Peptide-Mediated Gene Transfer into Marine Purple Photosynthetic Bacteria

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
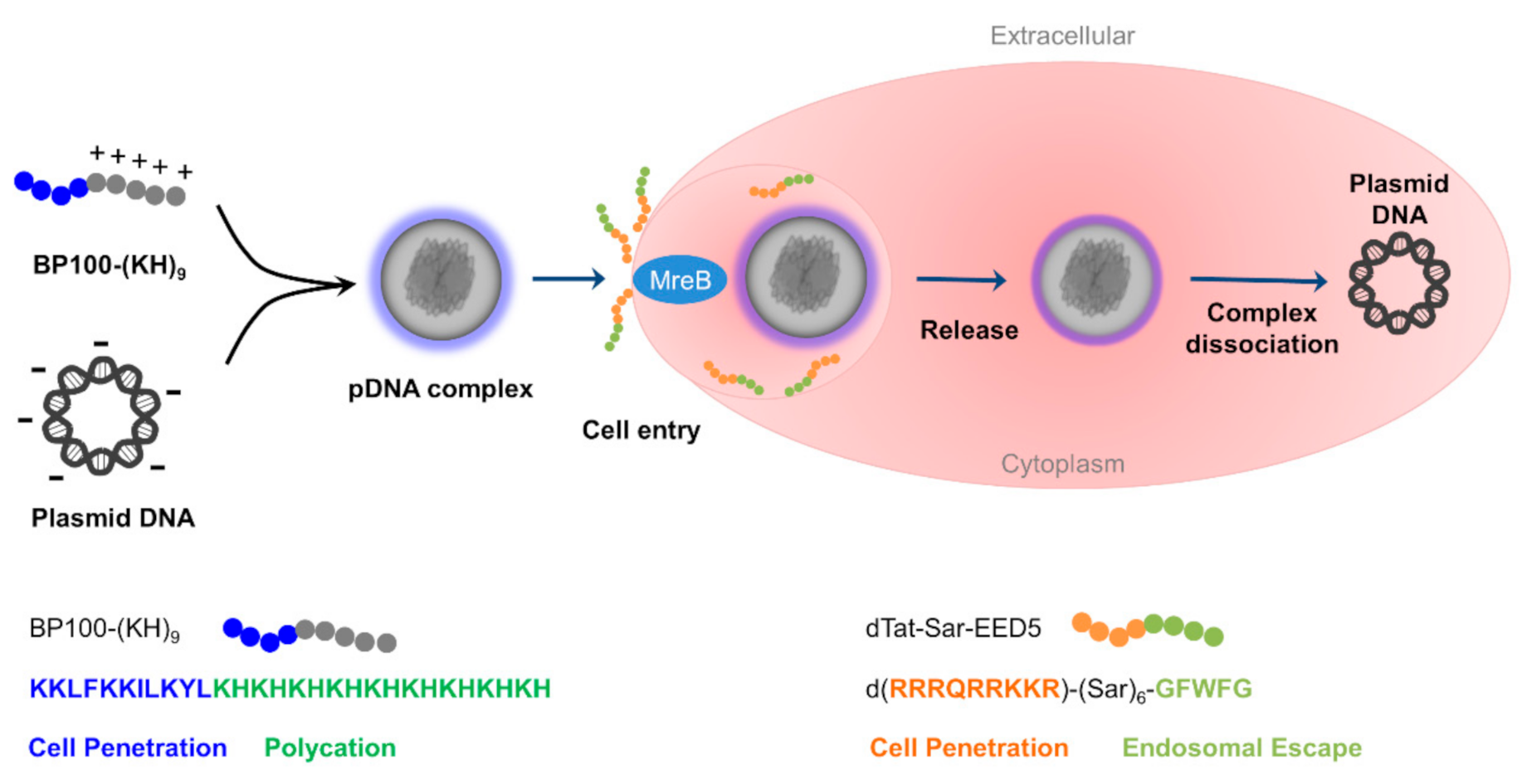
2.1. Characterization of Peptide-Plasmid DNA Complex
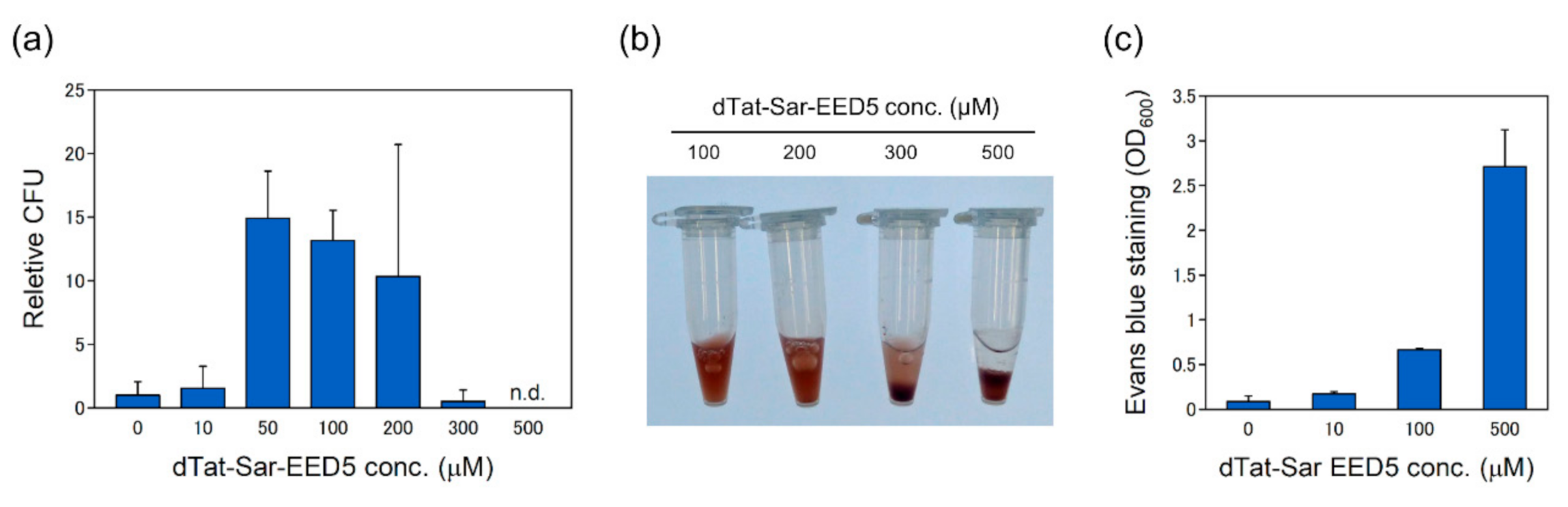
2.2. Effects of dTat-Sar-EED5 Peptide on pDNA Delivery and Cell Viability
2.3. Optimization of Plasmid DNA Delivery
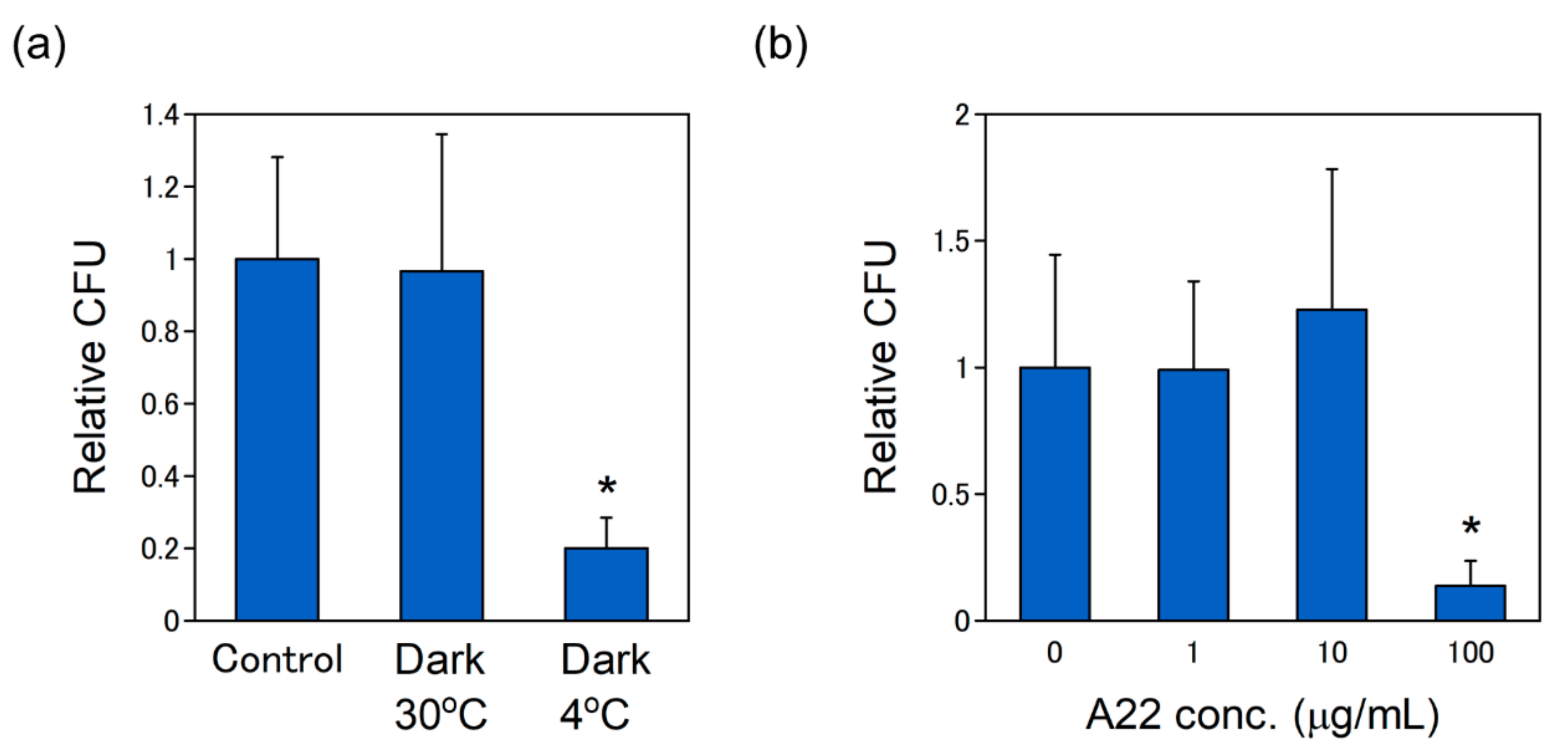
2.4. The pDNA Delivery via MreB-Mediated Energy Dependent Pathway
2.5. The dTat-Sar-EED Mediated pDNA Delivery into R. sulfidophilum
2.6. The pDNA Delivery into E. coli Cells
2.7. Mechanism of the Peptide Functions against Bacterial Cells
3. Materials and Methods
3.1. Culture Conditions
3.2. Preparation of Peptide-Plasmid DNA Complex
3.3. Transformation of R. sulfidophilum and E. coli Cells
3.4. Evans Blue Staining
3.5. Formation and Stability of pDNA Complexes
3.6. Dynamic Light Scattering (DLS) Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lorenz, M.G.; Wackernagel, W. Bacterial gene-transfer by natural genetic-transformation in the environment. Microbiol. Rev. 1994, 58, 563–602. [Google Scholar] [CrossRef] [Green Version]
- Veening, J.W.; Blokesch, M. Interbacterial predation as a strategy for DNA acquisition in naturally competent bacteria. Nat. Rev. Microbiol. 2017, 15, 621–629. [Google Scholar] [CrossRef]
- Mandel, M.; Higa, A. Calcium-dependent bacteriophage DNA infection. J. Mol. Biol. 1970, 53, 159–162. [Google Scholar] [CrossRef]
- Dagert, M.; Ehrlich, S.D. Prolonged incubation in calcium chloride improves the competence of Escherichia coli cells. Gene 1979, 6, 23–28. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Fornari, C.S.; Kaplan, S. Genetic-transformation of Rhodopseudomonas sphaeroides by plasmid DNA. J. Bacteriol. 1982, 152, 89–97. [Google Scholar]
- Russo, R.; Panangala, V.S.; Wood, R.R.; Klesius, P.H. Chemical and electroporated transformation of Edwardsiella ictaluri using three different plasmids. FEMS Microbiol. Lett. 2009, 298, 105–110. [Google Scholar] [CrossRef]
- Asif, A.; Mohsin, H.; Tanvir, R.; Rehman, Y. Revisiting the mechanisms involved in calcium chloride induced bacterial transformation. Front. Microbiol. 2017, 8, 2169. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Jessee, J.; Bloom, F.R. Plasmid transformation of Escherichia coli and other bacteria. Methods Enzymol. 1991, 204, 63–113. [Google Scholar] [CrossRef] [PubMed]
- Inaba, H.; Matsuura, K. Peptide nanomaterials designed from natural supramolecular systems. Chem. Rec. 2019, 19, 843–858. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K. Synthetic approaches to construct viral capsid-like spherical nanomaterials. Chem. Commun. (Camb.) 2018, 54, 8944–8959. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, M.; Kodama, Y.; Yoshizumi, T.; Sudesh, K.; Numata, K. Rapid and efficient gene delivery into plant cells using designed peptide carriers. Biomacromolecules 2013, 14, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Numata, K.; Ohtani, M.; Yoshizumi, T.; Demura, T.; Kodama, Y. Local gene silencing in plants via synthetic dsRNA and carrier peptide. Plant. Biotechnol. J. 2014, 12, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.; Motoda, Y.; Watanabe, S.; Sofiman Othman, A.; Kigawa, T.; Kodama, Y.; Numata, K. Intracellular delivery of proteins via fusion peptides in intact plants. PLoS ONE 2016, 11, e0154081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Itami, J.; Oikawa, K.; Motoda, Y.; Kigawa, T.; Numata, K. Native protein delivery into rice callus using ionic complexes of protein and cell-penetrating peptides. PLoS ONE 2019, 14, e0214033. [Google Scholar] [CrossRef] [Green Version]
- Chuah, J.A.; Odahara, M.; Kodama, Y.; Miyamoto, T.; Tsuchiya, K.; Motoda, Y.; Kigawa, T.; Numata, K. Synthetic peptide-induced internalization of biomolecules into various plant and algal cells via micropinocytosis. bioRxiv 2019. [Google Scholar] [CrossRef]
- Lonn, P.; Kacsinta, A.D.; Cui, X.S.; Hamil, A.S.; Kaulich, M.; Gogoi, K.; Dowdy, S.F. Enhancing endosomal escape for intracellular delivery of macromolecular biologic therapeutics. Sci. Rep. 2016, 6, 32301. [Google Scholar] [CrossRef]
- Miyamoto, T.; Tsuchiya, K.; Numata, K. Dual peptide-based gene delivery system for the efficient transfection of plant callus cells. Biomacromolecules 2020. [Google Scholar] [CrossRef]
- Higuchi-Takeuchi, M.; Numata, K. Marine purple photosynthetic bacteria as sustainable microbial production hosts. Front. Bioeng. Biotechnol. 2019, 7, 258. [Google Scholar] [CrossRef]
- Idi, A.; Nor, M.H.M.; Wahab, M.F.A.; Ibrahim, Z. Photosynthetic bacteria: An eco-friendly and cheap tool for bioremediation. Rev. Environ. Sci. Bio-Technol. 2015, 14, 271–285. [Google Scholar] [CrossRef]
- Gallon, J.R. N2 fixation in phototrophs: Adaptation to a specialized way of life. Plant Soil 2001, 230, 39–48. [Google Scholar] [CrossRef]
- Jensen, P.R. Natural products and the gene cluster revolution. Trends Microbiol. 2016, 24, 968–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, P.R.; Fenical, W. Strategies for the discovery of secondary metabolites from marine bacteria: Ecological perspectives. Annu. Rev. Microbiol. 1994, 48, 559–584. [Google Scholar] [CrossRef] [PubMed]
- Satpute, S.K.; Banat, I.M.; Dhakephalkar, P.K.; Banpurkar, A.G.; Chopade, B.A. Biosurfactants, bioemulsifiers and exopolysaccharides from marine microorganisms. Biotechnol. Adv. 2010, 28, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Higuchi-Takeuchi, M.; Morisaki, K.; Toyooka, K.; Numata, K. Synthesis of high-molecular-weight polyhydroxyalkanoates by marine photosynthetic purple bacteria. PLoS ONE 2016, 11, e0160981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi-Takeuchi, M.; Numata, K. Acetate-inducing metabolic states enhance polyhydroxyalkanoate production in marine purple non-sulfur bacteria under aerobic conditions. Front. Bioeng. Biotechnol. 2019, 7, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foong, C.P.; Higuchi-Takeuchi, M.; Numata, K. Optimal iron concentrations for growth-associated polyhydroxyalkanoate biosynthesis in the marine photosynthetic purple bacterium Rhodovulum sulfidophilum under photoheterotrophic condition. PLoS ONE 2019, 14, e0212654. [Google Scholar] [CrossRef] [Green Version]
- Maeda, I.; Miyasaka, H.; Umeda, F.; Kawase, M.; Yagi, K. Maximization of hydrogen production ability in high-density suspension of Rhodovulum sulfidophilum cells using intracellular poly(3-hydroxybutyrate) as sole substrate. Biotechnol. Bioeng. 2003, 81, 474–481. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Umekage, S. Extracellular nucleic acids of the marine bacterium Rhodovulum sulfidophilum and recombinant RNA production technology using bacteria. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Foong, C.P.; Higuchi-Takeuchi, M.; Malay, A.D.; Oktaviani, N.A.; Thagun, C.; Numata, K. A marine photosynthetic microbial cell factory as a platform for spider silk production. Commun. Biol. 2020, 3, 357. [Google Scholar] [CrossRef]
- Numata, K. How to define and study structural proteins as biopolymer materials. Polym. J. 2020, 52, 1043–1056. [Google Scholar] [CrossRef]
- Higuchi-Takeuchi, M.; Morisaki, K.; Numata, K. Method for the facile transformation of marine purple photosynthetic bacteria using chemically competent cells. Microbiologyopen 2020, 9, e00953. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M., 2nd; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- El-Andaloussi, S.; Jarver, P.; Johansson, H.J.; Langel, U. Cargo-dependent cytotoxicity and delivery efficacy of cell-penetrating peptides: A comparative study. Biochem. J. 2007, 407, 285–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Annu. Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [Green Version]
- Conner, S.D.; Schmid, S.L. Regulated portals of entry into the cell. Nature 2003, 422, 37–44. [Google Scholar] [CrossRef]
- Jones, L.J.; Carballido-Lopez, R.; Errington, J. Control of cell shape in bacteria: Helical, actin-like filaments in Bacillus subtilis. Cell 2001, 104, 913–922. [Google Scholar] [CrossRef]
- Van den Ent, F.; Amos, L.A.; Lowe, J. Prokaryotic origin of the actin cytoskeleton. Nature 2001, 413, 39–44. [Google Scholar] [CrossRef]
- Gital, Z.; Dye, N.A.; Reisenauer, A.; Wachi, M.; Shapiro, L. MreB actin-mediated segregation of a specific region of a bacterial chromosome. Cell 2005, 120, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Bean, G.J.; Flickinger, S.T.; Westler, W.M.; McCully, M.E.; Sept, D.; Weibel, D.B.; Amann, K.J. A22 disrupts the bacterial actin cytoskeleton by directly binding and inducing a low-affinity state in MreB. Biochemistry 2009, 48, 4852–4857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, R.A.; Errington, J. Control of cell morphogenesis in bacteria: Two distinct ways to make a rod-shaped cell. Cell 2003, 113, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Lonhienne, T.G.; Sagulenko, E.; Webb, R.I.; Lee, K.C.; Franke, J.; Devos, D.P.; Nouwens, A.; Carroll, B.J.; Fuerst, J.A. Endocytosis-like protein uptake in the bacterium Gemmata obscuriglobus. Proc. Natl. Acad. Sci. USA 2010, 107, 12883–12888. [Google Scholar] [CrossRef] [Green Version]
- Rajarao, G.K.; Nekhotiaeva, N.; Good, L. Peptide-mediated delivery of green fluorescent protein into yeasts and bacteria. FEMS Microbiol. Lett. 2002, 215, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Toyohara, D.; Yokoi, Y.; Inoue, G.; Muraoka, T.; Mori, T. Abiotic factors promote cell penetrating peptide permeability in Enterobacteriaceae models. Front. Microbiol. 2019, 10, 2534. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, K.; Islam, M.M.; Horii, Y.; Yoshizumi, T.; Numata, K. Screening of a cell-penetrating peptide library in Escherichia coli: Relationship between cell penetration efficiency and cytotoxicity. Acs Omega 2018, 3, 16489–16499. [Google Scholar] [CrossRef] [Green Version]
- Strahl, H.; Errington, J. Bacterial membranes: Structure, domains, and function. Annu. Rev. Microbiol. 2017, 71, 519–538. [Google Scholar] [CrossRef]
- Mullineaux, C.W.; Liu, L.N. Membrane dynamics in phototrophic bacteria. Annu. Rev. Microbiol. 2020, 74, 633–654. [Google Scholar] [CrossRef]
- Lizier, M.; Sarra, P.G.; Cauda, R.; Lucchini, F. Comparison of expression vectors in Lactobacillus reuteri strains. FEMS Microbiol. Lett. 2010, 308, 8–15. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Higuchi-Takeuchi, M.; Miyamoto, T.; Foong, C.P.; Goto, M.; Morisaki, K.; Numata, K. Peptide-Mediated Gene Transfer into Marine Purple Photosynthetic Bacteria. Int. J. Mol. Sci. 2020, 21, 8625. https://doi.org/10.3390/ijms21228625
Higuchi-Takeuchi M, Miyamoto T, Foong CP, Goto M, Morisaki K, Numata K. Peptide-Mediated Gene Transfer into Marine Purple Photosynthetic Bacteria. International Journal of Molecular Sciences. 2020; 21(22):8625. https://doi.org/10.3390/ijms21228625
Chicago/Turabian StyleHiguchi-Takeuchi, Mieko, Takaaki Miyamoto, Choon Pin Foong, Mami Goto, Kumiko Morisaki, and Keiji Numata. 2020. "Peptide-Mediated Gene Transfer into Marine Purple Photosynthetic Bacteria" International Journal of Molecular Sciences 21, no. 22: 8625. https://doi.org/10.3390/ijms21228625
APA StyleHiguchi-Takeuchi, M., Miyamoto, T., Foong, C. P., Goto, M., Morisaki, K., & Numata, K. (2020). Peptide-Mediated Gene Transfer into Marine Purple Photosynthetic Bacteria. International Journal of Molecular Sciences, 21(22), 8625. https://doi.org/10.3390/ijms21228625