Tangeretin Ameliorates Glucose-Induced Podocyte Injury through Blocking Epithelial to Mesenchymal Transition Caused by Oxidative Stress and Hypoxia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
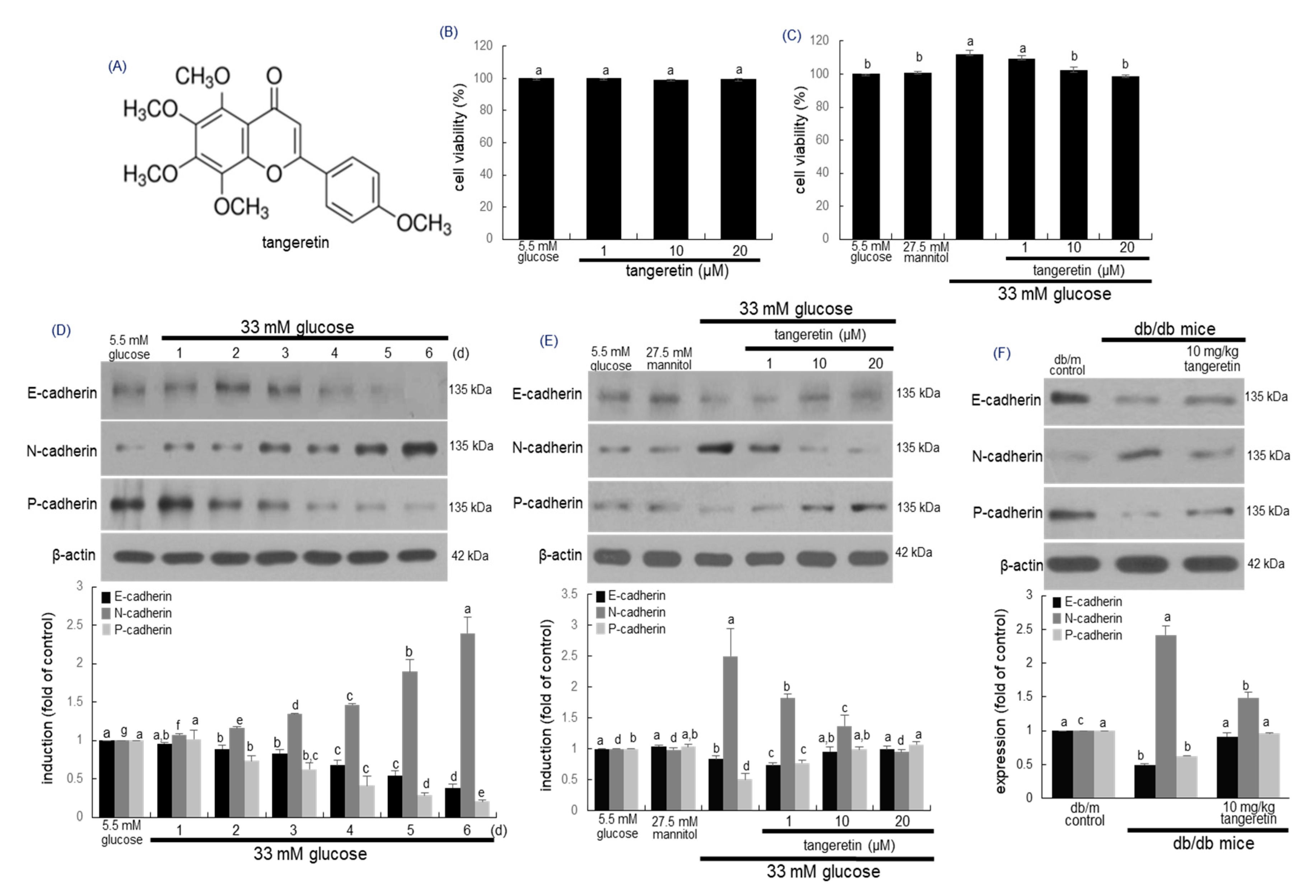
2.1. Inhibitory Effects of Tangeratein on Glucose-Induced Podocyte EMT
2.2. Inhibition of Formation of Myofibroblasts by Tangeretin
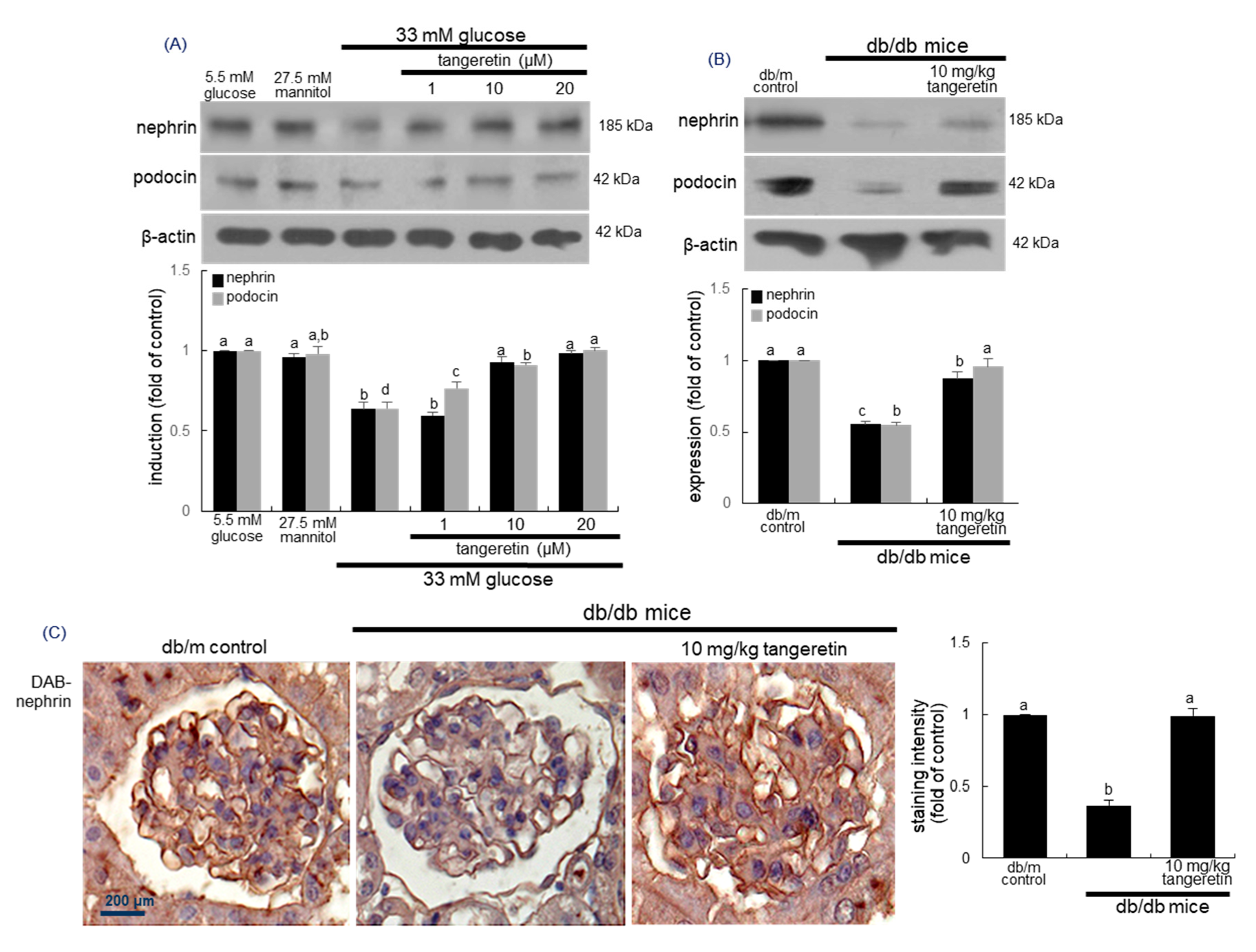
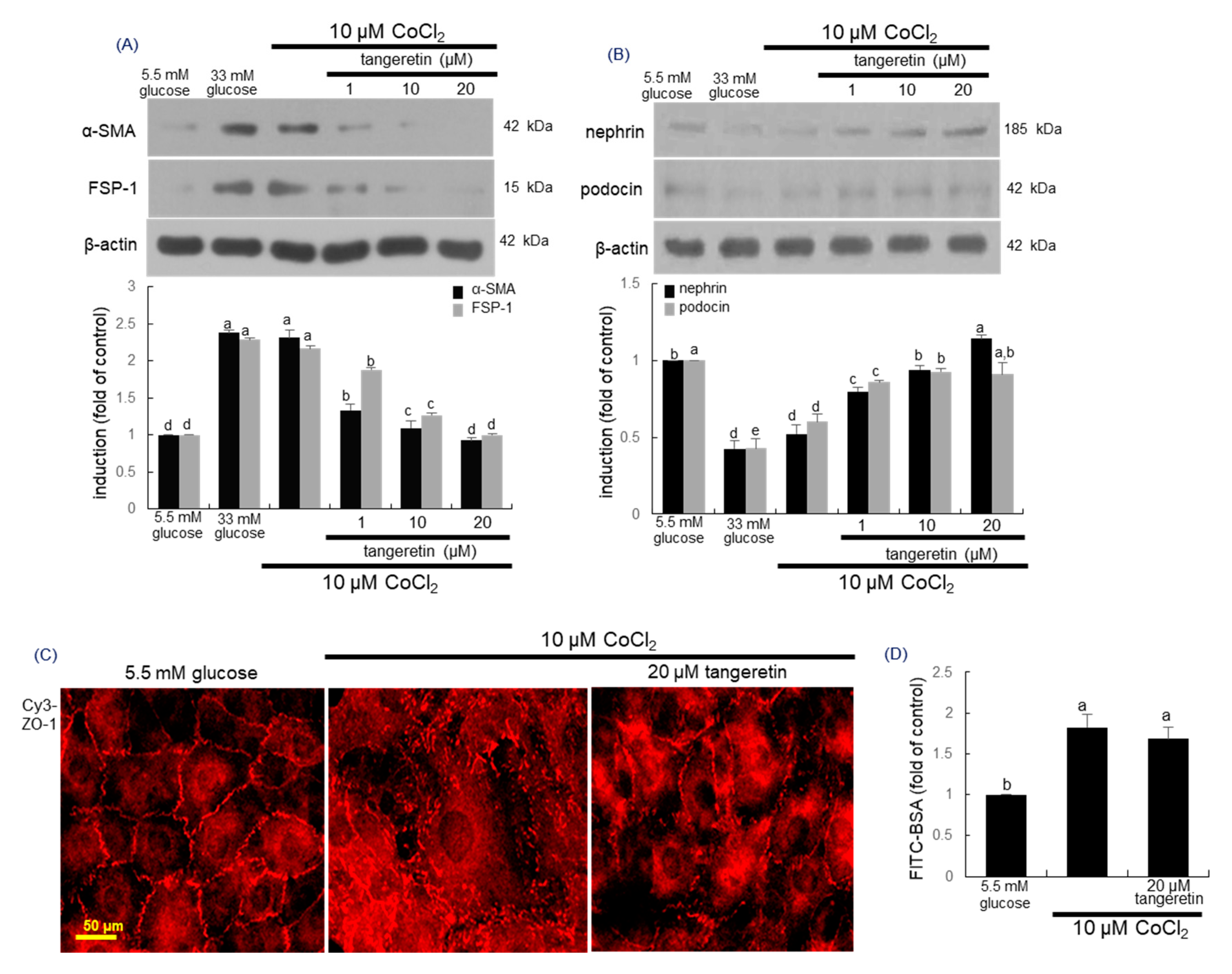
2.3. Inhibition of Loss of Slit Diaphragm Proteins by Tangeretin
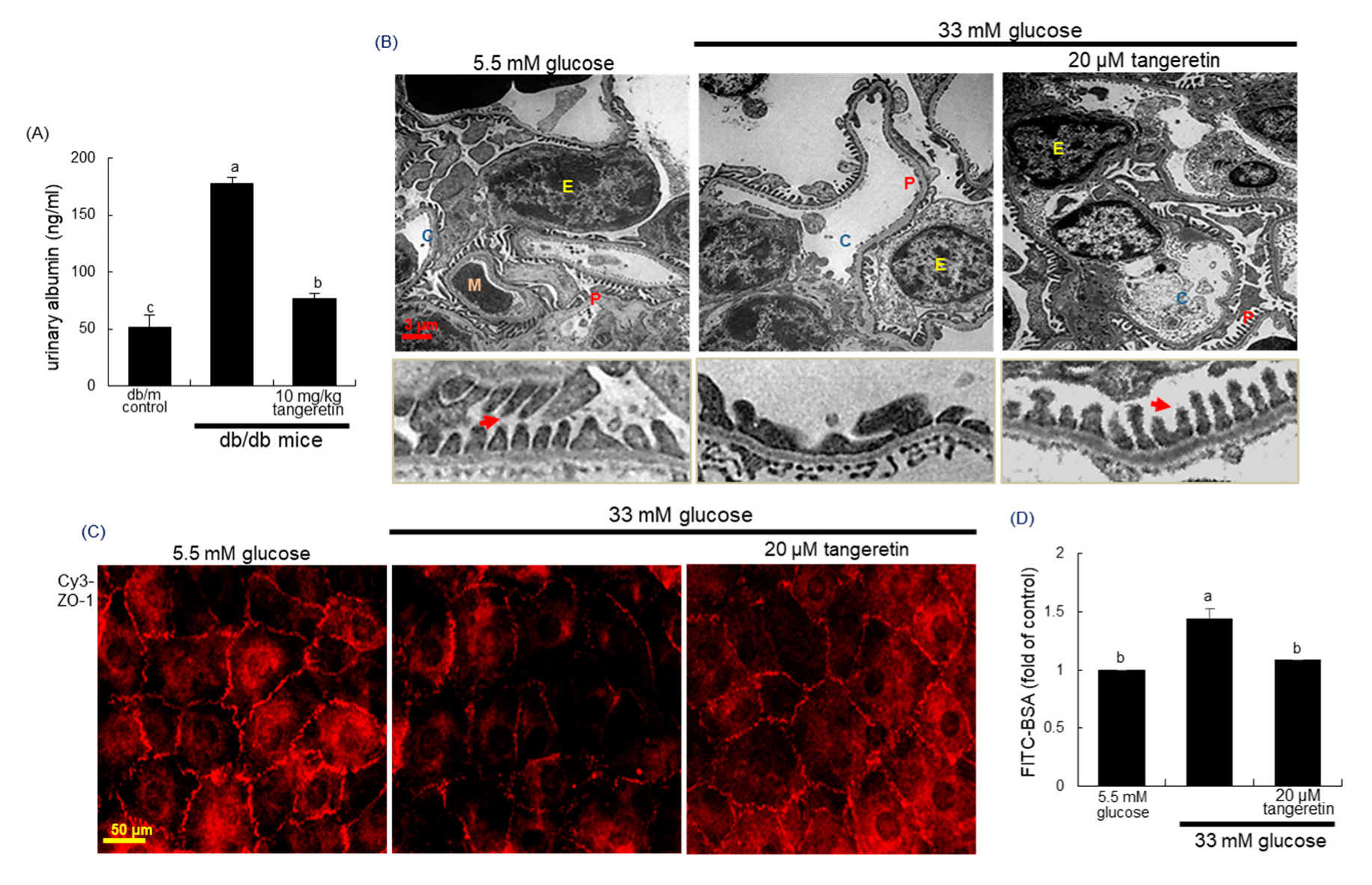
2.4. Blockade of Podocyte Injury by Tangeretin
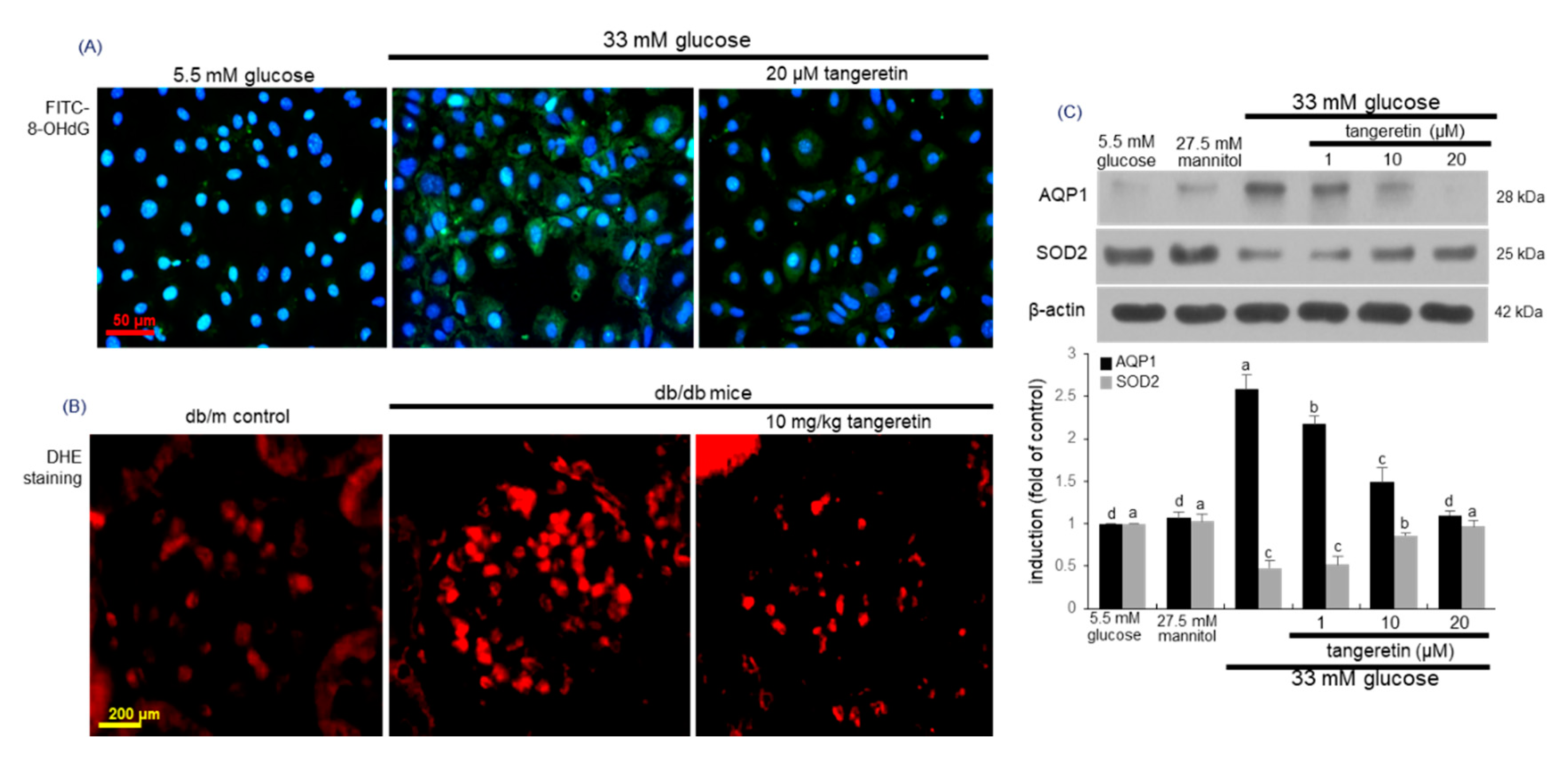
2.5. Effects of Tangeretin on Glucose-Induced Oxidative Stress in Podocytes
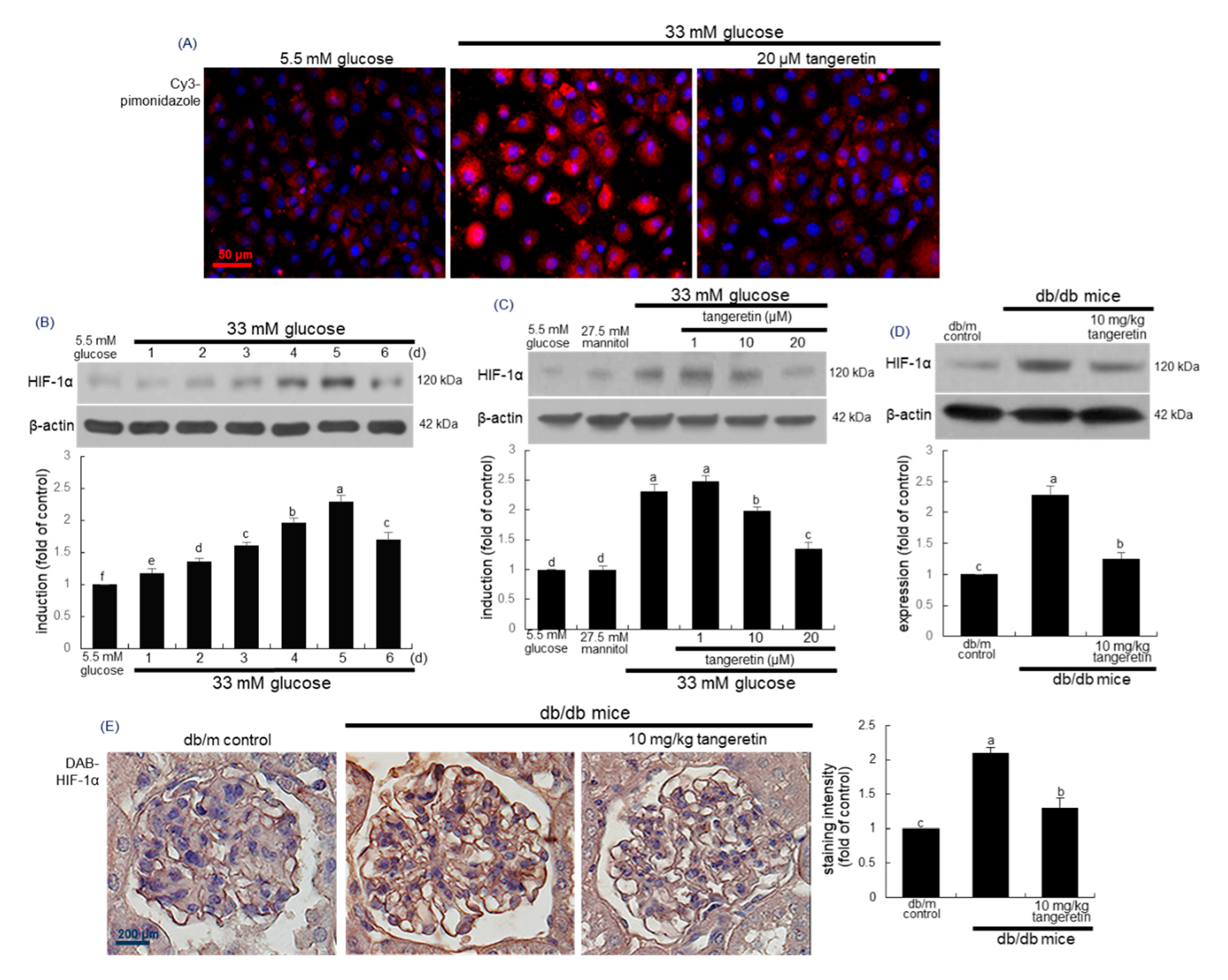
2.6. Suppressive Effects of Tangeretin on Hypoxia-Induced Podocyte Injury
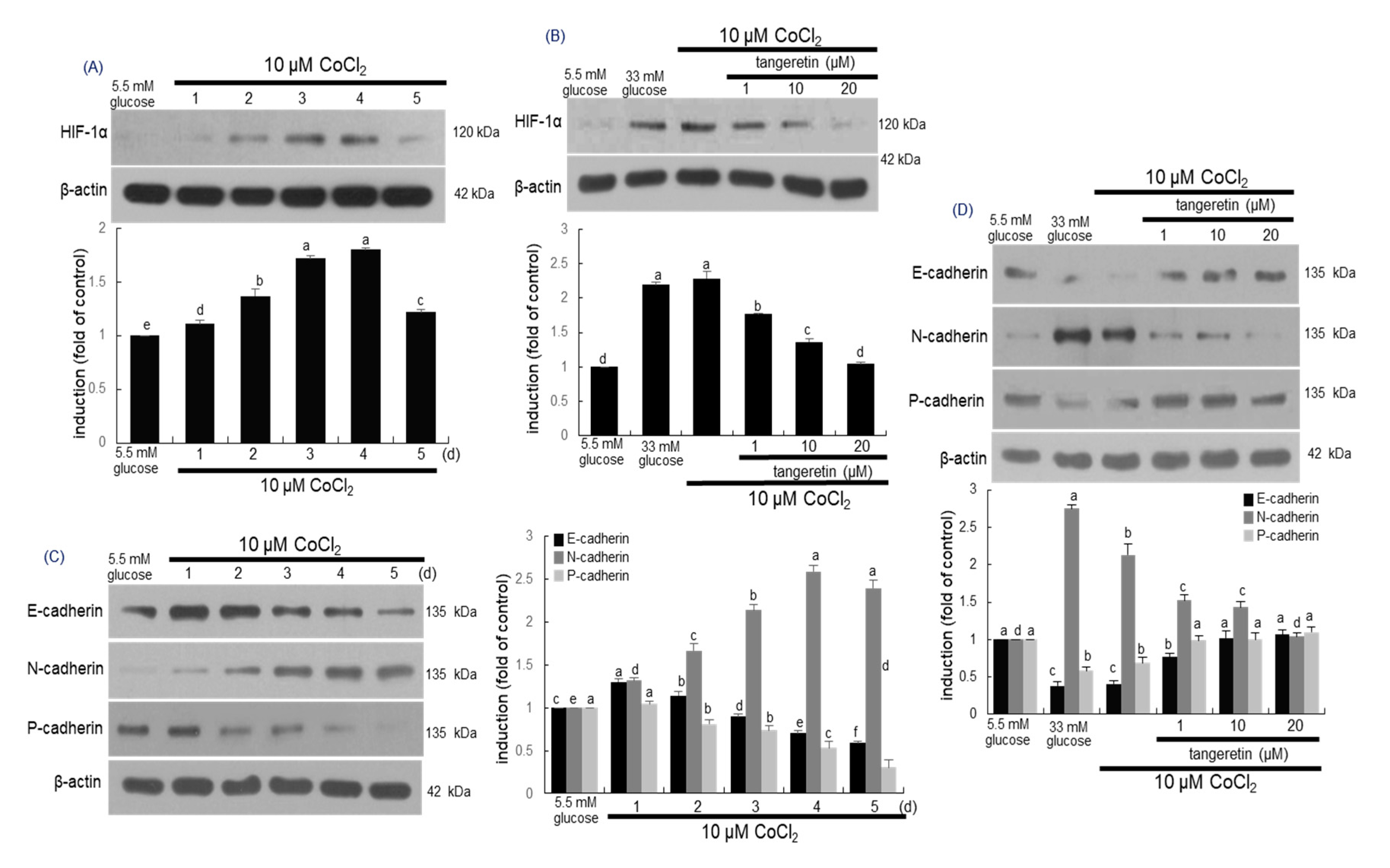
2.7. Inhibitory Effects of Tangeretin on Hypoxia and EMT Induced by CoCl2
2.8. Inhibition of Hypoxia-Induced Disruption of Adherens Junction by Tangeretin
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Murine Kidney Podocyte Culture
4.3. Western Blot Analysis
4.4. In Vivo Mouse Experiments
4.5. Immunohistochemical Staining
4.6. Urinary Albumin Excretion
4.7. TEM Analysis
4.8. Immunocytochemical Staining
4.9. Albumin Permeability Assay
4.10. Measurement of ROS Production
4.11. Data Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AQP1 | aquaporin-1 |
| CoCl2 | cobalt chloride |
| DN | diabetic nephropathy |
| EMT | epithelial-to-mesenchymal transition |
| FSP-1 | fibroblast-specific protein-1 |
| HIF-1α | hypoxia inducible factor-1α |
| 8-OHdG | 8-hydroxy-2-deoxy guanosine |
| α-SMA | α-smooth muscle actin |
| SOD2 | superoxide dismutase 2 |
| ZO-1 | zonula occludens-1 |
References
- Wiggins, R.C. The spectrum of podocytopathies: A unifying view of glomerular diseases. Kidney Int. 2007, 71, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Davies, D.J.; Messina, A.; Thumwood, C.M.; Ryan, G.B. Glomerular podocytic injury in protein overload proteinuria. Pathology 1985, 17, 412–419. [Google Scholar] [CrossRef]
- Pavenstadt, H.; Kriz, W.; Kretzler, M. Cell biology of the glomerular podocyte. Physiol. Rev. 2003, 83, 253–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asanuma, K.; Mundel, P. The role of podocytes in glomerular pathobiology. Clin. Exp. Nephrol. 2003, 7, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Benzing, T. Signaling at the slit diaphragm. J. Am. Soc. Nephrol. 2004, 15, 1382–1391. [Google Scholar] [CrossRef] [Green Version]
- Denhez, B.; Geraldes, P. Regulation of nephrin phosphorylation in diabetes and chronic kidney injury. Adv. Exp. Med. Biol. 2017, 966, 149–161. [Google Scholar] [PubMed]
- Marshall, S.M. The podocyte: A potential therapeutic target in diabetic nephropathy? Curr. Pharm. Des. 2007, 13, 2713–2720. [Google Scholar] [CrossRef]
- Li, Y.; Kang, Y.S.; Dai, C.; Kiss, L.P.; Wen, X.; Liu, Y. Epithelial-to-mesenchymal transition is a potential pathway leading to podocyte dysfunction and proteinuria. Am. J. Pathol. 2008, 172, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Iwano, M.; Toyoda, M.; Kimura, K.; Harada, K.; Nakatani, Y.; Yoshimoto, S.; Asai, O.; Akai, Y.; Suzuki, D.; et al. Epithelial-mesenchymal transition as an explanation for podocyte depletion in diabetic nephropathy. Am. J. Kidney Dis. 2009, 54, 653–664. [Google Scholar] [CrossRef]
- Ying, Q.; Wu, G. Molecular mechanisms involved in podocyte EMT and concomitant diabetic kidney diseases: An update. Ren. Fail. 2017, 39, 474–483. [Google Scholar] [CrossRef] [Green Version]
- Dai, H.; Liu, Q.; Liu, B. Research progress on mechanism of podocyte depletion in diabetic nephropathy. J. Diabetes Res. 2017, 2017, 2615286. [Google Scholar] [CrossRef] [PubMed]
- Giaccia, A.J.; Simon, M.C.; Johnson, R. The biology of hypoxia: The role of oxygen sensing in development, normal function, and disease. Genes Dev. 2004, 18, 2183–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mvafagh, S.; Crook, S.; Vo, K. Regulation of hypoxia-inducible factor-1α by reactive oxygen species: New developments in an old debate. J. Cell. Biochem. 2015, 116, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, H.; Inagi, R. Stress signal network between hypoxia and ER stress in chronic kidney disease. Front. Physiol. 2017, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Sada, K.; Nishikawa, T.; Kukidome, D.; Yoshinaga, T.; Kajihara, N.; Sonoda, K.; Senokuchi, T.; Motoshima, H.; Matsumura, T.; Araki, E. Hyperglycemia induces cellular hypoxia through production of mitochondrial ROS followed by suppression of aquaporin-1. PLoS ONE 2016, 11, e0158619. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Colgan, S.P.; Shelley, C.S. Hypoxia: The force that drives chronic kidney disease. Clin. Med. Res. 2016, 14, 15–39. [Google Scholar] [CrossRef] [Green Version]
- Rosenberger, C.; Khamaisi, M.; Abassi, Z.; Shilo, V.; Weksler-Zangen, S.; Goldfarb, M.; Shina, A.; Zibertrest, F.; Eckardt, K.U.; Rosen, S.; et al. Adaptation to hypoxia in the diabetic rat kidney. Kidney Int. 2008, 73, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Higgins, D.F.; Kimura, K.; Bernhardt, W.M.; Shrimanker, N.; Akai, Y.; Hohenstein, B.; Saito, Y.; Johnson, R.S.; Kretzler, M.; Cohen, C.D.; et al. Hypoxia promotes fibrogenesis in vivo via HIF-1 stimulation of epithelial-to-mesenchymal transition. J. Clin. Investig. 2007, 117, 3810–3820. [Google Scholar] [CrossRef]
- Haase, V.H. Oxygen regulates epithelial-to-mesenchymal transition: Insights into molecular mechanisms and relevance to disease. Kidney Int. 2009, 76, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Reiser, J.; Gupta, V.; Kistler, A.D. Toward the development of podocyte-specific drugs. Kidney Int. 2010, 77, 662–668. [Google Scholar] [CrossRef] [Green Version]
- Xiang, C.; Xiao, X.; Jiang, B.; Zhou, M.; Zhang, Y.; Li, H.; Hu, Z. Epigallocatechin-3-gallate protects from high glucose induced podocyte apoptosis via suppressing endoplasmic reticulum stress. Mol. Med. Rep. 2017, 16, 6142–6147. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.X.; Wang, Q.J.; Li, H.; Huang, Q. Silencing of USP22 suppresses high glucose-induced apoptosis, ROS production and inflammation in podocytes. Mol. Biosyst. 2016, 12, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.K.; Park, S.H.; Kim, Y.H.; Lee, E.J.; Antika, L.D.; Kim, D.Y.; Choi, Y.J.; Kang, Y.H. Chrysin ameliorates podocyte injury and slit diaphragm protein loss via inhibition of the PERK-eIF2α-ATF-CHOP pathway in diabetic mice. Acta Pharmacol. Sin. 2017, 38, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Behzad, S.; Habtemariam, S.; Ahmed, T.; Daglia, M.; Nabavi, S.M.; Sobarzo-Sanchez, E.; Nabavi, S.F. Neuroprotective effects of citrus fruit-derived flavonoids, nobiletin and tangeretin in alzheimer’s and parkinson’s disease. C.N.S. Neurol. Disord. Drug Targets. 2017, 16, 387–397. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Ali, M.S.; Moraes, L.A.; Sage, T.; Lewis, K.R.; Jones, C.I. Tangeretin regulates platelet function through inhibition of phosphoinositide 3-kinase and cyclic nucleotide signaling. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2740–2749. [Google Scholar] [CrossRef] [Green Version]
- Arivazhagan, L.; Sorimuthu, P.S. Tangeretin, a citrus pentamethoxyflavone, exerts cytostatic effect via P53/P21 up-regulation and suppresses metastasis in 7,12-dimethylbenz(alpha)anthracene-induced rat mammary carcinoma. J. Nutr. Biochem. 2014, 25, 1140–1153. [Google Scholar] [CrossRef]
- Vinay, T.; Sivakumar, S.; Inho, H. Molecular and cellular response of co-cultured cells toward cobalt chloride (CoCl2)-induced hypoxia. ACS Omega 2019, 4, 20882–20893. [Google Scholar]
- Jorge, S.; María, C. The use of cobalt chloride as a chemical hypoxia model. J. Appl. Toxicol. 2019, 39, 556–570. [Google Scholar]
- Meran, S.; Steadman, R. Fibroblasts and myofibroblasts in renal fibrosis. Int. J. Exp. Pathol. 2011, 92, 158–167. [Google Scholar] [CrossRef]
- Wolf, G.; Ziyadeh, F.N. Cellular and molecular mechanisms of proteinuria in diabetic nephropathy. Nephron Physiol. 2007, 106, 26–31. [Google Scholar] [CrossRef]
- Shono, A.; Tsukaguchi, H.; Yaoita, E.; Nameta, M.; Kurihara, H.; Qin, X.S.; Yamamoto, T.; Doi, T. Podocin participates in the assembly of tight junctions between foot processes in nephrotic podocytes. J. Am. Soc. Nephrol. 2007, 18, 2525–2533. [Google Scholar] [CrossRef] [PubMed]
- Athanasios, V.; Thomais, V.; Constantinos, F. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar]
- Brandt, L.E.; Bohn, A.A.; Charles, J.B.; Ehrhart, E.J. Localization of canine, feline, and mouse renal membrane proteins. Vet. Pathol. 2012, 49, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Suwimol, K.; Sujatha, V.; Garry, R.B.; Larry, W.O. Manganese superoxide dismutase modulates hypoxia inducible factor-1 alpha induction via superoxide. Cancer Res. 2008, 68, 2781–2788. [Google Scholar]
- Sasabe, E.; Yang, Z.; Ohno, S.; Yamamoto, T. Reactive oxygen species produced by the knockdown of manganese-superoxide dismutase up-regulate hypoxia-inducible factor-1alpha expression in oral squamous cell carcinoma cells. Free Radic. Biol. Med. 2010, 48, 1321–1329. [Google Scholar] [CrossRef]
- Rana, N.K.; Singh, P.; Koch, B. CoCl2 simulated hypoxia induce cell proliferation and alter the expression pattern of hypoxia associated genes involved in angiogenesis and apoptosis. Biol. Res. 2019, 52, 12. [Google Scholar] [CrossRef] [Green Version]
- Dennis, J.M.; Witting, P.K. Protective role for antioxidants in acute kidney disease. Nutrients 2017, 9, 718. [Google Scholar] [CrossRef] [Green Version]
- Cervellati, F.; Cervellati, C.; Romani, A.; Cremonini, E.; Sticozzi, C.; Belmonte, G.; Pessina, F.; Valacchi, G. Hypoxia induces cell damage via oxidative stress in retinal epithelial cells. Free Radic. Res. 2014, 48, 303–312. [Google Scholar] [CrossRef]
- Reiser, J.; Kriz, W.; Kretzler, M.; Mundel, P.P. The glomerular slit diaphragm is a modified adherens junction. J. Am. Soc. Nephrol. 2000, 11, 1–8. [Google Scholar]
- Avila-Carrasco, L.; Majano, P.; Sánchez-Toméro, J.A.; Selgas, R.; López-Cabrera, M.; Aguilera, A.; Mateo, G.G. Natural plants compounds as modulators of epithelial-to-mesenchymal transition. Front. Pharmacol. 2019, 10, 715. [Google Scholar] [CrossRef] [Green Version]
- Kawachi, H.; Fukusumi, Y. New insight into podocyte slit diaphragm, a therapeutic target of proteinuria. Clin. Exp. Nephrol. 2020, 24, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.Y.; Kang, M.K.; Park, S.H.; Lee, E.J.; Kim, Y.H.; Oh, H.; Choi, Y.J.; Kang, Y.H. Eucalyptol ameliorates Snail1/β-catenin-dependent diabetic disjunction of renal tubular epithelial cells and tubulointerstitial fibrosis. Oncotarget 2017, 8, 106190–106205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Liu, Y. Wnt/β-catenin signalling and podocyte dysfunction in proteinuric kidney disease. Nat. Rev. Nephrol. 2015, 11, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.A.; Welsh, G.I.; Saleem, M.A.; Menon, R.K. Molecular and cellular events Mediating glomerular Podocyte dysfunction and depletion in diabetes mellitus. Front. Endocrinol. Lausanne 2014, 5, 151. [Google Scholar]
- Choi, D.; Kim, C.L.; Kim, J.E.; Mo, J.S.; Jeong, H.S. Hesperetin inhibit EMT in TGF-β treated podocyte by regulation of mTOR pathway. Biochem. Biophys. Res. Commun. 2020, 528, 154–159. [Google Scholar] [CrossRef]
- Hill, C.; Li, J.; Liu, D.; Conforti, F.; Brereton, C.J.; Yao, L.; Zhou, Y.; Alzetani, A.; Chee, S.J.; Marshall, B.G.; et al. Autophagy inhibition-mediated epithelial-mesenchymal transition augments local myofibroblast differentiation in pulmonary fibrosis. Cell Death Dis. 2019, 10, 591. [Google Scholar] [CrossRef]
- Wang, X.; Gao, Y.; Tian, N.; Wang, T.; Shi, Y.; Xu, J.; Wu, B. Astragaloside IV inhibits glucose-induced epithelial-mesenchymal transition of podocytes through autophagy enhancement via the SIRT–NF-κB p65 axis. Sci. Rep. 2019, 9, 323. [Google Scholar] [CrossRef] [Green Version]
- Möller, M.N.; Cuevasanta, E.; Orrico, F.; Lopez, A.C.; Thomson, L.; Denicola, A. Diffusion and transport of reactive species across cell membranes. Adv. Exp. Med. Biol. 2019, 1127, 3–19. [Google Scholar]
- Kitada, M.; Xu, J.; Ogura, Y.; Monno, I.; Koya, D. manganese superoxide dismutase dysfunction and the pathogenesis of kidney disease. Front. Physiol. 2020, 11, 755. [Google Scholar] [CrossRef]
- Jens, R.N.; Yassuo, I.; Charles, K.; Joseph, R.W. Interactions between hyperglycemia and hypoxia: Implications for diabetic retinopathy. Diabetes 2004, 53, 2931–2938. [Google Scholar]
- Sergiu, B.C. Impaired hypoxia-inducible factor (HIF) regulation by hyperglycemia. J. Mol. Med. Berl. 2014, 92, 1025–1034. [Google Scholar]
- Schiffer, T.A.; Friederich-Persson, M. Mitochondrial reactive oxygen species and kidney hypoxia in the development of diabetic nephropathy. Front. Physiol. 2017, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danli, W.; Patricia, Y. Induction and testing of hypoxia in cell culture. J. Vis. Exp. 2011, 54, 2899. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, M.-K.; Kim, S.-I.; Oh, S.Y.; Na, W.; Kang, Y.-H. Tangeretin Ameliorates Glucose-Induced Podocyte Injury through Blocking Epithelial to Mesenchymal Transition Caused by Oxidative Stress and Hypoxia. Int. J. Mol. Sci. 2020, 21, 8577. https://doi.org/10.3390/ijms21228577
Kang M-K, Kim S-I, Oh SY, Na W, Kang Y-H. Tangeretin Ameliorates Glucose-Induced Podocyte Injury through Blocking Epithelial to Mesenchymal Transition Caused by Oxidative Stress and Hypoxia. International Journal of Molecular Sciences. 2020; 21(22):8577. https://doi.org/10.3390/ijms21228577
Chicago/Turabian StyleKang, Min-Kyung, Soo-Il Kim, Su Yeon Oh, Woojin Na, and Young-Hee Kang. 2020. "Tangeretin Ameliorates Glucose-Induced Podocyte Injury through Blocking Epithelial to Mesenchymal Transition Caused by Oxidative Stress and Hypoxia" International Journal of Molecular Sciences 21, no. 22: 8577. https://doi.org/10.3390/ijms21228577
APA StyleKang, M. -K., Kim, S. -I., Oh, S. Y., Na, W., & Kang, Y. -H. (2020). Tangeretin Ameliorates Glucose-Induced Podocyte Injury through Blocking Epithelial to Mesenchymal Transition Caused by Oxidative Stress and Hypoxia. International Journal of Molecular Sciences, 21(22), 8577. https://doi.org/10.3390/ijms21228577