Hiding in Plain Sight: Virtually Unrecognizable Memory Phenotype CD8+ T cells


{kind=link}
{kind=link}
Abstract
:1. Introduction
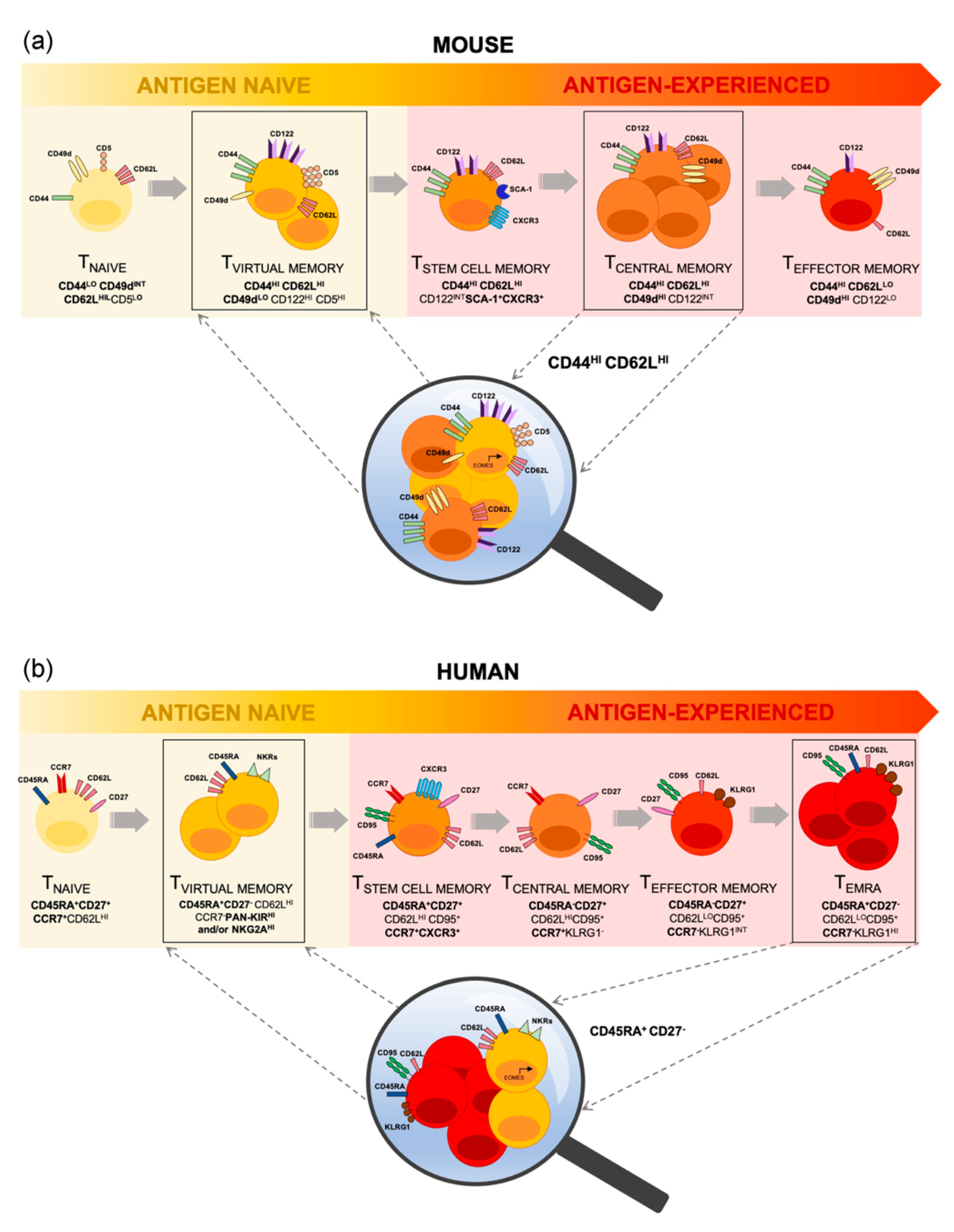
2. Conflation of Mouse TVM Cells with Conventional TMEM Cells
3. TVM Cells Are Contained within the Human TEMRA Cell Population
4. Heterogeneity within the TVM Cell Compartment
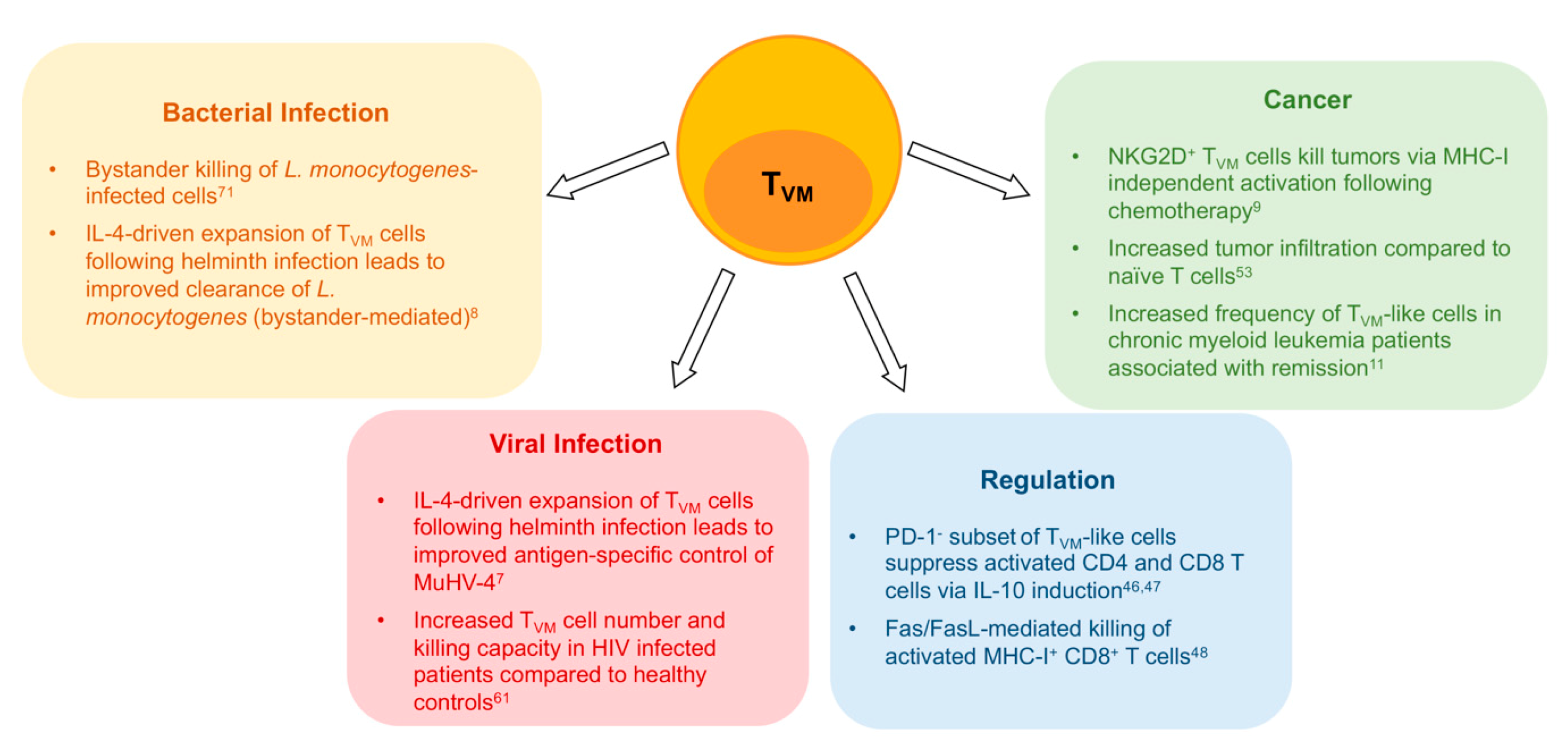
5. Heightened TCR Reactivity and Cytokine Sensitivity Are Key TVM Cell Characteristics
6. NKR Expression as Markers of Functionality and Senescence on TVM Cells
7. Transcriptional Regulation of TVM Development and Function
8. Epigenetic Regulation of TVM Cells and Age-Related T Cell Dysfunction
9. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Sallusto, F.; Lenig, D.; Förster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nat. Cell Biol. 1999, 401, 708–712. [Google Scholar] [CrossRef]
- Quinn, K.M.; Fox, A.; Harland, K.L.; Russ, B.E.; Li, J.; Nguyen, T.H.O.; Loh, L.; Olshanksy, M.; Naeem, H.; Tsyganov, K.; et al. Age-Related Decline in Primary CD8+ T Cell Responses Is Associated with the Development of Senescence in Virtual Memory CD8+ T Cells. Cell Rep. 2018, 23, 3512–3524. [Google Scholar] [CrossRef] [PubMed]
- Quinn, K.M.; Hussain, T.; Kraus, F.; Formosa, L.E.; Lam, W.K.; Dagley, M.J.; Saunders, E.C.; Assmus, L.M.; Wynne-Jones, E.; Loh, L.; et al. Metabolic characteristics of CD8+ T cell subsets in young and aged individuals are not predictive of functionality. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Decman, V.; Laidlaw, B.J.; Doering, T.A.; Leng, J.; Ertl, H.C.J.; Goldstein, D.R.; Wherry, E.J. Defective CD8 T Cell Responses in Aged Mice Are Due to Quantitative and Qualitative Changes in Virus-Specific Precursors. J. Immunol. 2012, 188, 1933–1941. [Google Scholar] [CrossRef] [PubMed]
- Haluszczak, C.; Akue, A.D.; Hamilton, S.E.; Johnson, L.D.; Pujanauski, L.; Teodorovic, L.; Jameson, S.C.; Kedl, R.M. The antigen-specific CD8+ T cell repertoire in unimmunized mice includes memory phenotype cells bearing markers of homeostatic expansion. J. Exp. Med. 2009, 206, 435–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-Y.; Hamilton, S.E.; Akue, A.D.; Hogquist, K.A.; Jameson, S.C. Virtual memory CD8 T cells display unique functional properties. Proc. Natl. Acad. Sci. USA 2013, 110, 13498–13503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolot, M.; Dougall, A.M.; Chetty, A.; Javaux, J.; Chen, T.; Xiao, X.; Machiels, B.; Selkirk, M.E.; Maizels, R.M.; Hokke, C.; et al. Helminth-induced IL-4 expands bystander memory CD8+ T cells for early control of viral infection. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.S.; Mohrs, K.; Szaba, F.M.; Kummer, L.W.; Leadbetter, E.A.; Mohrs, M. Virtual memory CD8 T cells expanded by helminth infection confer broad protection against bacterial infection. Mucosal Immunol. 2018, 12, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Waschke, B.C.; Woolaver, R.A.; Chen, S.M.Y.; Chen, Z.; Wang, J.H. MHC class I-independent activation of virtual memory CD8 T cells induced by chemotherapeutic agent-treated cancer cells. Cell. Mol. Immunol. 2020, 1–12. [Google Scholar] [CrossRef]
- Rufer, N.; Brümmendorf, T.H.; Kolvraa, S.; Bischoff, C.; Christensen, K.; Wadsworth, L.; Schulzer, M.; Lansdorp, P.M. Telomere Fluorescence Measurements in Granulocytes and T Lymphocyte Subsets Point to a High Turnover of Hematopoietic Stem Cells and Memory T Cells in Early Childhood. J. Exp. Med. 1999, 190, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Cayssials, E.; Jacomet, F.; Piccirilli, N.; Lefèvre, L.; Roy, L.; Guilhot, F.; Chomel, J.; Leleu, X.; Gombert, J.; Herbelin, A.; et al. Sustained treatment-free remission in chronic myeloid leukaemia is associated with an increased frequency of innate CD8(+) T-cells. Br. J. Haematol. 2019, 186, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Kaech, S.M.; Ahmed, R. Memory CD8+ T cell differentiation: Initial antigen encounter triggers a developmental program in naïve cells. Nat. Immunol. 2001, 2, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wherry, E.J.; Ahmed, R. Memory CD8 T-Cell Differentiation during Viral Infection. J. Virol. 2004, 78, 5535–5545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harty, J.T.; Badovinac, V.P. Shaping and reshaping CD8+ T-cell memory. Nat. Rev. Immunol. 2008, 8, 107–119. [Google Scholar] [CrossRef]
- Kieper, W.C.; Jameson, S.C. Homeostatic expansion and phenotypic conversion of naïve T cells in response to self peptide/MHC ligands. Proc. Natl. Acad. Sci. USA 1999, 96, 13306–13311. [Google Scholar] [CrossRef] [Green Version]
- Goldrath, A.W.; Bogatzki, L.Y.; Bevan, M.J. Naive T Cells Transiently Acquire a Memory-like Phenotype during Homeostasis-Driven Proliferation. J. Exp. Med. 2000, 192, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Sosinowski, T.; White, J.T.; Cross, E.W.; Haluszczak, C.; Marrack, P.; Gapin, L.; Kedl, R.M. CD8α+Dendritic CellTransPresentation of IL-15 to Naive CD8+T Cells Produces Antigen-Inexperienced T Cells in the Periphery with Memory Phenotype and Function. J. Immunol. 2013, 190, 1936–1947. [Google Scholar] [CrossRef] [Green Version]
- Chiu, B.-C.; Martin, B.E.; Stolberg, V.R.; Chensue, S.W. Cutting Edge: Central Memory CD8 T Cells in Aged Mice Are Virtual Memory Cells. J. Immunol. 2013, 191, 5793–5796. [Google Scholar] [CrossRef] [Green Version]
- Judge, A.D.; Zhang, X.; Fujii, H.; Surh, C.D.; Sprent, J. Interleukin 15 Controls both Proliferation and Survival of a Subset of Memory-Phenotype CD8+ T Cells. J. Exp. Med. 2002, 196, 935–946. [Google Scholar] [CrossRef]
- White, J.T.; Cross, E.W.; Burchill, M.A.; Danhorn, T.; McCarter, M.D.; Rosen, H.R.; O’Connor, B.; Kedl, R.M. Virtual memory T cells develop and mediate bystander protective immunity in an IL-15-dependent manner. Nat. Commun. 2016, 7, 11291. [Google Scholar] [CrossRef] [Green Version]
- Jameson, S.C. Maintaining the norm: T-cell homeostasis. Nat. Rev. Immunol. 2002, 2, 547–556. [Google Scholar] [CrossRef]
- Surh, C.D.; Sprent, J. Homeostasis of Naive and Memory T Cells. Immunity 2008, 29, 848–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murali-Krishna, K.; Lau, L.L.; Sambhara, S.; Lemonnier, F.; Altman, J.; Ahmed, R. Persistence of Memory CD8 T Cells in MHC Class I-Deficient Mice. Science 1999, 286, 1377–1381. [Google Scholar] [CrossRef] [PubMed]
- Boyman, O.; Cho, J.-H.; Tan, J.T.; Surh, C.D.; Sprent, J. A major histocompatibility complex class I–dependent subset of memory phenotype CD8+ cells. J. Exp. Med. 2006, 203, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Kurepa, Z.; Su, J.; Forman, J. Memory phenotype of CD8+ T cells in MHC class Ia-deficient mice. J. Immunol. 2003, 170, 5414–5420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacomet, F.; Cayssials, E.; Basbous, S.; Levescot, A.; Piccirilli, N.; Desmier, D.; Robin, A.; Barra, A.; Giraud, C.; Guilhot, F.; et al. Evidence for eomesodermin-expressing innate-like CD8+KIR/NKG2A+T cells in human adults and cord blood samples. Eur. J. Immunol. 2015, 45, 1926–1933. [Google Scholar] [CrossRef]
- Galletti, G.; De Simone, G.; Mazza, E.M.C.; Puccio, S.; Mezzanotte, C.; Bi, T.M.; Davydov, A.N.; Metsger, M.; Scamardella, E.; Alvisi, G.; et al. Two subsets of stem-like CD8+ memory T cell progenitors with distinct fate commitments in humans. Nat. Immunol. 2020, 1–11. [Google Scholar] [CrossRef]
- Geginat, J.; Lanzavecchia, A.; Sallusto, F. Proliferation and differentiation potential of human CD8+ memory T-cell subsets in response to antigen or homeostatic cytokines. Blood 2003, 101, 4260–4266. [Google Scholar] [CrossRef]
- White, J.T.; Cross, E.W.; Kedl, R.M. Antigen-inexperienced memory CD8+ T cells: Where they come from and why we need them. Nat. Rev. Immunol. 2017, 17, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Appay, V.; Dunbar, P.R.; Callan, M.; Klenerman, P.; Gillespie, G.M.A.; Papagno, L.; Ogg, G.S.; King, A.; Lechner, F.; Spina, C.A.; et al. Memory CD8+ T cells vary in differentiation phenotype in different persistent virus infections. Nat. Med. 2002, 8, 379–385. [Google Scholar] [CrossRef]
- Henson, S.M.; Lanna, A.; Riddell, N.E.; Franzese, O.; Macaulay, R.; Griffiths, S.J.; Puleston, D.J.; Watson, A.S.; Simon, A.K.; Tooze, S.A.; et al. p38 signaling inhibits mTORC1-independent autophagy in senescent human CD8+ T cells. J. Clin. Investig. 2014, 124, 4004–4016. [Google Scholar] [CrossRef] [PubMed]
- Zinser, M.; Highton, A.J.; Kurioka, A.; Kronsteiner, B.; Hagel, J.; Leng, T.; Marchi, E.; Phetsouphanh, C.; Willberg, C.B.; Dunachie, S.J.; et al. Human MAIT cells show metabolic quiescence with rapid glucose-dependent upregulation of granzyme B upon stimulation. Immunol. Cell Biol. 2018, 96, 666–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, P.; Morris, S.C.; Perkins, C.; Sholl, A.; Finkelman, F.D.; Hildeman, D.A. IL-4 and IL-15 promotion of virtual memory CD8+T cells is determined by genetic background. Eur. J. Immunol. 2016, 46, 2333–2339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Holzapfel, K.L.; Zhu, J.; Jameson, S.C.; Hogquist, K.A. Steady-state production of IL-4 modulates immunity in mouse strains and is determined by lineage diversity of iNKT cells. Nat. Immunol. 2013, 14, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Zhu, J.; Wang, T.; Hu-Li, J.; Terabe, M.; Berzofsky, J.A.; Clayberger, C.; Krensky, A.M. KLF13 sustains thymic memory-like CD8+ T cells in BALB/c mice by regulating IL-4–generating invariant natural killer T cells. J. Exp. Med. 2011, 208, 1093–1103. [Google Scholar] [CrossRef] [Green Version]
- Jameson, S.C.; Lee, Y.J.; Hogquist, K.A. Innate Memory T cells. Adv. Immunol. 2015, 126, 173–213. [Google Scholar] [CrossRef] [Green Version]
- Atherly, L.O.; Lucas, J.A.; Felices, M.; Yin, C.C.; Reiner, S.L.; Berg, L.J. The Tec Family Tyrosine Kinases Itk and Rlk Regulate the Development of Conventional CD8+ T Cells. Immunology 2006, 25, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Broussard, C.; Fleischecker, C.; Horai, R.; Chetana, M.; Venegas, A.M.; Sharp, L.L.; Hedrick, S.M.; Fowlkes, B.; Schwartzberg, P.L. Altered Development of CD8+ T Cell Lineages in Mice Deficient for the Tec Kinases Itk and Rlk. Immunity 2006, 25, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Weinreich, M.A.; Odumade, O.A.; Jameson, S.C.; Hogquist, K.A. T cells expressing the transcription factor PLZF regulate the development of memory-like CD8+ T cells. Nat. Immunol. 2010, 11, 709–716. [Google Scholar] [CrossRef] [Green Version]
- Hirose, S.; Touma, M.; Go, R.; Katsuragi, Y.; Sakuraba, Y.; Gondo, Y.; Abe, M.; Sakimura, K.; Mishima, Y.; Kominami, R. Bcl11b prevents the intrathymic development of innate CD8 T cells in a cell intrinsic manner. Int. Immunol. 2014, 27, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Kastner, P.; Chan, S.; Vogel, W.K.; Zhang, L.-J.; Topark-Ngarm, A.; Golonzhka, O.; Jost, B.; Le Gras, S.; Gross, M.K.; Leid, M. Bcl11b represses a mature T-cell gene expression program in immature CD4+CD8+ thymocytes. Eur. J. Immunol. 2010, 40, 2143–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosokawa, H.; Romero-Wolf, M.; Yui, M.A.; Ungerback, J.; Quiloan, M.L.G.; Matsumoto, M.; Nakayama, K.I.; Tanaka, T.; Rothenberg, E.V. Bcl11b sets pro-T cell fate by site-specific cofactor recruitment and by repressing Id2 and Zbtb16. Nat. Immunol. 2018, 19, 1427–1440. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.L.; Patel, R.K.; Reynaldi, A.; Grenier, J.K.; Wang, J.; Watson, N.B.; Nzingha, K.; Mon, K.J.Y.; Peng, S.A.; Grimson, A.; et al. Developmental Origin Governs CD8+ T Cell Fate Decisions during Infection. Cell 2018, 174, 117–130.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, T.; Quinn, K.M. Similar but different: Virtual memory CD8 T cells as a memory-like cell population. Immunol. Cell Biol. 2019, 97, 675–684. [Google Scholar] [CrossRef]
- Pereira, B.I.; De Maeyer, R.P.H.; Covre, L.P.; Nehar-Belaid, D.; Lanna, A.; Ward, S.; Marches, R.; Chambers, E.S.; Gomes, D.C.O.; Riddell, N.E.; et al. Sestrins induce natural killer function in senescent-like CD8+ T cells. Nat. Immunol. 2020, 21, 684–694. [Google Scholar] [CrossRef]
- Smith, T.R.; Kumar, V. Revival of CD8+ Treg–mediated suppression. Trends Immunol. 2008, 29, 337–342. [Google Scholar] [CrossRef]
- Dai, Z.; Wan, N.; Zhang, S.; Moore, Y.; Wan, F. Cutting Edge: Programmed Death-1 Defines CD8+CD122+ T Cells as Regulatory versus Memory T Cells. J. Immunol. 2010, 185, 803–807. [Google Scholar] [CrossRef] [Green Version]
- Akane, K.; Kojima, S.; Mak, T.W.; Shiku, H.; Suzuki, H. CD8+CD122+CD49dlow regulatory T cells maintain T-cell homeostasis by killing activated T cells via Fas/FasL-mediated cytotoxicity. Proc. Natl. Acad. Sci. USA 2016, 113, 2460–2465. [Google Scholar] [CrossRef] [Green Version]
- Quinn, K.M.; Zaloumis, S.G.; Cukalac, T.; Kan, W.-T.; Sng, X.Y.X.; Mirams, M.; Watson, K.A.; McCaw, J.M.; Doherty, P.C.; Thomas, P.G.; et al. Heightened self-reactivity associated with selective survival, but not expansion, of naïve virus-specific CD8+ T cells in aged mice. Proc. Natl. Acad. Sci. USA 2016, 113, 1333–1338. [Google Scholar] [CrossRef] [Green Version]
- Moran, A.E.; Holzapfel, K.L.; Xing, Y.; Cunningham, N.R.; Maltzman, J.S.; Punt, J.; Hogquist, K.A. T cell receptor signal strength in Treg and iNKT cell development demonstrated by a novel fluorescent reporter mouse. J. Exp. Med. 2011, 208, 1279–1289. [Google Scholar] [CrossRef]
- Ashouri, J.F.; Weiss, A. Endogenous Nur77 Is a Specific Indicator of Antigen Receptor Signaling in Human T and B Cells. J. Immunol. 2017, 198, 657–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzam, H.S.; Grinberg, A.; Lui, K.; Shen, H.; Shores, E.W.; Love, P.E. CD5 Expression Is Developmentally Regulated By T Cell Receptor (TCR) Signals and TCR Avidity. J. Exp. Med. 1998, 188, 2301–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.H.; Klawon, D.E.J.; Zeng, S.; Lee, V.; Socci, N.D.; Savage, P.A. Eomes identifies thymic precursors of self-specific memory-phenotype CD8+ T cells. Nat. Immunol. 2020, 21, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Rudd, B.D.; Venturi, V.; Li, G.; Samadder, P.; Ertelt, J.M.; Way, S.S.; Davenport, M.P.; Nikolich-Žugich, J. Nonrandom attrition of the naive CD8+ T-cell pool with aging governed by T-cell receptor:pMHC interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 13694–13699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, E.L.; Mullen, A.C.; Martins, G.A.; Krawczyk, C.M.; Hutchins, A.S.; Zediak, V.P.; Banica, M.; DiCioccio, C.B.; Gross, D.A.; Mao, C.-A.; et al. Control of Effector CD8+ T Cell Function by the Transcription Factor Eomesodermin. Science 2003, 302, 1041–1043. [Google Scholar] [CrossRef]
- Intlekofer, A.M.; Takemoto, N.; Wherry, E.J.; A Longworth, S.; Northrup, J.T.; Palanivel, V.R.; Mullen, A.C.; Gasink, C.R.; Kaech, S.M.; Miller, J.D.; et al. Effector and memory CD8+ T cell fate coupled by T-bet and eomesodermin. Nat. Immunol. 2005, 6, 1236–1244. [Google Scholar] [CrossRef]
- Gett, A.V.; Sallusto, F.; Lanzavecchia, A.; Geginat, J. T cell fitness determined by signal strength. Nat. Immunol. 2003, 4, 355–360. [Google Scholar] [CrossRef]
- Martinet, V.; Tonon, S.; Torres, D.; Azouz, A.; Nguyen, M.; Köhler, A.; Flamand, V.; Mao, C.-A.; Klein, W.H.; Leo, O.; et al. Type I interferons regulate eomesodermin expression and the development of unconventional memory CD8+ T cells. Nat. Commun. 2015, 6, 7089. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Wang, X.; Jameson, S.C.; A Hogquist, Y.X.X.W.S.C.J.K. Late stages of T cell maturation in the thymus involve NF-κB and tonic type I interferon signaling. Nat. Immunol. 2016, 17, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Akue, A.D.; Lee, J.-Y.; Jameson, S.C. Derivation and Maintenance of Virtual Memory CD8 T Cells. J. Immunol. 2012, 188, 2516–2523. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.-H.; Huang, H.-H.; Zhou, M.-J.; Li, J.; Hu, W.; Huang, L.; Xu, Z.; Tu, B.; Yang, G.; Shi, M.; et al. Virtual memory CD8+ T cells restrain the viral reservoir in HIV-1-infected patients with antiretroviral therapy through derepressing KIR-mediated inhibition. Cell. Mol. Immunol. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.T.; Ernst, B.; Kieper, W.C.; Leroy, E.; Sprent, J.; Surh, C.D. Interleukin (IL)-15 and IL-7 Jointly Regulate Homeostatic Proliferation of Memory Phenotype CD8+ Cells but Are Not Required for Memory Phenotype CD4+ Cells. J. Exp. Med. 2002, 195, 1523–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, R.E.; Crossley, E.; Murray, S.; Forman, J. Memory CD8+ T Cells Provide Innate Immune Protection against Listeria monocytogenes in the Absence of Cognate Antigen. J. Exp. Med. 2003, 198, 1583–1593. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Catalfamo, M.; Li, Y.; Henkart, P.A.; Weng, N.-P. IL-15 mimics T cell receptor crosslinking in the induction of cellular proliferation, gene expression, and cytotoxicity in CD8+ memory T cells. Proc. Natl. Acad. Sci. USA 2002, 99, 6192–6197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Sun, S.; Hwang, I.; Tough, D.F.; Sprent, J. Potent and Selective Stimulation of Memory-Phenotype CD8+ T Cells In Vivo by IL-15. Immunity 1998, 8, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Pangrazzi, L.; Naismith, E.; Meryk, A.; Keller, M.; Jenewein, B.; Trieb, K.; Grubeck-Loebenstein, B. Increased IL-15 Production and Accumulation of Highly Differentiated CD8+ Effector/Memory T Cells in the Bone Marrow of Persons with Cytomegalovirus. Front. Immunol. 2017, 8, 715. [Google Scholar] [CrossRef] [Green Version]
- Ershler, W.B.; Keller, E.T. Age-Associated Increased Interleukin-6 Gene Expression, Late-Life Diseases, and Frailty. Annu. Rev. Med. 2000, 51, 245–270. [Google Scholar] [CrossRef]
- Ferrucci, L.; Corsi, A.; Lauretani, F.; Bandinelli, S.; Bartali, B.; Taub, D.D.; Guralnik, J.M.; Longo, D.L. The origins of age-related proinflammatory state. Blood 2005, 105, 2294–2299. [Google Scholar] [CrossRef] [Green Version]
- Bruunsgaard, H.; Skinhoj, P.; Pedersen, A.N.; Schroll, M.; Pedersen, B.K. Ageing, tumour necrosis factor-alpha (TNF-alpha) and atherosclerosis. Clin. Exp. Immunol. 2000, 121, 255–260. [Google Scholar] [CrossRef]
- Baez, N.S.; Cerbán, F.; Savid-Frontera, C.; Hodge, D.L.; Tosello, J.; Acosta-Rodriguez, E.; Almada, L.; Gruppi, A.; Viano, M.E.; Young, H.A.; et al. Thymic expression of IL-4 and IL-15 after systemic inflammatory or infectious Th1 disease processes induce the acquisition of "innate" characteristics during CD8+ T cell development. PLOS Pathog. 2019, 15, e1007456. [Google Scholar] [CrossRef] [Green Version]
- Chu, T.; Tyznik, A.J.; Roepke, S.; Berkley, A.M.; Woodward-Davis, A.; Pattacini, L.; Bevan, M.J.; Zehn, D.; Prlic, M. Bystander-Activated Memory CD8 T Cells Control Early Pathogen Load in an Innate-like, NKG2D-Dependent Manner. Cell Rep. 2013, 3, 701–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugolini, S.; Arpin, C.; Anfossi, N.; Walzer, T.; Cambiaggi, A.; Förster, R.; Lipp, M.; Toes, R.E.M.; Melief, C.J.; Marvel, J.; et al. Involvement of inhibitory NKRs in the survival of a subset of memory-phenotype CD8+ T cells. Nat. Immunol. 2001, 2, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, N.; Pascal, V.; Vivier, E.; Ugolini, S. Biology of T memory type 1 cells. Immunol. Rev. 2001, 181, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Coles, M.C.; McMahon, C.W.; Takizawa, H.; Raulet, D.H. Memory CD8 T lymphocytes express inhibitory MHC-specific Ly49 receptors. Eur. J. Immunol. 2000, 30, 236–244. [Google Scholar] [CrossRef]
- Kambayashi, T.; Assarsson, E.; Chambers, B.J.; Ljunggren, H.-G. Expression of the DX5 antigen on CD8+ T cells is associated with activation and subsequent cell death or memory during influenza virus infection. Eur. J. Immunol. 2001, 31, 1523–1530. [Google Scholar] [CrossRef]
- Bonorino, P.; Leroy, V.; Dufeu-Duchesne, T.; Tongiani-Dashan, S.; Sturm, N.; Pernollet, M.; Vivier, E.; Zarski, J.-P.; Marche, P.N.; Jouvin-Marche, E. Features and distribution of CD8 T cells with human leukocyte antigen class I-specific receptor expression in chronic hepatitis C. Hepatology 2007, 46, 1375–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkström, N.K.; Béziat, V.; Cichocki, F.; Liu, L.L.; Levine, J.; Larsson, S.; Koup, R.A.; Anderson, S.K.; Ljunggren, H.-G.; Malmberg, K.-J. CD8 T cells express randomly selected KIRs with distinct specificities compared with NK cells. Blood 2012, 120, 3455–3465. [Google Scholar] [CrossRef] [Green Version]
- Michel, J.J.; Griffin, P.; Vallejo, A.N. Functionally Diverse NK-Like T Cells Are Effectors and Predictors of Successful Aging. Front. Immunol. 2016, 7, 530. [Google Scholar] [CrossRef] [Green Version]
- Young, N.T.; Uhrberg, M.; Phillips, J.H.; Lanier, L.L.; Parham, P. Differential Expression of Leukocyte Receptor Complex-Encoded Ig-Like Receptors Correlates with the Transition from Effector to Memory CTL. J. Immunol. 2001, 166, 3933–3941. [Google Scholar] [CrossRef] [Green Version]
- Roger, J.; Chalifour, A.; Lemieux, S.; Duplay, P. Cutting edge: Ly49A inhibits TCR/CD3-induced apoptosis and IL-2 secretion. J. Immunol. 2001, 167, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Alter, G.; Rihn, S.; Streeck, H.; Teigen, N.; Piechocka-Trocha, A.; Moss, K.; Cohen, K.; Meier, A.; Pereyra, F.; Walker, B.; et al. Ligand-Independent Exhaustion of Killer Immunoglobulin-Like Receptor-Positive CD8+ T Cells in Human Immunodeficiency Virus Type 1 Infection. J. Virol. 2008, 82, 9668–9677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prajapati, K.; Perez, C.; Rojas, L.B.P.; Burke, B.; Guevara-Patino, J.A. Functions of NKG2D in CD8+ T cells: An opportunity for immunotherapy. Cell. Mol. Immunol. 2018, 15, 470–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, H.C.; Jeng, E.K.; Rhode, P.R. The IL-15-based superagonist ALT-803 promotes the antigen-independent conversion of memory CD8+T cells into innate-like effector cells with antitumor activity. OncoImmunology 2013, 2, e26442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tietze, J.K.; Wilkins, D.E.C.; Sckisel, G.D.; Bouchlaka, M.N.; Alderson, K.L.; Weiss, J.M.; Ames, E.; Bruhn, K.W.; Craft, N.; Wiltrout, R.H.; et al. Delineation of antigen-specific and antigen-nonspecific CD8+ memory T-cell responses after cytokine-based cancer immunotherapy. Blood 2012, 119, 3073–3083. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, N.; Robbins, S.H.; Ugolini, S.; Georgel, P.; Hoebe, K.; Bouneaud, C.; Ronet, C.; Kaser, A.; DiCioccio, C.B.; Tomasello, E.; et al. Expansion and Function of CD8+T Cells Expressing Ly49 Inhibitory Receptors Specific for MHC Class I Molecules. J. Immunol. 2004, 173, 3773–3782. [Google Scholar] [CrossRef] [Green Version]
- He, B.; Xing, S.; Chen, C.; Gao, P.; Teng, L.; Shan, Q.; Gullicksrud, J.A.; Martin, M.D.; Yu, S.; Harty, J.T.; et al. CD8+ T Cells Utilize Highly Dynamic Enhancer Repertoires and Regulatory Circuitry in Response to Infections. Immunity 2016, 45, 1341–1354. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.T.; Wherry, E.J.; Goldrath, B.Y.J.J.M.C.T.A.W. Molecular regulation of effector and memory T cell differentiation. Nat. Immunol. 2014, 15, 1104–1115. [Google Scholar] [CrossRef]
- Wang, D.; Diao, H.; Getzler, A.J.; Rogal, W.; Frederick, M.A.; Milner, J.; Yu, B.; Crotty, S.; Goldrath, A.W.; Pipkin, M.E. The Transcription Factor Runx3 Establishes Chromatin Accessibility of cis-Regulatory Landscapes that Drive Memory Cytotoxic T Lymphocyte Formation. Immunity 2018, 48, 659–674.e6. [Google Scholar] [CrossRef] [Green Version]
- Istaces, N.; Splittgerber, M.; Silva, V.L.; Nguyen, M.; Thomas, S.; Le, A.; Achouri, Y.; Calonne, E.; Defrance, M.; Fuks, F.; et al. EOMES interacts with RUNX3 and BRG1 to promote innate memory cell formation through epigenetic reprogramming. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.M.; Carty, S.A.; Kim, J.S.; Zou, T.; Smith-Garvin, J.; Alonzo, E.S.; Haimm, E.; Sant’Angelo, D.B.; Koretzky, G.A.; Reiner, S.L.; et al. Requirements for eomesodermin and promyelocytic leukemia zinc finger in the development of innate-like CD8+ T cells. J. Immunol. 2011, 186, 4573–4578. [Google Scholar] [CrossRef] [Green Version]
- Albu, D.I.; Feng, D.; Bhattacharya, D.; Jenkins, N.A.; Copeland, N.G.; Liu, P.; Avram, D. BCL11B is required for positive selection and survival of double-positive thymocytes. J. Exp. Med. 2007, 204, 3003–3015. [Google Scholar] [CrossRef] [Green Version]
- Avram, D.; Califano, D. The Multifaceted Roles of Bcl11b in Thymic and Peripheral T Cells: Impact on Immune Diseases. J. Immunol. 2014, 193, 2059–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrero, M.J.; Boué, S.; Belmonte, J.C.I. Epigenetic Mechanisms that Regulate Cell Identity. Cell Stem Cell 2010, 7, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Russ, B.E.; Olshanksy, M.; Smallwood, H.S.; Li, J.; Denton, A.E.; Prier, J.E.; Stock, A.T.; Croom, H.A.; Cullen, J.G.; Nguyen, M.L.T.; et al. Distinct Epigenetic Signatures Delineate Transcriptional Programs during Virus-Specific CD8+ T Cell Differentiation. Immunity 2014, 41, 853–865. [Google Scholar] [CrossRef] [Green Version]
- Araki, Y.; Wang, Z.; Zang, C.; Wood, W.H.; E Schones, D.; Cui, K.; Roh, T.-Y.; Lhotsky, B.; Wersto, R.P.; Peng, W.; et al. Genome-wide Analysis of Histone Methylation Reveals Chromatin State-Based Regulation of Gene Transcription and Function of Memory CD8+ T Cells. Immunity 2009, 30, 912–925. [Google Scholar] [CrossRef] [Green Version]
- Henning, A.N.; Roychoudhuri, R.; Restifo, N.P. Epigenetic control of CD8+ T cell differentiation. Nat. Rev. Immunol. 2018, 18, 340–356. [Google Scholar] [CrossRef]
- Joshi, N.S.; Cui, W.; Chandele, A.; Lee, H.K.; Urso, D.R.; Hagman, J.; Gapin, L.; Kaech, S.M. Inflammation Directs Memory Precursor and Short-Lived Effector CD8+ T Cell Fates via the Graded Expression of T-bet Transcription Factor. Immunity 2007, 27, 281–295. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.P.; Effros, R.B. T CELL REPLICATIVE SENESCENCE IN HUMAN AGING. Curr. Pharm. Des. 2013, 19, 1680–1698. [Google Scholar]
- Hodes, R.J.; Hathcock, K.S.; Weng, N.-P. Telomeres in T and B cells. Nat. Rev. Immunol. 2002, 2, 699–706. [Google Scholar] [CrossRef]
- Sen, P.; Shah, P.P.; Nativio, R.; Berger, S.L. Epigenetic Mechanisms of Longevity and Aging. Cell 2016, 166, 822–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rando, T.A.; Chang, H.Y. Aging, Rejuvenation, and Epigenetic Reprogramming: Resetting the Aging Clock. Cell 2012, 148, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucar, D.; Márquez, E.J.; Chung, C.-H.; Marches, R.; Rossi, R.J.; Uyar, A.; Wu, T.-C.; George, J.; Stitzel, M.L.; Palucka, A.K.; et al. The chromatin accessibility signature of human immune aging stems from CD8+ T cells. J. Exp. Med. 2017, 214, 3123–3144. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, D.M.; Zhang, D.W.; Hu, B.; Le Saux, S.; Yanes, R.E.; Ye, Z.; Buenrostro, J.D.; Weyand, C.M.; Greenleaf, W.J.; Goronzy, J.J. Epigenomics of human CD8 T cell differentiation and aging. Sci. Immunol. 2017, 2, eaag0192. [Google Scholar] [CrossRef] [Green Version]
- Kurachi, M.; Barnitz, R.A.; Yosef, N.; Odorizzi, P.M.; Dilorio, M.A.; Lemieux, M.E.; Yates, K.; Godec, J.; Klatt, M.G.; Regev, A.; et al. The transcription factor BATF operates as an essential differentiation checkpoint in early effector CD8+ T cells. Nat. Immunol. 2014, 15, 373–383. [Google Scholar] [CrossRef]
- Kwesi-Maliepaard, E.M.; Aslam, M.A.; Alemdehy, M.F.; Brand, T.V.D.; McLean, C.; Vlaming, H.; Van Welsem, T.; Korthout, T.; Lancini, C.; Hendriks, S.; et al. The histone methyltransferase DOT1L prevents antigen-independent differentiation and safeguards epigenetic identity of CD8+T cells. Proc. Natl. Acad. Sci. USA 2020, 117, 20706–20716. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thiele, D.; La Gruta, N.L.; Nguyen, A.; Hussain, T. Hiding in Plain Sight: Virtually Unrecognizable Memory Phenotype CD8+ T cells. Int. J. Mol. Sci. 2020, 21, 8626. https://doi.org/10.3390/ijms21228626
Thiele D, La Gruta NL, Nguyen A, Hussain T. Hiding in Plain Sight: Virtually Unrecognizable Memory Phenotype CD8+ T cells. International Journal of Molecular Sciences. 2020; 21(22):8626. https://doi.org/10.3390/ijms21228626
Chicago/Turabian StyleThiele, Daniel, Nicole L. La Gruta, Angela Nguyen, and Tabinda Hussain. 2020. "Hiding in Plain Sight: Virtually Unrecognizable Memory Phenotype CD8+ T cells" International Journal of Molecular Sciences 21, no. 22: 8626. https://doi.org/10.3390/ijms21228626
APA StyleThiele, D., La Gruta, N. L., Nguyen, A., & Hussain, T. (2020). Hiding in Plain Sight: Virtually Unrecognizable Memory Phenotype CD8+ T cells. International Journal of Molecular Sciences, 21(22), 8626. https://doi.org/10.3390/ijms21228626