Invertebrate Gonadotropin-Releasing Hormone Receptor Signaling and Its Relevant Biological Actions



Abstract
:1. Introduction
2. Urochordate GnRH Signaling
2.1. Urochordate GnRHs
2.2. Urochordate GnRH Receptors
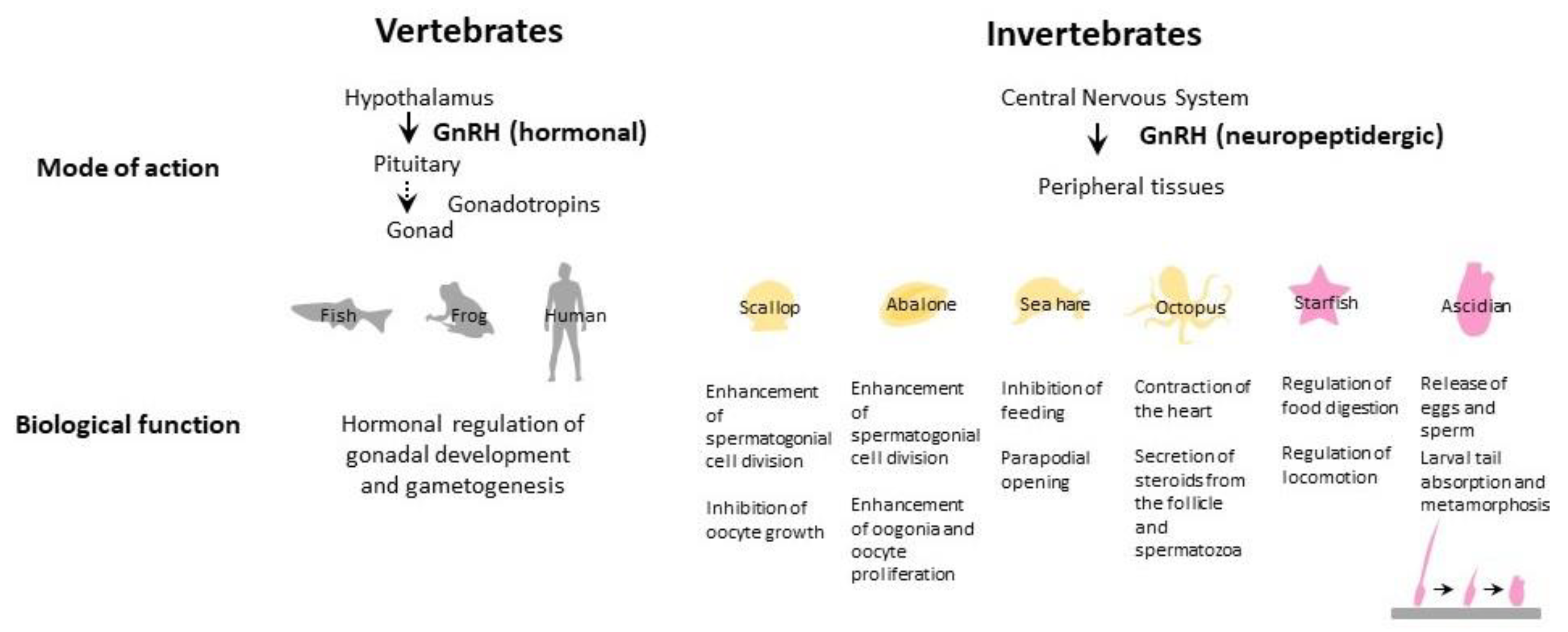
2.3. Biological Functions
2.3.1. Reproductive Effects
2.3.2. Nonreproductive Effects
3. Cephalochordate GnRH Signaling
3.1. Cephalochordate GnRHs
3.2. Cephalochordate Receptors
4. Echinoderm GnRH Signaling
4.1. Echinoderm GnRHs
4.2. Echinoderm GnRH Receptors
4.3. Biological Effects of Echinoderm GnRH
5. Protostome GnRH Signaling
5.1. Protostome GnRHs
5.2. Protostome GnRH Receptors
5.3. Biological Effects of Protostome GnRHs
6. Conclusions and Perspectives
Funding
Conflicts of Interest
References
- Holland, L.Z.; Sower, S.A. “Insights of early chordate genomics: Endocrinology and development in amphioxus, tunicates and lampreys”: Introduction to the symposium. Integr. Comp. Biol. 2010, 50, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Millar, R.P. GnRHs and GnRH receptors. Anim. Reprod. Sci. 2005, 88, 5–28. [Google Scholar]
- Abraham, E.; Palevitch, O.; Gothilf, Y.; Zohar, Y. Targeted gonadotropin-releasing hormone-3 neuron ablation in zebrafish: Effects on neurogenesis, neuronal migration, and reproduction. Endocrinology 2010, 151, 332–340. [Google Scholar]
- Nishiguchi, R.; Azuma, M.; Yokobori, E.; Uchiyama, M.; Matsuda, K. Gonadotropin-releasing hormone 2 suppresses food intake in the zebrafish, Danio rerio. Front. Endocrinol. 2012, 3, 122. [Google Scholar]
- Marvel, M.M.; Spicer, O.S.; Wong, T.T.; Zmora, N.; Zohar, Y. Knockout of Gnrh2 in zebrafish (Danio rerio) reveals its roles in regulating feeding behavior and oocyte quality. Gen. Comp. Endocrinol. 2019, 280, 15–23. [Google Scholar]
- Ogawa, S.; Parhar, I. Morphological evidence for functional crosstalk between multiple GnRH systems in the male tilapia, Oreochromis niloticus. Front. Endocrinol. 2020, 11, 586. [Google Scholar]
- Kawada, T.; Aoyama, M.; Sakai, T.; Satake, H. Structure, function, and evolutionary aspects of invertebrate GnRHs and their receptors. In Gonadotropin-Releasing Hormone (GnRH): Production, Structure and Functions; Scott-Sills, E., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2013; pp. 1–15. [Google Scholar]
- Naor, Z.; Huhtaniemi, I. Interactions of the GnRH receptor with heterotrimeric G proteins. Front. Neuroendocrinol. 2013, 34, 88–94. [Google Scholar]
- Sakai, T.; Shiraishi, A.; Kawada, T.; Matsubara, S.; Aoyama, M.; Satake, H. Invertebrate gonadotropin-releasing hormone-related peptides and their receptors: An update. Front. Endocrinol. (Lausanne) 2017, 8, 217. [Google Scholar] [CrossRef]
- Semmens, D.C.; Elphick, M.R. The evolution of neuropeptide signalling: Insights from echinoderms. Brief. Funct. Genom. 2017, 16, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Zandawala, M.; Tian, S.; Elphick, M.R. The evolution and nomenclature of GnRH-type and corazonin-type neuropeptide signaling systems. Gen. Comp. Endocrinol. 2018, 264, 64–77. [Google Scholar] [CrossRef]
- Satake, H.; Matsubara, S.; Shiraishi, A.; Yamamoto, T.; Osugi, T.; Sakai, T.; Kawada, T. Neuropeptides, peptide hormones, and their receptors of a tunicate, Ciona intestinalis. Results Probl. Cell Differ. 2019, 68, 107–125. [Google Scholar] [CrossRef]
- Satake, H.; Matsubara, S.; Shiraishi, A.; Yamamoto, T.; Osugi, T.; Sakai, T.; Kawada, T. Peptide receptors and immune-related proteins expressed in the digestive system of a urochordate, Ciona intestinalis. Cell Tissue Res. 2019, 377, 293–308. [Google Scholar] [CrossRef]
- Satoh, N.; Rokhsar, D.; Nishikawa, T. Chordate evolution and the three-phylum system. Proc. Biol. Sci. 2014, 281, 20141729. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, S.; Kawada, T.; Sakai, T.; Aoyama, M.; Osugi, T.; Shiraishi, A.; Satake, H. The significance of Ciona intestinalis as a stem organism in integrative studies of functional evolution of the chordate endocrine, neuroendocrine, and nervous systems. Gen. Comp. Endocrinol. 2016, 227, 101–108. [Google Scholar] [CrossRef]
- Powell, J.F.; Reska-Skinner, S.M.; Prakash, M.O.; Fischer, W.H.; Park, M.; Rivier, J.E. Two new forms of gonadotropin-releasing hormone in a protochordate and the evolutionary implications. Proc. Natl. Acad. Sci. USA 1996, 93, 10461–10464. [Google Scholar] [CrossRef] [Green Version]
- Adams, B.A.; Tello, J.A.; Erchegyi, J.; Warby, C.; Hong, D.J.; Akinsanya, K.O.; Mackie, G.O.; Vale, W.; Rivier, J.E.; Sherwood, N.M. Six novel gonadotropin-releasing hormones are encoded as triplets on each of two genes in the protochordate, Ciona intestinalis. Endocrinology 2003, 144, 1907–1919. [Google Scholar] [CrossRef] [Green Version]
- Hasunuma, I.; Terakado, K. Two novel gonadotropin-releasing hormones (GnRHs) from the urochordate ascidian, Halocynthia roretzi: Implications for the origin of vertebrate GnRH isoforms. Zool. Sci. 2013, 30, 311–318. [Google Scholar] [CrossRef]
- Kawada, T.; Aoyama, M.; Okada, I.; Sakai, T.; Sekiguchi, T.; Ogasawara, M.; Satake, H. A novel inhibitory gonadotropin-releasing hormone-related neuropeptide in the ascidian, Ciona intestinalis. Peptides 2009, 30, 2200–2205. [Google Scholar] [CrossRef]
- Osugi, T.; Sasakura, Y.; Satake, H. The nervous system of the adult ascidian Ciona intestinalis Type A (Ciona robusta): Insights from transgenic animal models. PLoS ONE 2017, 12, e0180227. [Google Scholar] [CrossRef] [Green Version]
- Terakado, K. Induction of gamete release by gonadotropin-releasing hormone in a protochordate, Ciona intestinalis. Gen. Comp. Endocrinol. 2001, 124, 277–284. [Google Scholar] [CrossRef]
- Kusakabe, T.; Mishima, S.; Shimada, I.; Kitajima, Y.; Tsuda, M. Structure, expression, and cluster organization of genes encoding gonadotropin-releasing hormone receptors found in the neural complex of the ascidian Ciona intestinalis. Gene 2003, 322, 77–84. [Google Scholar] [CrossRef]
- Tello, J.A.; Rivier, J.E.; Sherwood, N.M. Tunicate GnRH peptides selectively activate Ciona intestinalis GnRH receptors and the green monkey type II GnRH receptor. Endocrinology 2005, 146, 4061–4073. [Google Scholar] [CrossRef]
- Kusakabe, T.G.; Sakai, T.; Aoyama, M.; Kitajima, Y.; Miyamoto, Y.; Takigawa, T.; Daido, Y.; Fujiwara, K.; Terashima, Y.; Sugiuchi, Y.; et al. A conserved non-reproductive GnRH system in chordates. PLoS ONE 2012, 7, e41955. [Google Scholar] [CrossRef] [Green Version]
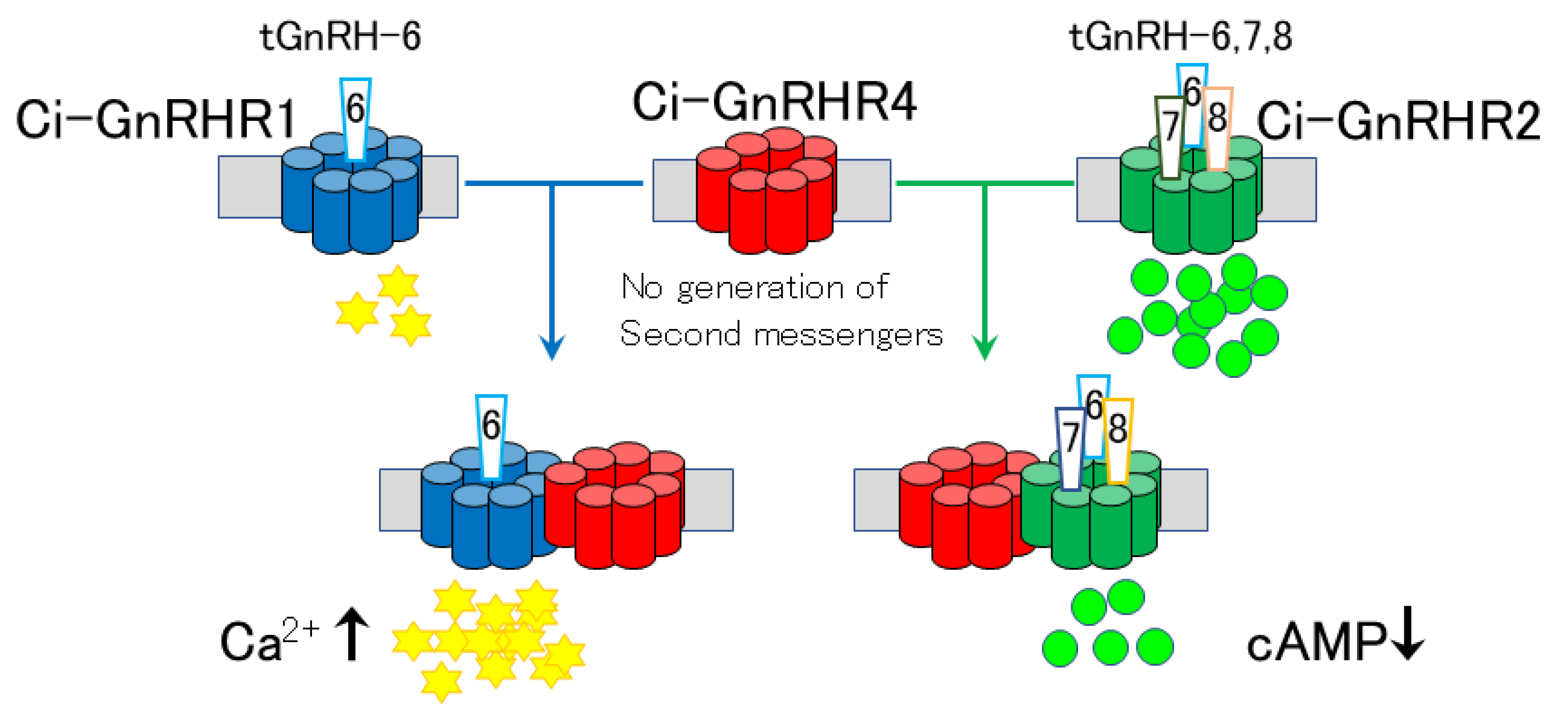
- Sakai, T.; Aoyama, M.; Kusakabe, T.; Tsuda, M.; Satake, H. Functional diversity of signaling pathways through G protein-coupled receptor heterodimerization with a species-specific orphan receptor subtype. Mol. Biol. Evol. 2010, 27, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
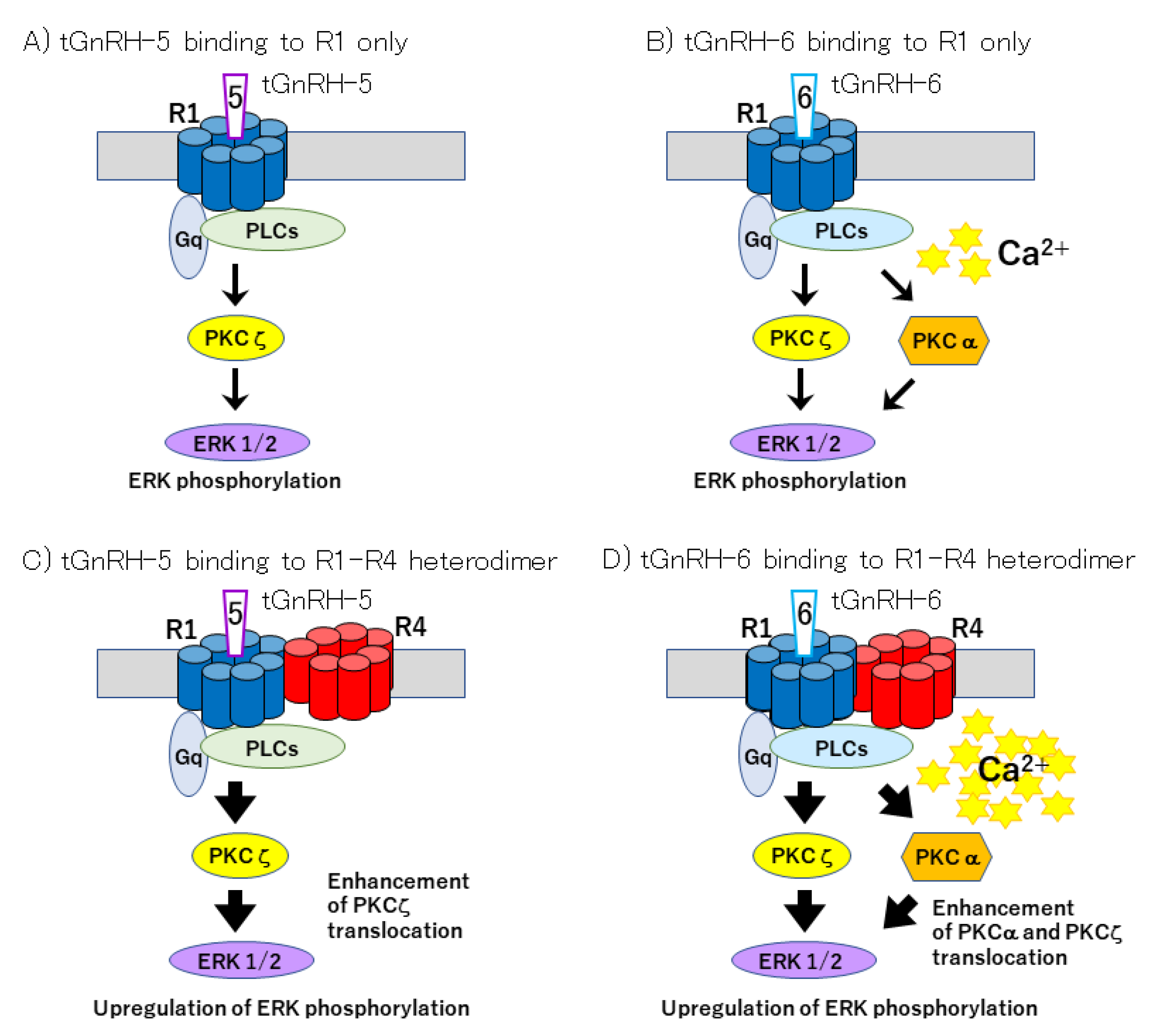
- Sakai, T.; Aoyama, M.; Kawada, T.; Kusakabe, T.; Tsuda, M.; Satake, H. Evidence for differential regulation of GnRH signaling via heterodimerization among GnRH receptor paralogs in the protochordate, Ciona intestinalis. Endocrinology 2012, 153, 1841–1849. [Google Scholar] [CrossRef] [Green Version]
- Satake, H.; Matsubara, S.; Aoyama, M.; Kawada, T.; Sakai, T. GPCR heterodimerization in the reproductive system: Functional regulation and implication for biodiversity. Front. Endocrinol. 2013, 4, 100. [Google Scholar] [CrossRef] [Green Version]
- Kavanaugh, S.I.; Root, A.R.; Sower, S.A. Distribution of gonadotropin-releasing hormone (GnRH) by in situ hybridization in the tunicate Ciona intestinalis. Gen. Comp. Endocrinol. 2005, 141, 76–83. [Google Scholar] [CrossRef]
- Kamiya, C.; Ohta, N.; Ogura, Y.; Yoshida, K.; Horie, T.; Kusakabe, T.G.; Satake, H.; Sasakura, Y. Nonreproductive role of gonadotropin-releasing hormone in the control of ascidian metamorphosis. Dev. Dyn. 2014, 243, 1524–1535. [Google Scholar] [CrossRef]
- Hozumi, A.; Matsunobu, S.; Mita, K.; Treen, N.; Sugihara, T.; Horie, T.; Sakuma, T.; Yamamoto, T.; Shiraishi, A.; Hamada, M.; et al. GABA-induced GnRH release triggers chordate metamorphosis. Curr. Biol. 2020, 30, 1555–1561. [Google Scholar] [CrossRef]
- Roch, G.J.; Tello, J.A.; Sherwood, N.M. At the transition from invertebrates to vertebrates, a novel GnRH-like peptide emerges in amphioxus. Mol. Biol. Evol. 2014, 31, 765–778. [Google Scholar] [CrossRef] [Green Version]
- Tello, J.A.; Sherwood, N.M. Amphioxus: Beginning of vertebrate and end of invertebrate type GnRH receptor lineage. Endocrinology 2009, 150, 2847–2856. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, T.; Suzuki, N.; Fujiwara, N.; Aoyama, M.; Kawada, T.; Sugase, K.; Satake, H. Calcitonin in a protochordate, Ciona intestinalis–The prototype of the vertebrate calcitonin/calcitonin gene-related peptide superfamily. FEBS J. 2009, 276, 4437–4447. [Google Scholar] [CrossRef] [Green Version]
- Rowe, M.L.; Elphick, M.R. The neuropeptide transcriptome of a model echinoderm, the sea urchin Strongylocentrotus purpuratus. Gen. Comp. Endocrinol. 2012, 179, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Zandawala, M.; Beets, I.; Baytemur, E.; Slade, S.E.; Scrivens, J.H.; Elphick, M.R. Urbilaterian origin of paralogous GnRH and corazonin neuropeptide signaling pathways. Sci. Rep. 2016, 6, 28788. [Google Scholar] [CrossRef] [Green Version]
- Tran, K.K.; Jayawardena, B.M.; Elphick, M.R.; Jones, C.E. A gonadotropin-releasing hormone type neuropeptide with a high affinity binding site for copper(ii) and nickel(ii). Metallomics 2019, 11, 404–414. [Google Scholar]
- Tian, S.; Egertová, M.; Elphick, M.R. Functional characterization of paralogous gonadotropin-releasing hormone-type and corazonin-type neuropeptides in an echinoderm. Front. Endocrinol. (Lausanne) 2017, 8, 259. [Google Scholar] [CrossRef] [Green Version]
- Iwakoshi, E.; Takuwa-Kuroda, K.; Fujisawa, Y.; Hisada, M.; Ukena, K.; Tsutsui, K.; Minakata, H. Isolation and characterization of a GnRH-like peptide from Octopus vulgaris. Biochem. Biophys. Res. Commun. 2002, 291, 1187–1193. [Google Scholar] [CrossRef]
- Minakata, H.; Tsutsui, K. Oct-GnRH, the first protostomian gonadotropin-releasing hormone-like peptide and a critical mini-review of the presence of vertebrate sex steroids in molluscs. Gen. Comp. Endocrinol. 2016, 227, 109–114. [Google Scholar] [CrossRef]
- Nuurai, P.; Cummins, S.F.; Natasha, A.; Botwright, N.A.; Sobhon, P. Characterization of an abalone gonadotropin-releasing hormone and its effect on ovarian cell proliferation. Aquaculture 2016, 450, 116–122. [Google Scholar]
- Bose, U.; Suwansa-Ard, S.; Maikaeo, L.; Motti, C.A.; Hall, M.R.; Cummins, S.F. Neuropeptides encoded within a neural transcriptome of the giant triton snail Charonia tritonis, a Crown-of-Thorns Starfish predator. Peptides 2017, 98, 3–14. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, M.A.; Kim, K.S.; Kim, J.W.; Lim, H.K.; Lee, J.S.; Sohn, Y.C. Characterization and spatiotemporal expression of gonadotropin-releasing hormone in the Pacific abalone, Haliotis discus hannai. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 209, 1–9. [Google Scholar] [CrossRef]
- Uengwetwanit, T.; Ponza, P.; Sangsrakru, D.; Wichadakul, D.; Ingsriswang, S.; Leelatanawit, R.; Klinbunga, S.; Tangphatsornruang, S.; Karoonuthaisiri, N. Transcriptome-based discovery of pathways and genes related to reproduction of the black tiger shrimp (Penaeus monodon). Mar. Genom. 2018, 37, 69–73. [Google Scholar] [CrossRef]
- Fodor, I.; Zrínyi, Z.; Horváth, R.; Urbán, P.; Herczeg, R.; Buki, G.; Koene, J.M.; Tsai, P.S.; Pirger, Z. Identification, presence, and possible multifunctional regulatory role of invertebrate gonadotropin-releasing hormone/corazonin molecule in the great pond snail (Lymnaea stagnalis). Gen. Comp. Endocrinol. 2020, 299, 113621. [Google Scholar] [CrossRef]
- Ma, K.Y.; Zhang, S.F.; Wang, S.S.; Qiu, G.F. Molecular cloning and characterization of a gonadotropin-releasing hormone receptor homolog in the Chinese mitten crab, Eriocheir sinensis. Gene 2018, 665, 111–118. [Google Scholar]
- Sharker, M.R.; Sukhan, Z.P.; Kim, S.C.; Lee, W.K.; Kho, K.H. Molecular identification, characterization, and expression analysis of a gonadotropin-releasing hormone receptor (GnRH-R) in pacific abalone, Haliotis discus hannai. Molecules 2020, 25, 2733. [Google Scholar] [CrossRef]
- Kanda, A.; Takahashi, T.; Satake, H.; Minakata, H. Molecular and functional characterization of a novel gonadotropin-releasing-hormone receptor isolated from the common octopus (Octopus vulgaris). Biochem. J. 2006, 395, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Kavanaugh, S.I.; Tsai, P.S. Functional authentication of a novel gastropod gonadotropin-releasing hormone receptor reveals unusual features and evolutionary insight. PLoS ONE 2016, 11, e0160292. [Google Scholar] [CrossRef]
- Nagasawa, K.; Matsubara, S.; Satake, H.; Osada, M. Functional characterization of an invertebrate gonadotropin-releasing hormone receptor in the Yesso scallop Mizuhopecten yessoensis. Gen. Comp. Endocrinol. 2019, 282, 113201. [Google Scholar] [CrossRef]
- Iwakoshi-Ukena, E.; Ukena, K.; Takuwa-Kuroda, K.; Kanda, A.; Tsutsui, K.; Minakata, H. Expression and distribution of octopus gonadotropin-releasing hormone in the central nervous system and peripheral organs of the octopus (Octopus vulgaris) by in situ hybridization and immunohistochemistry. J. Comp. Neurol. 2004, 477, 310–323. [Google Scholar] [CrossRef]
- Tsai, P.S.; Sun, B.; Rochester, J.R.; Wayne, N.L. Gonadotropin-releasing hormone-like molecule is not an acute reproductive activator in the gastropod, Aplysia californica. Gen. Comp. Endocrinol. 2010, 166, 280–288. [Google Scholar] [CrossRef]
- Jung, L.H.; Kavanaugh, S.I.; Sun, B.; Tsai, P.S. Localization of a molluscan gonadotropin-releasing hormone in Aplysia californica by in situ hybridization and immunocytochemistry. Gen. Comp. Endocrinol. 2014, 195, 132–137. [Google Scholar] [CrossRef]
- Sharker, M.R.; Sukhan, Z.P.; Kim, S.C.; Hossen, S.; Cho, Y.; Choi, C.Y.; Kho, K.H. In vivo effects of GnRH peptide on gonadal proliferation and related gene expression of pacific abalone, Haliotis discus hannai. Aquaculture 2020, 531, 735911. [Google Scholar]
- Treen, N.; Itoh, N.; Miura, H.; Kikuchi, I.; Ueda, T.; Takahashi, K.G.; Ubuka, T.; Yamamoto, K.; Sharp, P.J.; Tsutsui, K.; et al. Mollusc gonadotropin-releasing hormone directly regulates gonadal functions: A primitive endocrine system controlling reproduction. Gen. Comp. Endocrinol. 2012, 176, 167–172. [Google Scholar]
- Nagasawa, K.; Oouchi, H.; Itoh, N.; Takahashi, K.G.; Osada, M. In Vivo administration of scallop GnRH-like peptide influences on gonad development in the yesso scallop, Patinopecten yessoensis. PLoS ONE 2015, 10, e0129571. [Google Scholar]
- Nagasawa, K.; Muroi, K.; Thitiphuree, T.; Minegishi, Y.; Itoh, N.; Osada, M. Cloning of invertebrate gonadotropin-releasing hormone receptor (GnRHR)-like gene in Yesso scallop, Patinopecten yessoensis. Agri Gene 2017, 3, 46–56. [Google Scholar] [CrossRef]
- Shiraishi, A.; Okuda, T.; Miyasaka, N.; Osugi, T.; Okuno, Y.; Inoue, J.; Satake, H. Repertoires of G protein-coupled receptors for Ciona-specific neuropeptides. Proc. Natl. Acad. Sci. USA 2019, 116, 7847–7856. [Google Scholar] [CrossRef] [Green Version]
- Sasakura, Y.; Yoshida, K.; Treen, N. Genome editing of the ascidian Ciona intestinalis with TALE nuclease. Methods Mol. Biol. 2017, 1630, 235–245. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| GnRH | ||||
|---|---|---|---|---|
| Vertebrata | ||||
| Species | Scientific Name | Peptide Name | Amino Acid Sequence | Receptors and Signaling Pathway |
| Human | Homo sapiens | GnRH1 | pQ--HWSYGLRPGa | GnRHR1 (Ca2+) |
| GnRH2 | pQ--HWSHGWYPGa | Non-functional? | ||
| Guinea pig | Cavia porcellus | GnRH1 | pQ--YWSYGVRPGa | GnRHR1 (Ca2+) |
| Sea bass | Dicentrarchus labrax | GnRH3 | pQ--HWSYGWLPGa | dlGnRHR-II-1b (Ca2+, cAMP) |
| Lamprey | Petromyzon marinus | l-GnRH-I | pQ--HYSLEWKPGa | lGnRH-R-1 (Ca2+, cAMP), -3(Ca2+) |
| l-GnRH-II | pQ--HWSHGWFPGa | lGnRH-R-1 (Ca2+, cAMP) | ||
| l-GnRH-III | pQ--HWSHDWKPGa | lGnRH-R-1 (Ca2+, cAMP), -2(Ca2+) | ||
| Urochordata | ||||
| Tunicate | Chelyosoma productum | tGnRH-1 | pQ--HWSDYFKPGa | N.D. |
| tGnRH-2 | pQ--HWSLCHAPGa | N.D. | ||
| tGnRH-3 | pQ--HWSYEFMPGa | Ci-GnRHR-3 (cAMP) | ||
| Ciona intestinalis | tGnRH-4 | pQ--HWSNQLTPGa | Ci-GnRHR-2 (cAMP) | |
| tGnRH-5 | pQ--HWSYEYMPGa | Ci-GnRHR-3 (cAMP) | ||
| tGnRH-6 | pQ--HWSKGYSPGa | Ci-GnRHR-1 (Ca2+, cAMP), -2 (cAMP) | ||
| tGnRH-7 | pQ--HWSYALSPGa | Ci-GnRHR-2 (cAMP) | ||
| tGnRH-8 | pQ--HWSLALSPGa | Ci-GnRHR-2 (cAMP) | ||
| Ciona savignyi | tGnRH-9 | pQ--HWSNKLAPGa | N.D. | |
| Ciona intestinalis | Ci-GnRH-X | pQ—HWSNWWIPGAP | Partial antagonist for Ci-GnRHR-1, 3 | |
| GYNGa | ||||
| Halocynthia roretzi | tGnRH-10 | pQ--HWSYGFSPGa | N.D. | |
| tGnRH-11 | pQ--HWSYGFLPGa | N.D. | ||
| Cephalochordata | ||||
| Amphioxus | Branchiostoma floridae | Amph.GnRHv | pQE-HWQYGHWYa | unidentified |
| Amph.GnRH-like | pQILCARAFTYTHTWa | Bf-GnRHR-3 (Ca2+) | ||
| Echinodermata | ||||
| Sea urchin | Strongylocentrotus purpuratus | Sp-GnRHP | pQVHHRFSGWRPGa | N.D. |
| Starfish | Asterias rubens | Ar-GnRH | pQIHYKNPGWGPGa | ArGnRHR (Ca2+) |
| Protostomia | ||||
| Mollusca | ||||
| Octopus | Octopus vulgaris | oct-GnRH | pQNYHFSNGWHPGa | Oct-GnRHR (Ca2+) |
| Cuttlefish | Sepia officinalis | oct-GnRH | pQNYHFSNGWHPGa | N.D. |
| Swordtip squid | Loligo edulis | oct-GnRH | pQNYHFSNGWHPGa | N.D. |
| Oyster | Crassostrea gigas | Cg-GnRH | pQNYHFSNGWQPa | N.D. |
| Yesso scallop | Mizuhopecten (Patinopecten) yessoensis | my-invGnRH (py-GnRH) | pQNFHYSNGWQPa | my-invGnRHR (Ca2+) |
| Sea hare | Aplysia californica | ap-GnRH | pQNYHFSNGWYAa | ap-GnRHR (Ca2+) |
| Owl limpet | Lottia gigantean | Lg-GnRH | pQHYHFSNGWKSa | N.D. |
| Giant triton snail | Charonia tritonis | Ctr-GnRH | pQNYHYSNGWHPa | N.D. |
| Abalone | Haliotis asinina | Has-GnRH | pQNYHFSNGWYPGa | N.D. |
| Haliotis laevigata | Hlae-GnRH | pQNYHFSNGWHAa | N.D. | |
| Pacific abalone | Haliotis discus hannai | Hdh-GnRH | pQNYHFSNGWYAa | Hdh-GnRHR (No functional assay) |
| Annelida | ||||
| Marine worm | Capitella teleta | Ca-GnRH | pQAYHFSHGWFPa | N.D. |
| Leech | Helobdella robusta | Hr-GnRH | pQSIHFSRSWQPa | N.D. |
| Crustacea | ||||
| Mitten crab | Eriocheir Sinensis | N.D. | N.D. | Es-GnRHR (transcriptome) |
| Black tiger shrimp | Penaeus monodon | Pm-GnRH | (transcriptome) | Pm-GnRHR (transcriptome) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakai, T.; Yamamoto, T.; Matsubara, S.; Kawada, T.; Satake, H. Invertebrate Gonadotropin-Releasing Hormone Receptor Signaling and Its Relevant Biological Actions. Int. J. Mol. Sci. 2020, 21, 8544. https://doi.org/10.3390/ijms21228544
Sakai T, Yamamoto T, Matsubara S, Kawada T, Satake H. Invertebrate Gonadotropin-Releasing Hormone Receptor Signaling and Its Relevant Biological Actions. International Journal of Molecular Sciences. 2020; 21(22):8544. https://doi.org/10.3390/ijms21228544
Chicago/Turabian StyleSakai, Tsubasa, Tatsuya Yamamoto, Shin Matsubara, Tsuyoshi Kawada, and Honoo Satake. 2020. "Invertebrate Gonadotropin-Releasing Hormone Receptor Signaling and Its Relevant Biological Actions" International Journal of Molecular Sciences 21, no. 22: 8544. https://doi.org/10.3390/ijms21228544
APA StyleSakai, T., Yamamoto, T., Matsubara, S., Kawada, T., & Satake, H. (2020). Invertebrate Gonadotropin-Releasing Hormone Receptor Signaling and Its Relevant Biological Actions. International Journal of Molecular Sciences, 21(22), 8544. https://doi.org/10.3390/ijms21228544