TRPM8: A Therapeutic Target for Neuroinflammatory Symptoms Induced by Severe Dry Eye Disease

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
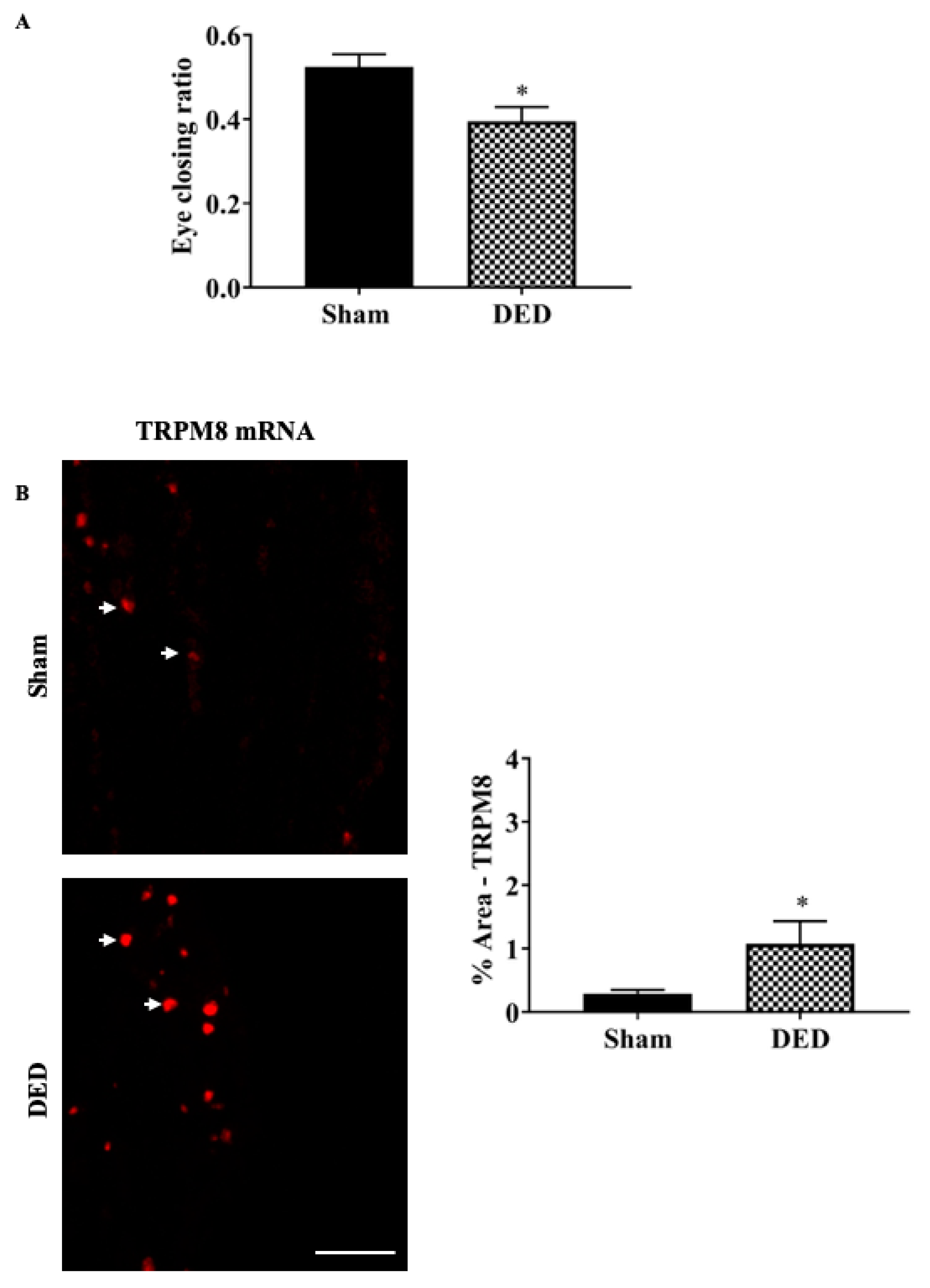
2.1. Severe DED Induced Corneal Cold Hypersensitivity Correlated with an Increase in TRPM8 mRNA Expression in the TG
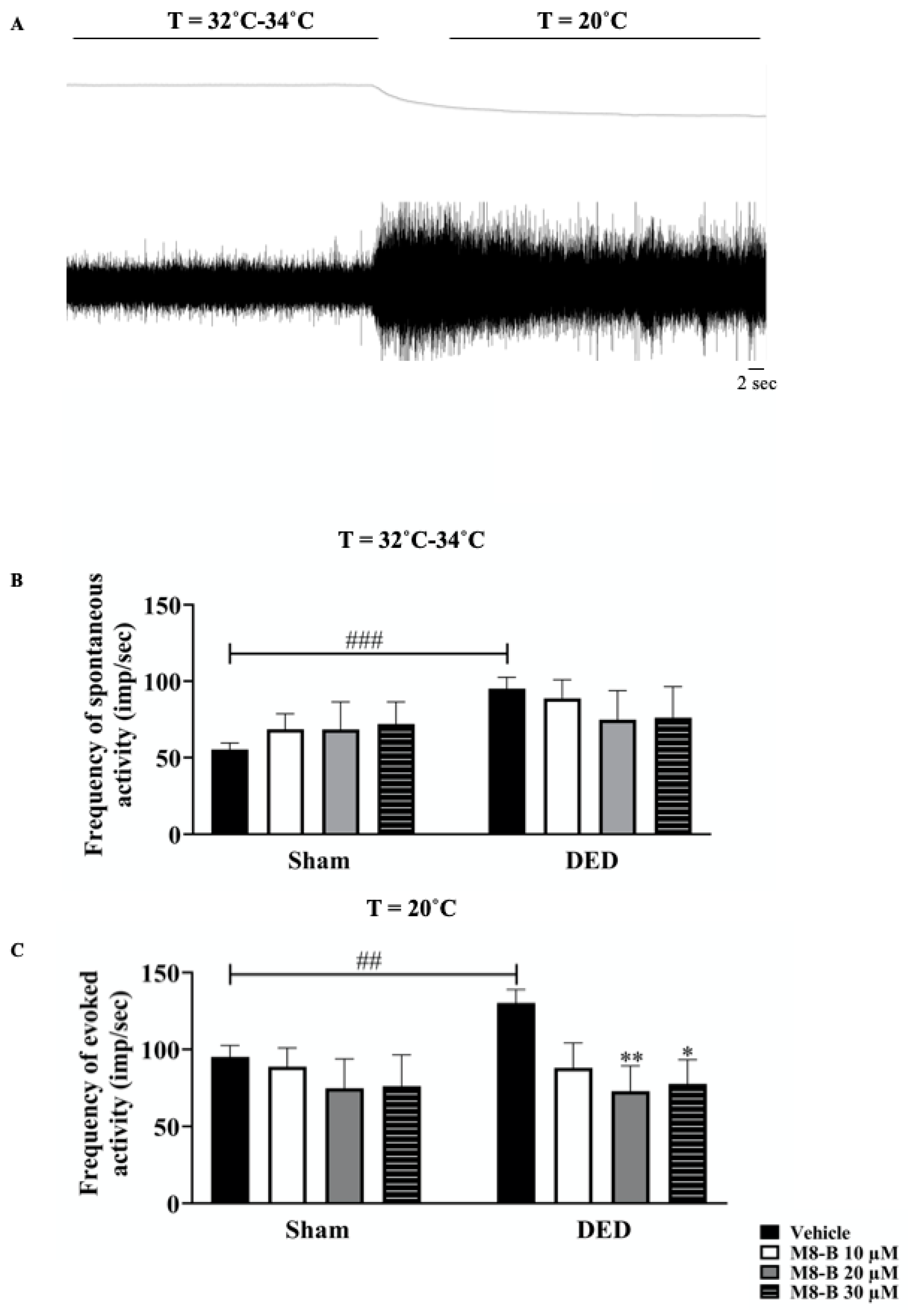
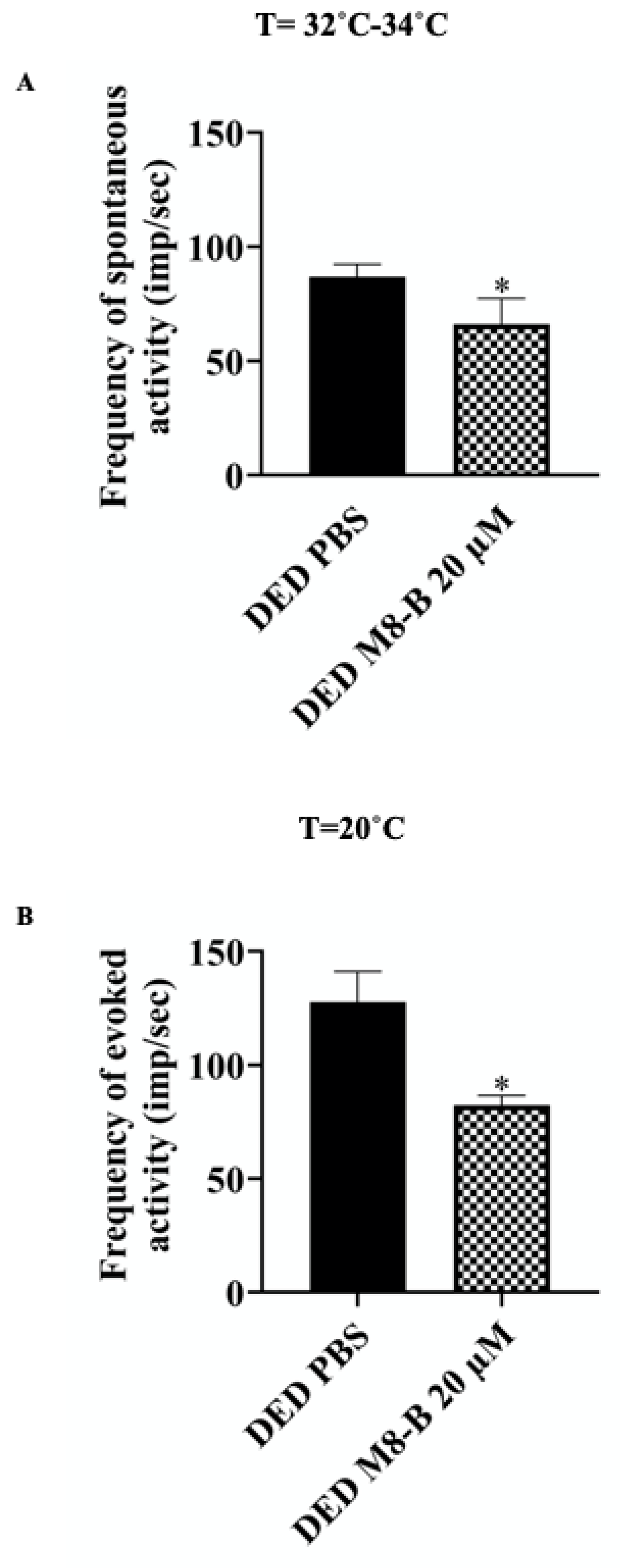
2.2. Instillation of M8-B Reduced Ongoing and Cold-Induced Electrical Activities of Thermoreceptors in Ex Vivo Eye Preparations from DED Animals
2.3. Chronic Topical Treatment with M8-B Reduced Corneal Cold Nociceptor Sensitization in DED Animals
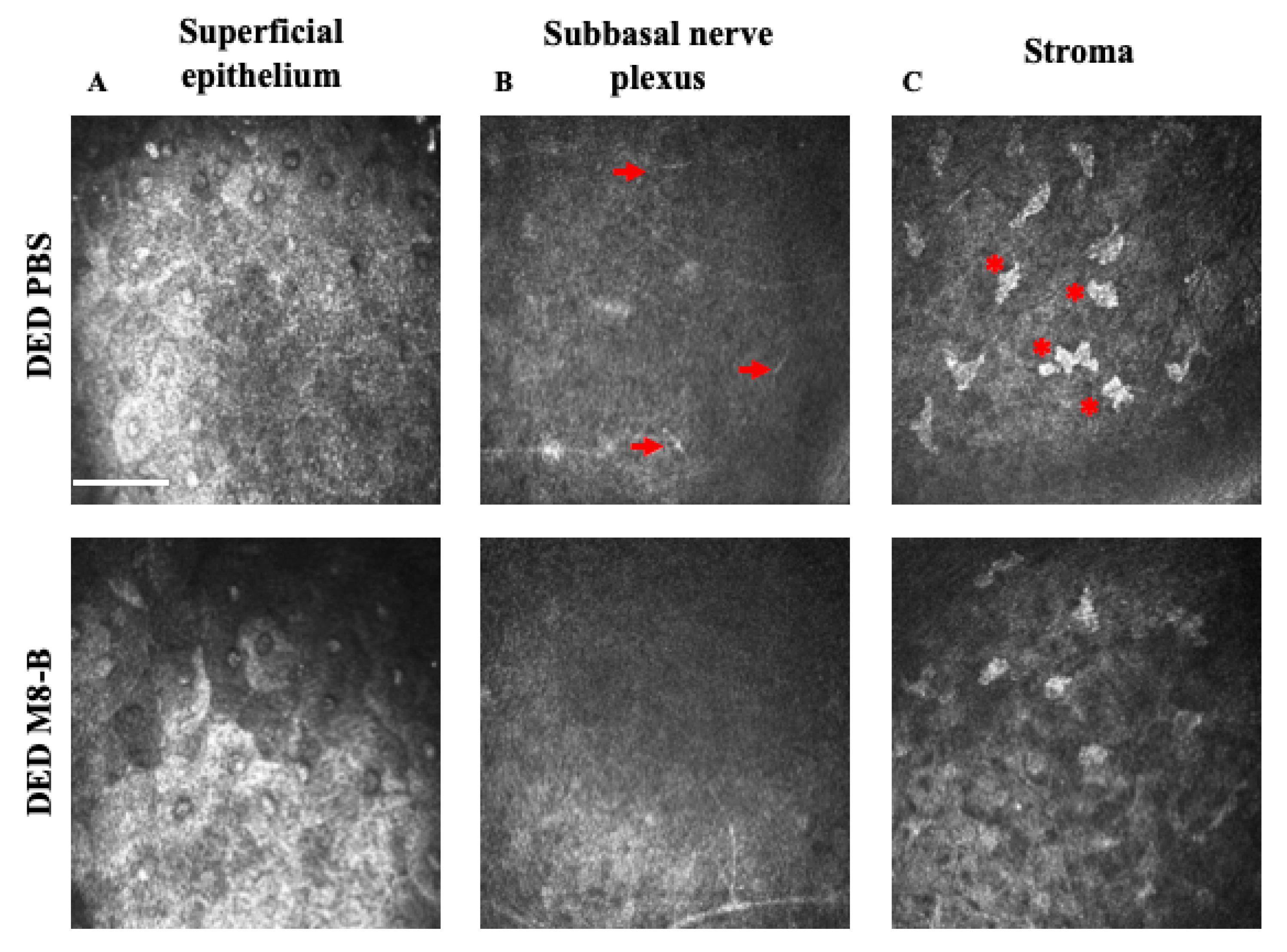
2.4. Chronic Topical Treatment with M8-B Reduced the Corneal Inflammation in the DED-Animals
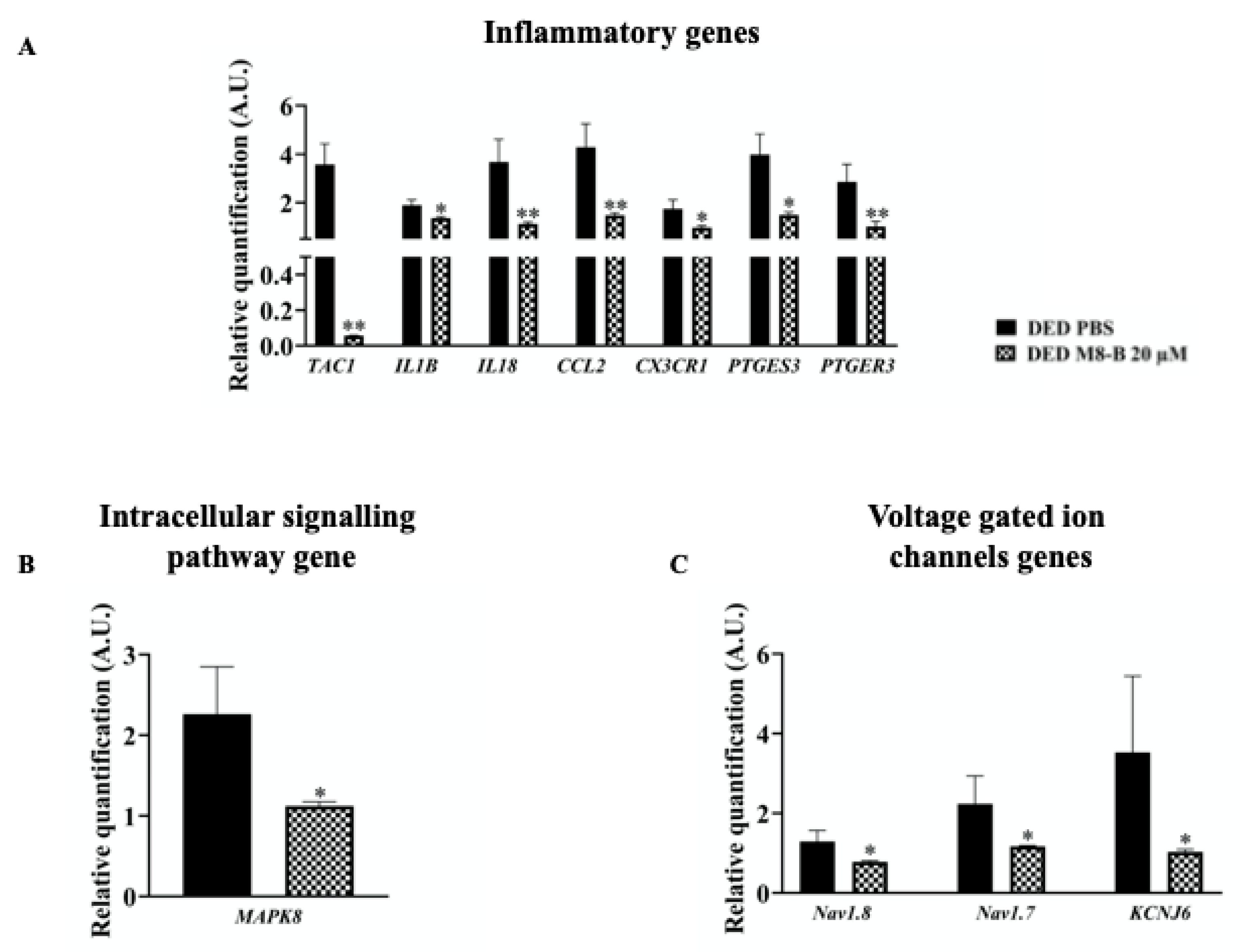
2.5. Chronic Topical Treatment with M8-B Reduced Inflammatory Gene Expressions in the TG of DED Animals
2.6. Chronic Topical Treatment with M8-B Decreased the Expression of Genes Implicated in Pain Conduction
2.6.1. Intracellular Signal Transduction
2.6.2. Voltage-Gated Ion Channels
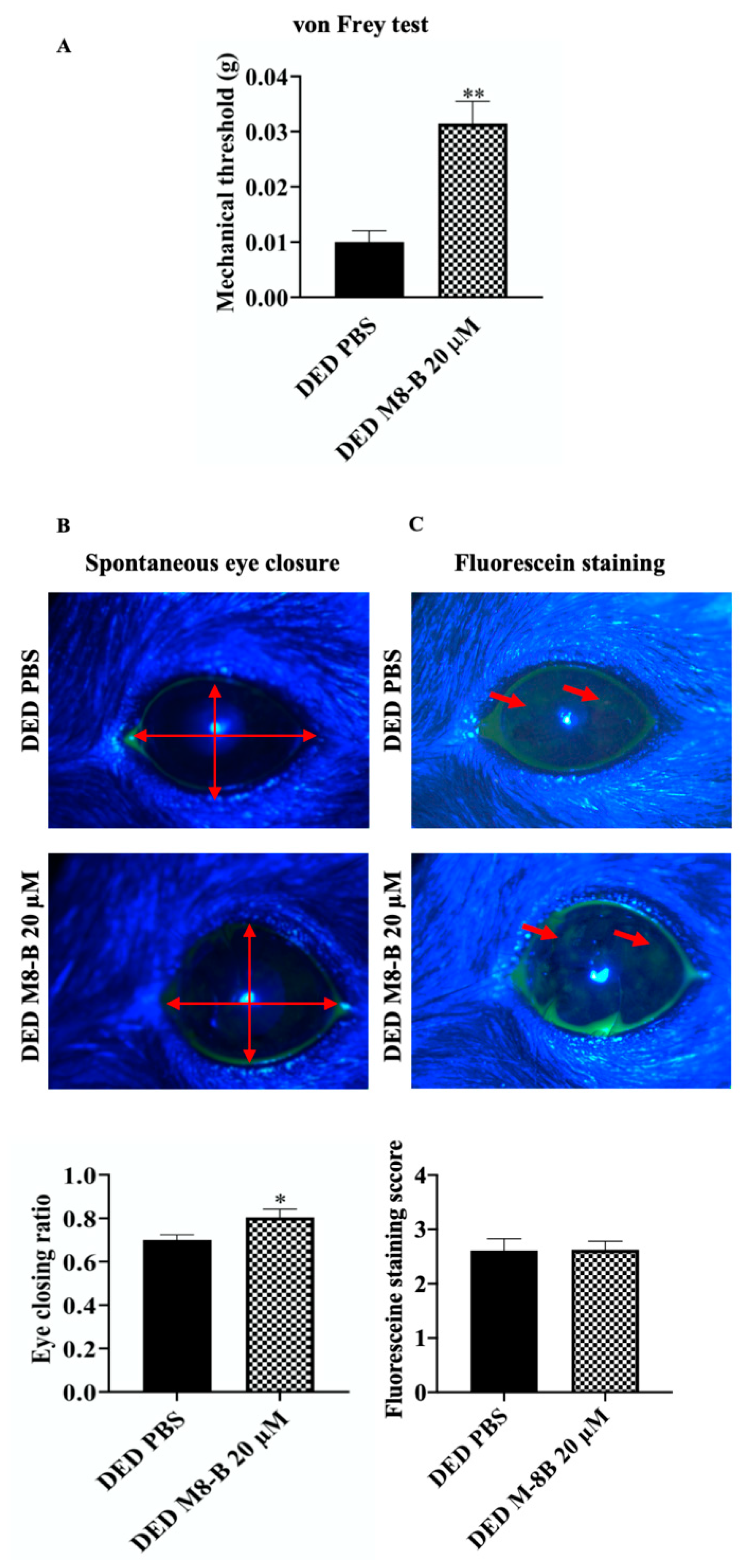
2.7. Pharmacological Blockage of TRPM8 Reduced Spontaneous and Mechanical-Evoked Ocular Pain Sensations in DED Mice
3. Discussion
4. Methods
4.1. Experimental Animals
4.2. Surgical Procedures
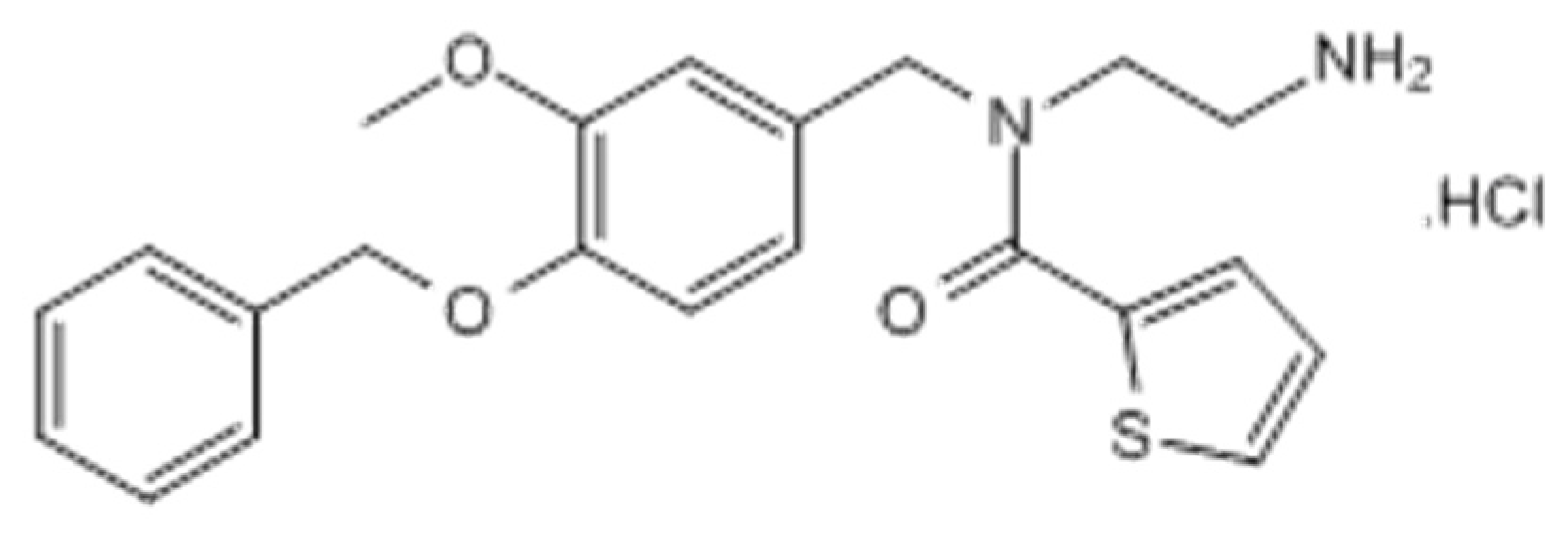
4.3. Drugs
4.4. Experimental Study
4.5. Behavioral Tests
4.6. Measurement of Corneal Sensitivity to Mechanical and Cold Stimulation
4.7. Spontaneous Pain: Eye Closing Ratio Measurement
4.8. Fluorescein Staining Score
4.9. In Vivo Confocal Microscopy
4.10. Multi-Unit Extracellular Recording of Evoked Ciliary Nerve Fiber Activity in Ex Vivo Eye Preparations
4.11. RT-qPCR Analysis
4.11.1. Tissue Preparation for RT-qPCR Analysis
4.11.2. RT-qPCR Analysis
4.12. Tissue Preparation for Fluorescent In Situ Hybridization
4.13. Fluorescent RNAscope In Situ Hybridization
4.14. Microscopic and Nanozoomer Analysis
4.15. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DED | Dry eye disease |
| TRPM8 | Transient receptor potential melastatin 8 |
| TG | Trigeminal ganglion |
| Min | Minutes |
| SEM | Standard error of the mean |
| Imp | Impulse |
| Sec | Seconds |
| TAC1 | Tachykinin Precursor 1 |
| IL-1β | Interleukin-1β |
| IL18 | Interleukin 18 |
| CCL2 | Chemokine (C-C motif) ligand 2 |
| CX3CR1 | CX3C chemokine receptor 1 |
| PTGES3 | Prostaglandin E synthase 3 |
| PTGER3 | Prostaglandin E receptor 3 |
| MAPK | Mitogen-activated protein kinase |
| Nav1.7 | Sodium channel 1.7 |
| Nav1.8 | Sodium channel 1.8 |
| KCNJ6 | Potassium channel 2 |
| g | Gram |
| ELG | Extraorbital lacrimal gland |
| HG | Harderian gland |
| HB-LTCs | high-background low-threshold cold thermoreceptors |
| LB-HTCs | Low-background high-threshold cold thermoreceptors |
| TRPV1 | The transient receptor potential vanilloid 1 |
| h | Hour |
| i.p. | Intraperitoneal |
| RT | Room temperature |
References
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.-S.; Schaumberg, D.; Uchino, M.; Vehof, J.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, C.; Nichols, J.J.; Cox, S.M.; Brock, J.A.; Begley, C.G.; Bereiter, D.A.; Dartt, D.A.; Galor, A.; Hamrah, P.; Ivanusic, J.J.; et al. TFOS DEWS II pain and sensation report. Ocul. Surf. 2017, 15, 404–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, S.; Hamrah, P. Understanding Neuropathic Corneal Pain-Gaps and Current Therapeutic Approaches. Semin. Ophthalmol. 2016, 31, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Cruzat, A.; Qazi, Y.; Hamrah, P. In Vivo Confocal Microscopy of Corneal Nerves in Health and Disease. Ocul. Surf. 2017, 15, 15–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marfurt, C.F.; Kingsley, R.E.; Echtenkamp, S.E. Sensory and sympathetic innervation of the mammalian cornea. A retrograde tracing study. Investig. Ophthalmol. Vis. Sci. 1989, 30, 461–472. [Google Scholar]
- Marfurt, C.F.; Del Toro, D.R. Corneal sensory pathway in the rat: A horseradish peroxidase tracing study. J. Comp. Neurol. 1987, 261, 450–459. [Google Scholar] [CrossRef]
- Belmonte, C.; Acosta, M.C.; Merayo-Lloves, J.; Gallar, J. What Causes Eye Pain? Curr. Ophthalmol. Rep. 2015, 3, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Meng, I.D. Cold-sensitive corneal afferents respond to a variety of ocular stimuli central to tear production: Implications for dry eye disease. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3969–3976. [Google Scholar] [CrossRef]
- Parra, A.; Madrid, R.; Echevarria, D.; del Olmo, S.; Morenilla-Palao, C.; Acosta, M.C.; Gallar, J.; Dhaka, A.; Viana, F.; Belmonte, C. Ocular surface wetness is regulated by TRPM8-dependent cold thermoreceptors of the cornea. Nat. Med. 2010, 16, 1396–1399. [Google Scholar] [CrossRef]
- Quallo, T.; Vastani, N.; Horridge, E.; Gentry, C.; Parra, A.; Moss, S.; Viana, F.; Belmonte, C.; Andersson, D.A.; Bevan, S. TRPM8 is a neuronal osmosensor that regulates eye blinking in mice. Nat. Commun. 2015, 6, 7150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmonte, C.; Gallar, J. Cold thermoreceptors, unexpected players in tear production and ocular dryness sensations. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3888–3892. [Google Scholar] [CrossRef] [Green Version]
- González-Muñiz, R.; Bonache, M.A.; Martín-Escura, C.; Gómez-Monterrey, I. Recent Progress in TRPM8 Modulation: An Update. Int. J. Mol. Sci. 2019, 20, 2618. [Google Scholar] [CrossRef] [Green Version]
- Alcalde, I.; Íñigo-Portugués, A.; González-González, O.; Almaraz, L.; Artime, E.; Morenilla-Palao, C.; Gallar, J.; Viana, F.; Merayo-Lloves, J.; Belmonte, C. Morphological and functional changes in TRPM8-expressing corneal cold thermoreceptor neurons during aging and their impact on tearing in mice. J. Comp. Neurol. 2018, 526, 1859–1874. [Google Scholar] [CrossRef]
- Piña, R.; Ugarte, G.; Campos, M.; Íñigo-Portugués, A.; Olivares, E.; Orio, P.; Belmonte, C.; Bacigalupo, J.; Madrid, R. Role of TRPM8 Channels in Altered Cold Sensitivity of Corneal Primary Sensory Neurons Induced by Axonal Damage. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 8177–8192. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Li, S.; Dhaka, A.; Story, G.M.; Cao, Y.-Q. Expression of the transient receptor potential channels TRPV1, TRPA1 and TRPM8 in mouse trigeminal primary afferent neurons innervating the dura. Mol. Pain 2012, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Colburn, R.W.; Lubin, M.L.; Stone Jr, D.J.; Wang, Y.; Lawrence, D.; D’Andrea, M.R.; Brandt, M.R.; Liu, Y.; Flores, C.M.; Qin, N. Attenuated cold sensitivity in TRPM8 null mice. Neuron 2007, 54, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Dhaka, A.; Murray, A.N.; Mathur, J.; Earley, T.J.; Petrus, M.J.; Patapoutian, A. TRPM8 is required for cold sensation in mice. Neuron 2007, 54, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Bautista, D.M.; Siemens, J.; Glazer, J.M.; Tsuruda, P.R.; Basbaum, A.I.; Stucky, C.L.; Jordt, S.-E.; Julius, D. The menthol receptor TRPM8 is the principal detector of environmental cold. Nature 2007, 448, 204–208. [Google Scholar] [CrossRef]
- Zimmermann, M. Pathobiology of neuropathic pain. Eur. J. Pharmacol. 2001, 429, 23–37. [Google Scholar] [CrossRef]
- Jensen, T.S.; Gottrup, H.; Sindrup, S.H.; Bach, F.W. The clinical picture of neuropathic pain. Eur. J. Pharmacol. 2001, 429, 1–11. [Google Scholar] [CrossRef]
- Marwaha, L.; Bansal, Y.; Singh, R.; Saroj, P.; Bhandari, R.; Kuhad, A. TRP channels: Potential drug target for neuropathic pain. Inflammopharmacology 2016, 24, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Caspani, O.; Zurborg, S.; Labuz, D.; Heppenstall, P.A. The Contribution of TRPM8 and TRPA1 Channels to Cold Allodynia and Neuropathic Pain. PLOS ONE 2009, 4, e7383. [Google Scholar] [CrossRef] [Green Version]
- Frederick, J.; Buck, M.E.; Matson, D.J.; Cortright, D.N. Increased TRPA1, TRPM8, and TRPV2 Expression in Dorsal Root Ganglia by Nerve Injury. Available online: http://pubmed.ncbi.nlm.nih.gov/17517374/ (accessed on 31 August 2020).
- Xing, H.; Chen, M.; Ling, J.; Tan, W.; Gu, J.G. TRPM8 mechanism of cold allodynia after chronic nerve injury. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 13680–13690. [Google Scholar] [CrossRef] [Green Version]
- Koh, W.U.; Choi, S.-S.; Kim, J.H.; Yoon, H.J.; Ahn, H.-S.; Lee, S.K.; Leem, J.G.; Song, J.G.; Shin, J.W. The preventive effect of resiniferatoxin on the development of cold hypersensitivity induced by spinal nerve ligation: Involvement of TRPM8. BMC Neurosci. 2016, 17, 38. [Google Scholar] [CrossRef] [Green Version]
- Knowlton, W.M.; Daniels, R.L.; Palkar, R.; McCoy, D.D.; McKemy, D.D. Pharmacological Blockade of TRPM8 Ion Channels Alters Cold and Cold Pain Responses in Mice. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.J.; Parsons, W.H.; Colburn, R.W.; Meegalla, S.K.; Ballentine, S.K.; Illig, C.R.; Qin, N.; Liu, Y.; Hutchinson, T.L.; Lubin, M.L.; et al. Design and optimization of benzimidazole-containing transient receptor potential melastatin 8 (TRPM8) antagonists. J. Med. Chem. 2011, 54, 233–247. [Google Scholar] [CrossRef]
- Sałat, K.; Filipek, B. Antinociceptive activity of transient receptor potential channel TRPV1, TRPA1, and TRPM8 antagonists in neurogenic and neuropathic pain models in mice. J. Zhejiang Univ. Sci. B 2015, 16, 167–178. [Google Scholar] [CrossRef]
- De Caro, C.; Russo, R.; Avagliano, C.; Cristiano, C.; Calignano, A.; Aramini, A.; Bianchini, G.; Allegretti, M.; Brandolini, L. Antinociceptive effect of two novel transient receptor potential melastatin 8 antagonists in acute and chronic pain models in rat. Br. J. Pharmacol. 2018, 175, 1691–1706. [Google Scholar] [CrossRef] [Green Version]
- De Caro, C.; Cristiano, C.; Avagliano, C.; Bertamino, A.; Ostacolo, C.; Campiglia, P.; Gomez-Monterrey, I.; La Rana, G.; Gualillo, O.; Calignano, A.; et al. Characterization of New TRPM8 Modulators in Pain Perception. Int. J. Mol. Sci. 2019, 20, 5544. [Google Scholar] [CrossRef] [Green Version]
- Kovács, I.; Luna, C.; Quirce, S.; Mizerska, K.; Callejo, G.; Riestra, A.; Fernández-Sánchez, L.; Meseguer, V.M.; Cuenca, N.; Merayo-Lloves, J.; et al. Abnormal activity of corneal cold thermoreceptors underlies the unpleasant sensations in dry eye disease. Pain 2016, 157, 399–417. [Google Scholar] [CrossRef] [PubMed]
- Kurose, M.; Meng, I.D. Dry eye modifies the thermal and menthol responses in rat corneal primary afferent cool cells. J. Neurophysiol. 2013, 110, 495–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, H.; Oshinsky, M.L. Ocular dryness excites two classes of corneal afferent neurons implicated in basal tearing in rats: Involvement of transient receptor potential channels. J. Neurophysiol. 2012, 107, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Robbins, A.; Kurose, M.; Winterson, B.J.; Meng, I.D. Menthol activation of corneal cool cells induces TRPM8-mediated lacrimation but not nociceptive responses in rodents. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7034–7042. [Google Scholar] [CrossRef] [PubMed]
- Weyer, A.D.; Lehto, S.G. Development of TRPM8 Antagonists to Treat Chronic Pain and Migraine. Pharmaceuticals 2017, 10, 37. [Google Scholar] [CrossRef]
- Yang, J.M.; Li, F.; Liu, Q.; Rüedi, M.; Wei, E.T.; Lentsman, M.; Lee, H.S.; Choi, W.; Kim, S.J.; Yoon, K.C. A novel TRPM8 agonist relieves dry eye discomfort. BMC Ophthalmol. 2017, 17, 101. [Google Scholar] [CrossRef] [Green Version]
- Wasner, G.; Schattschneider, J.; Binder, A.; Baron, R. Topical menthol—A human model for cold pain by activation and sensitization of C nociceptors. Brain 2004, 127, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Hatem, S.; Attal, N.; Willer, J.-C.; Bouhassira, D. Psychophysical study of the effects of topical application of menthol in healthy volunteers. Pain 2006, 122, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Acosta, M.C.; Belmonte, C.; Gallar, J. Sensory experiences in humans and single-unit activity in cats evoked by polymodal stimulation of the cornea. J. Physiol. 2001, 534, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Fakih, D.; Zhao, Z.; Nicolle, P.; Reboussin, E.; Joubert, F.; Luzu, J.; Labbé, A.; Rostène, W.; Baudouin, C.; Mélik Parsadaniantz, S.; et al. Chronic dry eye induced corneal hypersensitivity, neuroinflammatory responses, and synaptic plasticity in the mouse trigeminal brainstem. J. Neuroinflamm. 2019, 16, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Situ, P.; Simpson, T.L.; Fonn, D.; Jones, L.W. Conjunctival and corneal pneumatic sensitivity is associated with signs and symptoms of ocular dryness. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2971–2976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowlton, W.M.; Palkar, R.; Lippoldt, E.K.; McCoy, D.D.; Baluch, F.; Chen, J.; McKemy, D.D. A sensory-labeled line for cold: TRPM8-expressing sensory neurons define the cellular basis for cold, cold pain, and cooling-mediated analgesia. J. Neurosci. 2013, 33, 2837–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namer, B.; Seifert, F.; Handwerker, H.O.; Maihöfner, C. TRPA1 and TRPM8 activation in humans: Effects of cinnamaldehyde and menthol. Neuroreport 2005, 16, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, C.J.; Garry, E.M.; Cottrell, D.F.; Rosie, R.; Anderson, H.; Robertson, D.C.; Fleetwood-Walker, S.M.; Mitchell, R. Analgesia mediated by the TRPM8 cold receptor in chronic neuropathic pain. Curr. Biol. 2006, 16, 1591–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier, V.; Roudaut, Y.; Osorio, N.; Aimonetti, J.-M.; Ribot-Ciscar, E.; Penalba, V.; Merrot, T.; Lebonvallet, N.; Le Gall-Ianotto, C.; Misery, L.; et al. Merkel Cells Sense Cooling with TRPM8 Channels. J. Investig. Dermatol. 2018, 138, 946–956. [Google Scholar] [CrossRef] [Green Version]
- Urata, T.; Mori, N.; Fukuwatari, T. Vagus nerve is involved in the changes in body temperature induced by intragastric administration of 1,8-cineole via TRPM8 in mice. Neurosci. Lett. 2017, 650, 65–71. [Google Scholar] [CrossRef]
- Almeida, M.C.; Hew-Butler, T.; Soriano, R.N.; Rao, S.; Wang, W.; Wang, J.; Tamayo, N.; Oliveira, D.L.; Nucci, T.B.; Aryal, P.; et al. Pharmacological Blockade of the Cold Receptor TRPM8 Attenuates Autonomic and Behavioral Cold Defenses and Decreases Deep Body Temperature. J. Neurosci. 2012, 32, 2086–2099. [Google Scholar] [CrossRef] [Green Version]
- Hatta, A.; Kurose, M.; Sullivan, C.; Okamoto, K.; Fujii, N.; Yamamura, K.; Meng, I.D. Dry eye sensitizes cool cells to capsaicin-induced changes in activity via TRPV1. J. Neurophysiol. 2019, 121, 2191–2201. [Google Scholar] [CrossRef]
- Bereiter, D.A.; Rahman, M.; Thompson, R.; Stephenson, P.; Saito, H. TRPV1 and TRPM8 Channels and Nocifensive Behavior in a Rat Model for Dry Eye. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3739–3746. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Okamoto, K.; Thompson, R.; Katagiri, A.; Bereiter, D.A. Sensitization of trigeminal brainstem pathways in a model for tear deficient dry eye. Pain 2015, 156, 942–950. [Google Scholar] [CrossRef]
- Belmonte, C.; Aracil, A.; Acosta, M.C.; Luna, C.; Gallar, J. Nerves and sensations from the eye surface. Ocul Surf. 2004, 2, 248–253. [Google Scholar] [CrossRef]
- González-González, O.; Bech, F.; Gallar, J.; Merayo-Lloves, J.; Belmonte, C. Functional Properties of Sensory Nerve Terminals of the Mouse Cornea. Investig. Ophthalmol. Vis. Sci. 2017, 58, 404–415. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.A.; Pianova, S.; Belmonte, C. Differences between nerve terminal impulses of polymodal nociceptors and cold sensory receptors of the guinea-pig cornea. J. Physiol. 2001, 533, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Bech, F.; González-González, O.; Artime, E.; Serrano, J.; Alcalde, I.; Gallar, J.; Merayo-Lloves, J.; Belmonte, C. Functional and Morphologic Alterations in Mechanical, Polymodal, and Cold Sensory Nerve Fibers of the Cornea Following Photorefractive Keratectomy. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2281–2292. [Google Scholar] [CrossRef]
- Baudouin, C.; Messmer, E.M.; Aragona, P.; Geerling, G.; Akova, Y.A.; Benítez-del-Castillo, J.; Boboridis, K.G.; Merayo-Lloves, J.; Rolando, M.; Labetoulle, M. Revisiting the vicious circle of dry eye disease: A focus on the pathophysiology of meibomian gland dysfunction. Br. J. Ophthalmol. 2016, 100, 300–306. [Google Scholar] [CrossRef]
- Liu, H.; Hua, L.; Liu, Q.; Pan, J.; Bao, Y. Cold Stimuli Facilitate Inflammatory Responses Through Transient Receptor Potential Melastatin 8 (TRPM8) in Primary Airway Epithelial Cells of Asthmatic Mice. Inflammation 2018, 41, 1266–1275. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Q.; Hua, L.; Pan, J. Inhibition of transient receptor potential melastatin 8 alleviates airway inflammation and remodeling in a murine model of asthma with cold air stimulus. Acta Biochim. Biophys. Sin. 2018, 50, 499–506. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.-F.; Zhang, Y.; Rittenhouse, K.D.; Pickering, E.H.; McDowell, M.T. Evaluations of Tear Protein Markers in Dry Eye Disease: Repeatability of Measurement and Correlation with Disease. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4556–4564. [Google Scholar] [CrossRef] [Green Version]
- Willcox, M.D.P.; Argüeso, P.; Georgiev, G.A.; Holopainen, J.M.; Laurie, G.W.; Millar, T.J.; Papas, E.B.; Rolland, J.P.; Schmidt, T.A.; Stahl, U.; et al. TFOS DEWS II Tear Film Report. Ocul. Surf. 2017, 15, 366–403. [Google Scholar] [CrossRef] [Green Version]
- Nicolle, P.; Liang, H.; Reboussin, E.; Rabut, G.; Warcoin, E.; Brignole-Baudouin, F.; Melik-Parsadaniantz, S.; Baudouin, C.; Labbe, A.; Reaux-Le Goazigo, A. Proinflammatory Markers, Chemokines, and Enkephalin in Patients Suffering from Dry Eye Disease. Int. J. Mol. Sci. 2018, 19, 1221. [Google Scholar] [CrossRef] [Green Version]
- Noël, J.; Zimmermann, K.; Busserolles, J.; Deval, E.; Alloui, A.; Diochot, S.; Guy, N.; Borsotto, M.; Reeh, P.; Eschalier, A.; et al. The mechano-activated K+ channels TRAAK and TREK-1 control both warm and cold perception. EMBO J. 2009, 28, 1308–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrid, R.; de la Peña, E.; Donovan-Rodriguez, T.; Belmonte, C.; Viana, F. Variable threshold of trigeminal cold-thermosensitive neurons is determined by a balance between TRPM8 and Kv1 potassium channels. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 3120–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahamsen, B.; Zhao, J.; Asante, C.O.; Cendan, C.M.; Marsh, S.; Martinez-Barbera, J.P.; Nassar, M.A.; Dickenson, A.H.; Wood, J.N. The cell and molecular basis of mechanical, cold, and inflammatory pain. Science 2008, 321, 702–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Yang, W.; Jiang, H.; Guo, C.; Huang, A.J.W.; Hu, H.; Liu, Q. TRPV1 activity and substance P release are required for corneal cold nociception. Nat. Commun. 2019, 10, 5678. [Google Scholar] [CrossRef]
- Cao, Y.Q.; Mantyh, P.W.; Carlson, E.J.; Gillespie, A.M.; Epstein, C.J.; Basbaum, A.I. Primary afferent tachykinins are required to experience moderate to intense pain. Nature 1998, 392, 390–394. [Google Scholar] [CrossRef]
- Amann, R.; Schuligoi, R.; Holzer, P.; Donnerer, J. The non-peptide NK1 receptor antagonist SR140333 produces long-lasting inhibition of neurogenic inflammation, but does not influence acute chemo- or thermonociception in rats. Naunyn Schmiedebergs Arch. Pharmacol. 1995, 352, 201–205. [Google Scholar] [CrossRef]
- Klein, A.H.; Sawyer, C.M.; Carstens, M.I.; Tsagareli, M.G.; Tsiklauri, N.; Carstens, E. Topical application of L-menthol induces heat analgesia, mechanical allodynia, and a biphasic effect on cold sensitivity in rats. Behav. Brain Res. 2010, 212, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Langford, D.J.; Bailey, A.L.; Chanda, M.L.; Clarke, S.E.; Drummond, T.E.; Echols, S.; Glick, S.; Ingrao, J.; Klassen-Ross, T.; Lacroix-Fralish, M.L.; et al. Coding of facial expressions of pain in the laboratory mouse. Nat. Methods 2010, 7, 447–449. [Google Scholar] [CrossRef]
- Matsumiya, L.C.; Sorge, R.E.; Sotocinal, S.G.; Tabaka, J.M.; Wieskopf, J.S.; Zaloum, A.; King, O.D.; Mogil, J.S. Using the Mouse Grimace Scale to reevaluate the efficacy of postoperative analgesics in laboratory mice. J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 42–49. [Google Scholar]
- Pauly, A.; Brignole-Baudouin, F.; Labbé, A.; Liang, H.; Warnet, J.-M.; Baudouin, C. New Tools for the Evaluation of Toxic Ocular Surface Changes in the Rat. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5473–5483. [Google Scholar] [CrossRef] [Green Version]
- Joubert, F.; Acosta, M.D.C.; Gallar, J.; Fakih, D.; Sahel, J.-A.; Baudouin, C.; Bodineau, L.; Mélik Parsadaniantz, S.; Réaux-Le Goazigo, A. Effects of corneal injury on ciliary nerve fibre activity and corneal nociception in mice: A behavioural and electrophysiological study. Eur. J. Pain 2019, 23, 589–602. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fakih, D.; Baudouin, C.; Réaux-Le Goazigo, A.; Mélik Parsadaniantz, S. TRPM8: A Therapeutic Target for Neuroinflammatory Symptoms Induced by Severe Dry Eye Disease. Int. J. Mol. Sci. 2020, 21, 8756. https://doi.org/10.3390/ijms21228756
Fakih D, Baudouin C, Réaux-Le Goazigo A, Mélik Parsadaniantz S. TRPM8: A Therapeutic Target for Neuroinflammatory Symptoms Induced by Severe Dry Eye Disease. International Journal of Molecular Sciences. 2020; 21(22):8756. https://doi.org/10.3390/ijms21228756
Chicago/Turabian StyleFakih, Darine, Christophe Baudouin, Annabelle Réaux-Le Goazigo, and Stéphane Mélik Parsadaniantz. 2020. "TRPM8: A Therapeutic Target for Neuroinflammatory Symptoms Induced by Severe Dry Eye Disease" International Journal of Molecular Sciences 21, no. 22: 8756. https://doi.org/10.3390/ijms21228756
APA StyleFakih, D., Baudouin, C., Réaux-Le Goazigo, A., & Mélik Parsadaniantz, S. (2020). TRPM8: A Therapeutic Target for Neuroinflammatory Symptoms Induced by Severe Dry Eye Disease. International Journal of Molecular Sciences, 21(22), 8756. https://doi.org/10.3390/ijms21228756