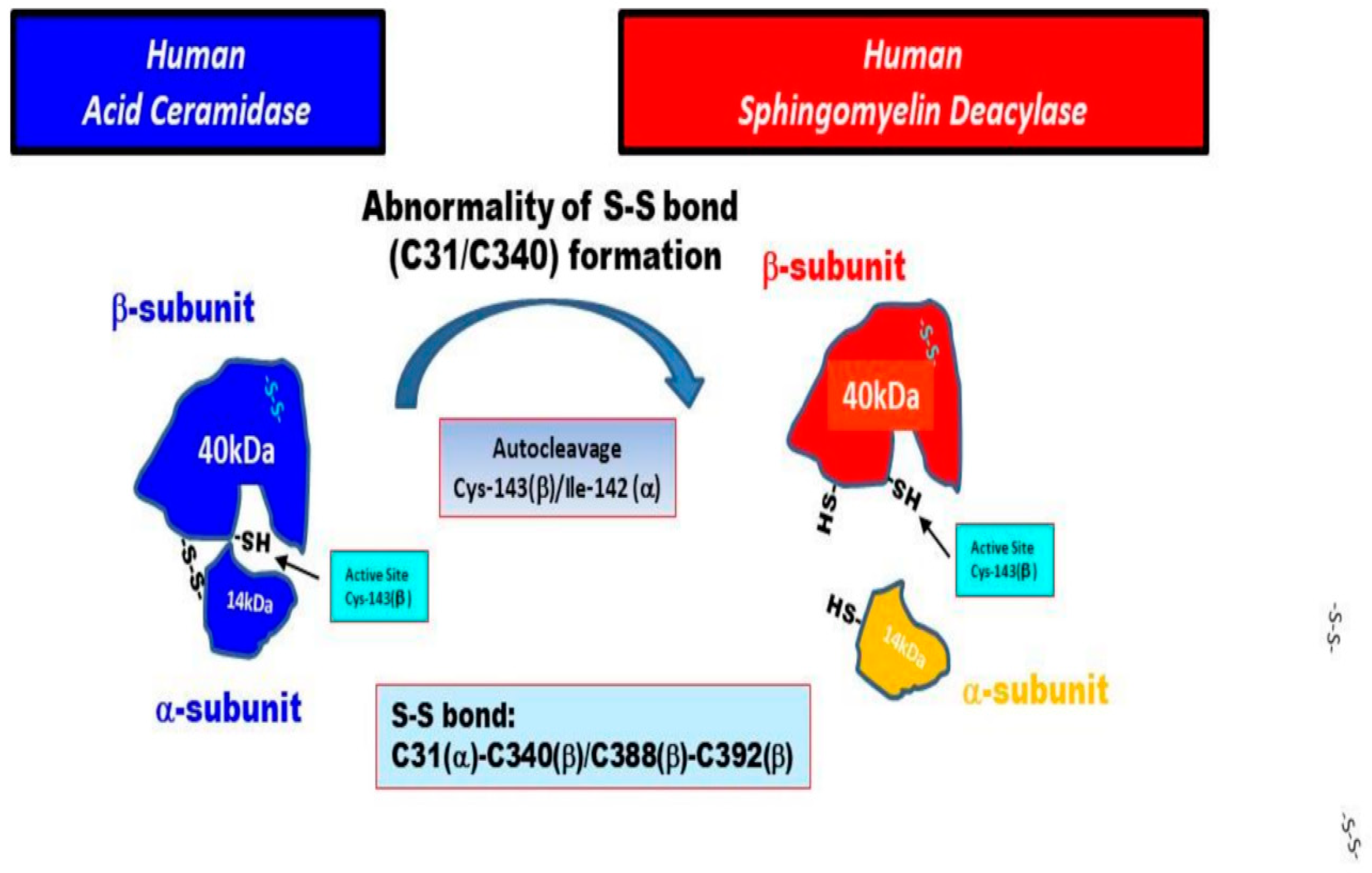
18. Identification of SM Deacylase at the Protein Level
19. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| acid ceramidase | aCDase |
| acid sphingomyelinase | aSMase |
| amiddleic dermatitis | AD |
| ceramide synthases | CERS |
| cell envelope | CE |
| conduritol b-epoxide | CBE |
| dithiothreitol | DTT |
| endothelin | EDN |
| endothelin B receptor | EDBR |
| GCer synthase | GCERS |
| globoid cell leukodystrophy | GLD |
| b-glucocerebrosidase | BGCase NOT GlcCDase |
| glucosylceramide | GCer |
| glucosylsphingosine | GSP |
| glycosphingolipids | GSL |
| healthy control | HC |
| intercellular adhesion molecule-1 | ICAM-1 |
| interleukin | IL |
| isoelectric focusing | IEF |
| lamellar granules | LGs |
| 2-mercaptoethanol | ME |
| microphthalmia-associated transcription factor | MITF |
| mitogen-activated protein | MAP |
| 1-oleoyl-2-acetyl-glycerol | OAG |
| phosphorylcholine | PC |
| photoacoustic spectrometry | PAS |
| pseudo-ceramide | pCer |
| radio-thin layer chromatography | RTLC |
| serine-palmitoyl transferase | SPT |
| SM synthase | SMS |
| sphingomyelin | SM |
| sphingosine | SS |
| sphingosine-1-phosphate | SSP |
| sphingosylphosphorylcholine | SPC |
| stearoyl CoA desaturase | SCD |
| stem cell factor | SCF |
| stratum corneum | SC |
| trans-epidermal water loss | TEWL |
| transglutaminases | TGase |
| tumor necrosis factor-α | TNFα |
References
- Nettis, E.; Ortoncelli, M.; Pellacani, G.; Foti, C.; Leo, E.D.; Patruno, C.; Rongioletti, F.; Argenziano, G.; Ferrucci, S.M.; Macchia, L.; et al. A multicenter study on the prevalence of clinical patterns and clinical phenotypes in adult atopic dermatitis. J. Investig. Allergol. Clin. Immunol. 2020, 30, 448–450. [Google Scholar] [CrossRef]
- Imokawa, G.; Ishida, K. Role of ceramide in the barrier function of the stratum corneum, Implications for the pathogenesis of atopic dermatitis. J. Clin. Exp. Derm. Res. 2014, 5, 206–218. [Google Scholar] [CrossRef] [Green Version]
- Hata, M.; Tokura, Y.; Takigawa, M.; Sato, M.; Shioya, Y.; Fujikura, Y.; Imokawa, G. Assessment of epidermal barrier function by photoacoustic spectrometry in relation to its importance in the pathogenesis of atopic dermatitis. Lab. Investig. 2002, 82, 1451–1461. [Google Scholar] [CrossRef] [Green Version]
- Matsuki, H.; Kiyokane, K.; Matsuki, T.; Sato, S.; Imokawa, G. Re-characterization of the non-lesional dry skin in atopic dermatitits through disrupted barrier function. Exog. Derm. 2004, 3, 282–292. [Google Scholar] [CrossRef]
- Hata, M.; Tokura, Y.; Takigawa, M.; Tamura, Y.; Imokawa, G. Efficacy of using pseudoCeramide-containg cream for the treatment of atopic dry skin in comparison with urea cream. Nishihihon J. Derm. 2002, 64, 606–611. [Google Scholar] [CrossRef]
- Takashima, I.; Hasebe, K.; Okuda, M.; Imokawa, G. Efficacy of pseudo-ceramide-containing detergent formulation for treatment of atopic dermatitis and asteatotic eczema. Nishihihon J. Derm. 2002, 64, 612–620. [Google Scholar] [CrossRef]
- Nakamura, T.; Satoh, H.; Imokawa, G.; Miyachi, Y. Clinical test of skin care products on atopic dry skin. Ski. Res. 2000, 42, 264–269. [Google Scholar]
- Yamanaka, M.; Tshibashi, O.; Takahashi, A.; Sato, H.; Imokawa, G. Clinical evaluation of Curel medicated cream in patients with atopic dermatitis. Ski. Res. 2001, 43, 278–285. [Google Scholar]
- Funasaka, Y.; Bito, T.; Yamamoto, M.; Nishigori, C.; Ishihashi, M.; Nakamura, T.; Ishida, K.; Sato, H.; Imokawa, G. Clinical evaluation of “Curel UV Milk” and “Curel UV Cream” in subjects with low barrier function’s skin. Ski. Res. 2004, 31, 62–74. [Google Scholar]
- Mizutani, J.; Takahashi, M.; Shimizu, M.; Kariya, N.; Sato, H.; Imokawa, G. Usage of pseudo-ceramide cream in atopic dry skin in comparison with 10% urea cream. Nishihihon J. Derm. 2001, 63, 457–461. [Google Scholar] [CrossRef]
- Akimoto, K.; Yoshikawa, N.; Higaki, Y.; Kawashima, M.; Imokawa, G. Quantitative analysis of stratum corneum lipids in xerosis and asteatotic eczema. J. Derm. 1993, 20, 1–6. [Google Scholar] [CrossRef]
- Ishida, K.; Takahashi, A.; Bito, K.; Draelos, Z.; Imokawa, G. Treatment with synthetic pudoceramide improves atopic skin, switching the ceramide profile to a healthy skin phenotype. J. Investig. Derm. 2020, 140, 1762–1770. [Google Scholar] [CrossRef]
- Halling-Overgaard, A.S.; Kezic, S.; Jakasa, I.; Engebretsen, K.A.; Maibach, H.; Thyssen, J.P. Skin absorption through atopic dermatitis skin: A systematic review. Br. J. Derm. 2017, 177, 84–106. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased level of ceramides in stratum corneum of atopic dermatitis: An etiologic factor in atopic dry skin? J. Investig. Derm. 1991, 96, 523–526. [Google Scholar] [CrossRef] [Green Version]
- Arikawa, J.; Ishibashi, M.; Kawashima, M.; Takagi, Y.; Ichikawa, Y.; Imokawa, G. Decreased levels of sphingosine, a natural antimicrobial agent, may be associated with vulnerability of the stratum corneum from patients with atopic dermatitis to colonization by Staphylococcus aureus. J. Investig. Derm. 2002, 119, 433–439. [Google Scholar] [CrossRef]
- Di Nardo, A.; Wertz, P.; Giannetti, A.; Seidenari, S. Ceramide and cholesterol composition of the skin of patients with atopic dermatitis. Acta Derm. Venereol. 1998, 78, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, M.; Arikawa, J.; Okamoto, R.; Kawashima, M.; Takagi, Y.; Ohguchi, K.; Imokawa, G. Abnormal expression of the novel epidermal enzyme, glucosylceramide deacylase, and the accumulation of its enzymatic reaction product, glucosylsphingosine, in the skin of patients with atopic dermatitis. Lab. Investig. 2003, 83, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, J.; Narita, H.; Kondo, N.; Hotta, M.; Takagi, Y.; Masukawa, Y.; Kitahara, T.; Takema, Y.; Koyano, S.; Yamazaki, S.; et al. Changes in the ceramide profile of atopic dermatitis patients. J. Investig. Derm. 2010, 130, 2511–2514. [Google Scholar] [CrossRef] [Green Version]
- Jungersted, J.M.; Scheer, H.; Mempel, M.; Baurecht, H.; Cifuentes, L.; Høgh, J.K.; Hellgren, L.I.; Jemec, G.B.E.; Agner, T.; Weidinger, S. Stratum corneum lipids, skin barrier function and filaggrin mutations in patients with atopic eczema. Allergy 2010, 65, 911–918. [Google Scholar] [CrossRef]
- Okamoto, R.; Arikawa, J.; Ishibashi, M.; Kawashima, M.; Takagi, Y.; Imokawa, G. Sphingosylphosphorylcholine is upregulated in the stratum corneum of patients with atopic dermatitis. J. Lipid Res. 2003, 44, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, A.; Nomura, T.; Mizuno, A.; Imokawa, G. Reevaluation of the non-lesional dry skin in atopic dermatitis by acute barrier disruption: An abnormal permeability barrier homeostasis with defective processing to generate ceramide. Arch. Derm. Res. 2014, 306, 427–440. [Google Scholar] [CrossRef]
- Choi, M.J.; Maibach, H.I. Role of ceramides in barrier function of healthy and diseased skin. Am. J. Clin. Derm. 2005, 6, 215–223. [Google Scholar] [CrossRef]
- Kim, D.; Lee, N.R.; Park, S.Y.; Jun, M.; Lee, K.; Kim, S.; Park, C.S.; Liu, K.H.; Choi, E.H. As in atopic dermatitis, nonlesional skin in allergic contact dermatitis displays abnormalities in barrier function and ceramide content. J. Investig. Derm. 2017, 137, 748–750. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G.; Hattori, M. A possible function of structural lipid in the water-holding properties of the stratum corneum. J. Investig. Derm. 1985, 84, 282–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imokawa, G.; Kuno, H.; Kawai, M. Stratum corneum lipids serve as a bound-water modulator. J. Investig. Derm. 1991, 96, 845–851. [Google Scholar] [CrossRef]
- Holleran, W.M.; Feingold, K.R.; Man, M.Q.; Gao, W.N.; Lee, J.M.; Elias, P.M. Regulation of epidermal sphingolipid synthesis by permeability barrier function. J. Lipid Res. 1991, 32, 1151–1158. [Google Scholar] [CrossRef]
- Holleran, W.M.; Man, M.Q.; Gao, W.N.; Menon, G.K.; Elias, P.M.; Feingold, K.R. Sphingolipids are required for mammalian epidermal barrier function. Inhibition of sphingolipid synthesis delays barrier recovery after acute perturbation. J. Clin. Investig. 1991, 88, 1338–1345. [Google Scholar] [CrossRef] [Green Version]
- Holleran, W.M.; Takagi, Y.; Menon, G.K.; Legler, G.; Feingold, K.R.; Elias, P.M. Processing of epidermal glucosylceramides is required for optimal mammalian cutaneous permeability barrier function. J. Clin. Investig. 1993, 91, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Holleran, W.M.; Ginns, E.I.; Menon, G.K.; Grundmann, J.U.; Fartasch, M.; McKinney, C.E.; Elias, P.M.; Sidransky, E. Consequences of beta-glucocerebrosidase deficiency in epidermis. Ultrastructure and permeability barrier alterations in Gaucher disease. J. Clin. Investig. 1994, 93, 1756–1764. [Google Scholar] [CrossRef] [Green Version]
- Holleran, W.M.; Takagi, Y.; Menon, G.K.; Jackson, S.M.; Lee, J.M.; Feingold, K.R.; Elias, P.M. Permeability barrier requirements regulate epidermal beta-glucocerebrosidase. J. Lipid Res. 1994, 35, 905–912. [Google Scholar] [CrossRef]
- Janssens, M.; van Smeden, J.; Gooris, G.S.; Bras, W.; Portale, G.; Caspers, P.J.; Vreeken, R.J.; Kezic, S.; Lavrijsen, A.P.; Bouwstra, J.A. Lamellar lipid organization and ceramide composition in the stratum corneum of patients with atopic eczema. J. Investig. Derm. 2011, 131, 2136–2138. [Google Scholar] [CrossRef] [Green Version]
- Matsuki, H.; Kiyokane, K.; Matsuki, T.; Sato, S.; Imokawa, G. Re-evaluation of the importance of barrier dysfunction in the non-lesional dry skin of atopic dermatitis through the use of two barrier creams. Exog. Derm. 2004, 3, 293–302. [Google Scholar] [CrossRef]
- Imokawa, G.; Akasaki, S.; Hattori, M.; Yoshizuka, N. Selective recovery of deranged water-holding properties by stratum corneum lipids. J. Investig. Derm. 1986, 87, 758–761. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G.; Akasaki, S.; Minematsu, Y.; Kawai, M. Importance of intercellular lipids in water-retention properties of the stratum corneum: Induction and recovery study of surfactant dry skin. Arch. Derm. Res. 1989, 281, 45–51. [Google Scholar] [CrossRef]
- Imokawa, G. Surfactant-induced depletion of ceramides and other intercellular lipids: Implication for the mechanism leading to dehydration of the stratum corneum. Exog. Derm. 2004, 3, 81–98. [Google Scholar] [CrossRef]
- Imokawa, G.; Akasaki, S.; Minematsu, Y.; Kuno, O.; Zama, M.; Kawai, M.; Hattori, M.; Yoshizuka, N.; Kawamata, A.; Yano, Y.; et al. Function of lipids on human skin. J. Disp. Sci. Tech. 1989, 10, 617–641. [Google Scholar] [CrossRef]
- Imokawa, G.; Akasaki, S.; Kawamata, A.; Yano, S.; Takaishi, N. Water-retaining function in the stratum corneum and its recovery properties by synthetic pseudo-ceramides. J. Soc. Cosmet. Chem. 1989, 40, 273–285. [Google Scholar]
- Imokawa, G. Water and The Stratum Corneum, In vitro and in vivo models. In Bioengineering of Skin; Elsner, P., Berardesca, E., Maibach, H.I., Eds.; CRC Press: Boca Raton, FL, USA, 1994; Volume 1, Chapter 3; pp. 23–47. [Google Scholar]
- Shen, C.P.; Zhao, M.T.; Jia, Z.X.; Zhang, J.L.; Jiao, L.; Ma, L. Skin ceramide profile in children with atopic dermatitis. Dermatitis 2018, 29, 219–222. [Google Scholar] [CrossRef]
- Van Smeden, J.; Hoppel, L.; van der Heijden, R.; Hankemeier, T.; Vreeken, R.J.; Bouwstra, J.A. LC/MS analysis of stratum corneum lipids: Ceramide profiling and discovery. J. Lipid Res. 2011, 52, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Koyano, S.H.A.; Yamazaki, S.; Ishikawa, J.; Kitahara, T.; Narita, H.; Kondo, N.; Masukawa, Y. Psoriasis patients have abnormal ceramide profile in stratum corneum. Nishihihon J. Derm. 2010, 72, 494–499. [Google Scholar] [CrossRef]
- Kikuchi, K.; Kobayashi, H.; O’goshi, K.; Tagami, H. Impairment of skin barrier function is not inherent in atopic dermatitis patients: A prospective study conducted in newborns. Pediatr. Derm. 2006, 23, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G. Role of stratum corneum ceramide in water and barrier functions: Implication to pathogenesis of atopic dermatitis. In Cutting Edge on Molecular Mechanisms of Skin Ageing and Its Rational Anti-Aging Treatments and Pathogenesis of Atopic Dermatitis; Imokawa, G., Ed.; Book Publisher International: London, UK, 2021; Chapter 4; pp. 146–148, 172–173. [Google Scholar] [CrossRef]
- Smith, F.J.D.; Irvine, A.D.; Terron-Kwiatkowski, A.; Sandilands, A.; Campbell, L.E.; Zhao, Y.; Liao, H.; Evans, A.T.; Goudie, D.R.; Lewis-Jones, S.; et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris. Nat. Genet. 2006, 38, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Sandilands, A.; O’Regan, G.M.; Liao, H.; Zhao, Y.; Terron-Kwiatkowski, A.; Watson, R.M.; Cassidy, A.J.; Goudie, D.R.; Smith, F.J.D.; McLean, W.H.I.; et al. Prevalent and rare mutations in the gene encoding filaggrin cause ichthyosis vulgaris and predispose individuals to atopic dermatitis. J. Investig. Dermatol. 2006, 126, 1770–1775. [Google Scholar] [CrossRef] [Green Version]
- Sandilands, A.; Smith, F.J.; Irvine, A.D.; McLean, W.H. Filaggrin’s fuller figure: A glimpse into the genetic architecture of atopic dermatitis. J. Investig. Dermatol. 2007, 127, 1282–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Schmuth, M.; Man, M.Q.; Weber, F.; Gao, W.; Feingold, K.R.; Fritsch, P.; Elias, P.M.; Holleran, W.M. Permeability barrier disorder in Niemann-Pick disease: Sphingomyelin-ceramide processing required for normal barrier homeostasis. J. Investig. Derm. 2000, 115, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.; Higaki, Y.; Takagi, Y.; Higuchi, K.; Yada, Y.; Kawashima, M.; Imokawa, G. Analysis of beta-glucocerebrosidase and ceramidase activities in atopic and aged dry skin. Acta Derm. Venereol. 1994, 74, 337–340. [Google Scholar]
- Hara, J.; Higuchi, K.; Okamoto, R.; Kawashima, M.; Imokawa, G. High-expression of sphingomyelin deacylase is an important determinant of ceramide deficiency leading to barrier disruption in atopic dermatitis. J. Investig. Derm. 2000, 115, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Kusuda, S.; Cui, C.Y.; Takahashi, M.; Tezuka, T. Localization of sphingomyelinase in lesional skin of atopic dermatitis patients. J. Investig. Derm. 1998, 111, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Danso, M.; Boiten, W.; van Drongelen, V.; Meijling, K.G.; Gooris, G.; Ghalbzouri, A.E.; Absalah, S.; Vreeken, R.; Kezic, S.; van Smeden, J.; et al. Altered expression of epidermal lipid bio-synthesis enzymes in atopic dermatitis skin is accompanied by changes in stratum corneum lipid composition. J. Derm. Sci. 2017, 88, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Jensen, J.M.; Fölster-Holst, R.; Baranowsky, A.; Schunck, M.; Winoto-Morbach, S.; Neumann, C.; Schütze, S.; Proksch, E. Impaired sphingomyelinase activity and epidermal differentiation in atopic dermatitis. J. Investig. Derm. 2004, 122, 1423–1431. [Google Scholar] [CrossRef] [Green Version]
- Murata, Y.; Ogata, J.; Higaki, Y.; Kawashima, M.; Yada, Y.; Higuchi, K.; Tsuchiya, T.; Kawainami, S.; Imokawa, G. Abnormal expression of sphingomyelin acylase in atopic dermatitis: An etiologic factor for ceramide deficiency? J. Investig. Derm. 1996, 106, 1242–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, K.; Hara, J.; Okamoto, R.; Kawashima, M.; Imokawa, G. The skin of atopic dermatitis patients contains a novel enzyme, glucosylceramide sphingomyelin deacylase, which cleaves the N-acyl linkage of sphingomyelin and glucosylceramide. Biochem. J. 2000, 350, 747–756. [Google Scholar] [CrossRef]
- Yada, Y.; Higuchi, K.; Imokawa, G. Purification and biochemical characterization of membrane-bound epidermal ceramidases from guinea pig skin. J. Biol. Chem. 1995, 270, 12677–12684. [Google Scholar] [CrossRef] [Green Version]
- Holleran, W.M.; Takagi, Y.; Imokawa, G.; Jackson, S.; Lee, J.M.; Elias, P.M. Beta-Glucocerebrosidase activity in murine epidermis: Characterization and localization in relation to differentiation. J. Lipid Res. 1992, 33, 1201–1209. [Google Scholar] [CrossRef]
- Takagi, Y.; Kriehuber, E.; Imokawa, G.; Elias, P.M.; Holleran, W.M. Beta-Glucocerebrosidase activity in mammalian stratum corneum. J. Lipid Res. 1999, 40, 861–869. [Google Scholar] [CrossRef]
- Bouwstra, J.A.; Dubbelaar, F.E.; Gooris, G.S.; Weerheim, A.M.; Ponec, M. The role of ceramide composition in the lipid organisation of the skin barrier. Biochim. Biophys. Acta 1999, 1419, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G.; Yada, Y.; Higuchi, K.; Okuda, M.; Ohashi, Y.; Kawamata, A. Pseudo-acylceramide with linoleic acid produces selective recovery of diminished cutaneous barrier function in essential fatty acid deficient rats and has an inhibitory effect on epidermal hyperplasia. J. Clin. Investig. 1994, 94, 89–96. [Google Scholar] [CrossRef]
- Umeda, Y.; Mizutani, H.; Imokawa, G.; Shimizu, M. Topical ceramide corrected epidermal cell hyperproliferation and stratum corneum dysmaturation in atopic eczema. New Trends Allergy 1997, 4, 237–239. [Google Scholar]
- Takagi, Y.; Nakagawa, H.; Matsuo, N.; Nomura, T.; Takizawa, M.; Imokawa, G. Biosynthesis of acylceramide in murine epidermis: Characterization by inhibition of glucosylation and deglucosylation, and by substrate specificity. J. Investig. Derm. 2004, 122, 722–729. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Lafrasse, C.; Vanier, M.T. Sphingosylphosphorylcholine in Niemann-Pick disease brain: Accumulation in type A but not in type B. Neurochem. Res. 1999, 24, 199–205. [Google Scholar] [CrossRef]
- Atsumi, S.; Nosaka, C.; Iinuma, H.; Umezawa, K. Accumulation of tissue glucosylsphingosine in Gaucher-like mouse induced by the glucosylceramidase inhibitor cyclophellitol. Arch. Biochem. Biophys. 1993, 304, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Sasagasako, N.; Goto, I.; Kobayashi, T. The synthetic pathway for glucosylsphingosine in cultured fibroblasts. J. Biochem. 1994, 116, 704–710. [Google Scholar] [CrossRef]
- Murugesan, V.; Chuang, W.L.; Liu, J.; Lischuk, A.; Kacena, K.; Lin, H.; Pastores, G.M.; Yang, R.; Keutzer, J.; Zhang, K.; et al. Glucosylsphingosine is a key biomarker of Gaucher disease. Am. J. Hematol. 2016, 91, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, T.; Nakamura, S.; Momoi, M.; Yamaji, T.; Takematsu, H.; Yano, H.; Sabe, H.; Yamamoto, A.; Kawasaki, T.; Kozutsumi, Y. Inhibition of cytokinesis by a lipid metabolite, psychosine. J. Cell Biol. 2000, 149, 943–950. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G.; Takagi, Y.; Higuchi, K.; Kondo, H.; Yada, Y. Sphingosylphosphorylcholine is a potent inducer of intercellular adhesion molecule-1 expression in human keratinocytes. J. Investig. Derm. 1999, 112, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, K.; Kawashima, M.; Takagi, Y.; Kondo, H.; Yada, Y.; Ichikawa, Y.; Imokawa, G. Sphingosylphosphorylcholine is an activator of transglutaminase activity in human keratinocytes. J. Lipid Res. 2001, 42, 1562–1570. [Google Scholar] [CrossRef]
- Higuchi, K.; Kawashima, M.; Ichikawa, Y.; Imokawa, G. Sphingosylphosphorylcholine is a melanogenic stimulator for human melanocytes. Pigment. Cell Res. 2003, 16, 670–678. [Google Scholar] [CrossRef]
- Teranishi, Y.; Kuwahara, H.; Ueda, M.; Takemura, T.; Kusumoto, M.; Nakamura, K.; Sakai, J.; Kimura, T.; Furutani, Y.; Kawashima, M.; et al. Sphingomyelin deacylase, the enzyme involved in the pathogenesis of atopic dermatitis, is identical to the β-subunit of acid ceramidase. Int. J. Mol. Sci. 2020, 21, 8789. [Google Scholar] [CrossRef]
- Oizumi, A.; Nakayama, H.; Okino, N.; Iwahara, C.; Kina, K.; Matsumoto, R.; Ogawa, H.; Takamori, K.; Ito, M.; Suga, Y.; et al. Pseudomonas-derived ceramidase induces production of inflammatory mediators from human keratinocytes via sphingosine-1-phosphate. PLoS ONE 2014, 9, e89402. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, K.; Hurwitz, R.; Zenk, T.; Desnick, R.J.; Ferlinz, K.; Schuchman, E.H.; Sandhoff, K. Purification, characterization, and biosynthesis of human acid ceramidase. J. Biol. Chem. 1995, 270, 11098–11102. [Google Scholar] [CrossRef] [Green Version]
- Linke, T.; Wilkening, G.; Sadeghlar, F.; Mozcall, H.; Bernardo, K.; Schuchman, E.; Sandhoff, K. Interfacial regulation of acid ceramidase activity. Stimulation of ceramide degradation by lysosomal lipids and sphingolipid activator proteins. J. Biol. Chem. 2001, 276, 5760–5768. [Google Scholar] [CrossRef] [Green Version]
- Koch, J.; Gartner, S.; Li, C.M.; Quintern, L.E.; Bernardo, K.; Levran, O.; Schnabel, D.; Desnick, R.J.; Schuchman, E.H.; Sandhoff, K. Molecular cloning and characterization of a full-length complementary DNA encoding human acid ceramidase. Identification Of the first molecular lesion causing Farber disease. J. Biol. Chem. 1996, 271, 33110–33115. [Google Scholar] [CrossRef] [Green Version]
- Ferlinz, K.; Kopal, G.; Bernardo, K.; Linke, T.; Bar, J.; Breiden, B.; Neumann, U.; Lang, F.; Schuchman, E.H.; Sandhoff, K. Human acid ceramidase: Processing, glycosylation, and lysosomal targeting. J. Biol. Chem. 2001, 276, 35352–35360. [Google Scholar] [CrossRef] [Green Version]
- Gebai, A.; Gorelik, A.; Li, Z.; Illes, K.; Nagar, B. Structural basis for the activation of acid ceramidase. Nat. Commun. 2018, 9, 1621. [Google Scholar] [CrossRef]
- Schulze, H.; Schepers, U.; Sandhoff, K. Overexpression and mass spectrometry analysis of mature human acid ceramidase. Biol. Chem. 2007, 388, 1333–1343. [Google Scholar] [CrossRef]
- Shtraizent, N.; Eliyahu, E.; Park, J.H.; He, X.; Shalgi, R.; Schuchman, E.H. Autoproteolytic cleavage and activation of human acid ceramidase. J. Biol. Chem. 2008, 283, 11253–11259. [Google Scholar] [CrossRef] [Green Version]
- Ferraz, M.J.; Marques, A.R.A.; Appelman, M.D.; Verhoek, M.; Strijland, A.; Mirzaian, M.; Scheij, S.; Ouairy, C.M.; Lahav, D.; Wisse, P.; et al. Lysosomal glycosphingolipid catabolism by acid ceramidase: Formation of glycosphingoid bases during deficiency of glycosidases. FEBS Lett. 2016, 590, 716–725. [Google Scholar] [CrossRef] [Green Version]
- Dattola, A.; Bennardo, L.; Silvestri, M.; Nisticò, S.P. What’s new in the treatment of atopic dermatitis? Dermatol. Ther. 2019, 32, e12787. [Google Scholar] [CrossRef]

































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endogenous Ceramide Species & pCer | Skin Conductance (µS) | Trans-Epidermal Water Loss (g/m2/h) | ||
|---|---|---|---|---|
| Correlation Coefficient | p-Value | Correlation Coefficient | p-Value | |
| pCer | 0.436 | 0.0056 ** | 0.1015 | 0.5386 |
| Total Ceramides | −0.2372 | 0.1459 | -0.0769 | 0.6417 |
| Cer [NDS] | 0.2357 | 0.1478 | -0.1312 | 0.4259 |
| Cer [NS] | −0.2141 | 0.1905 | 0.1739 | 0.2898 |
| Cer [NH] | −0.2049 | 0.2108 | 0.0157 | 0.9242 |
| Cer [NP] | −0.1506 | 0.36 | −0.1586 | 0.335 |
| Cer [ADS] | −0.2069 | 0.2604 | −0.2302 | 0.1585 |
| Cer [AS] | −0.2399 | 0.1413 | 0.0867 | 0.5992 |
| Cer [AH] | −0.2054 | 0.2097 | 0.0017 | 0.9917 |
| Cer [AP] | −0.2609 | 0.1087 | −0.2694 | 0.0972 |
| Cer [EOS] | −0.1978 | 0.2275 | 0.1239 | 0.42 |
| Cer [EOH] | −0.2045 | 0.2117 | 0.0301 | 0.8559 |
| Cer [EOP] | −0.197 | 0.2292 | −0.1329 | 0.4119 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imokawa, G. Cutting Edge of the Pathogenesis of Atopic Dermatitis: Sphingomyelin Deacylase, the Enzyme Involved in Its Ceramide Deficiency, Plays a Pivotal Role. Int. J. Mol. Sci. 2021, 22, 1613. https://doi.org/10.3390/ijms22041613
Imokawa G. Cutting Edge of the Pathogenesis of Atopic Dermatitis: Sphingomyelin Deacylase, the Enzyme Involved in Its Ceramide Deficiency, Plays a Pivotal Role. International Journal of Molecular Sciences. 2021; 22(4):1613. https://doi.org/10.3390/ijms22041613
Chicago/Turabian StyleImokawa, Genji. 2021. "Cutting Edge of the Pathogenesis of Atopic Dermatitis: Sphingomyelin Deacylase, the Enzyme Involved in Its Ceramide Deficiency, Plays a Pivotal Role" International Journal of Molecular Sciences 22, no. 4: 1613. https://doi.org/10.3390/ijms22041613
APA StyleImokawa, G. (2021). Cutting Edge of the Pathogenesis of Atopic Dermatitis: Sphingomyelin Deacylase, the Enzyme Involved in Its Ceramide Deficiency, Plays a Pivotal Role. International Journal of Molecular Sciences, 22(4), 1613. https://doi.org/10.3390/ijms22041613