Meibomian Gland Dysfunction: What Have Animal Models Taught Us?




Abstract
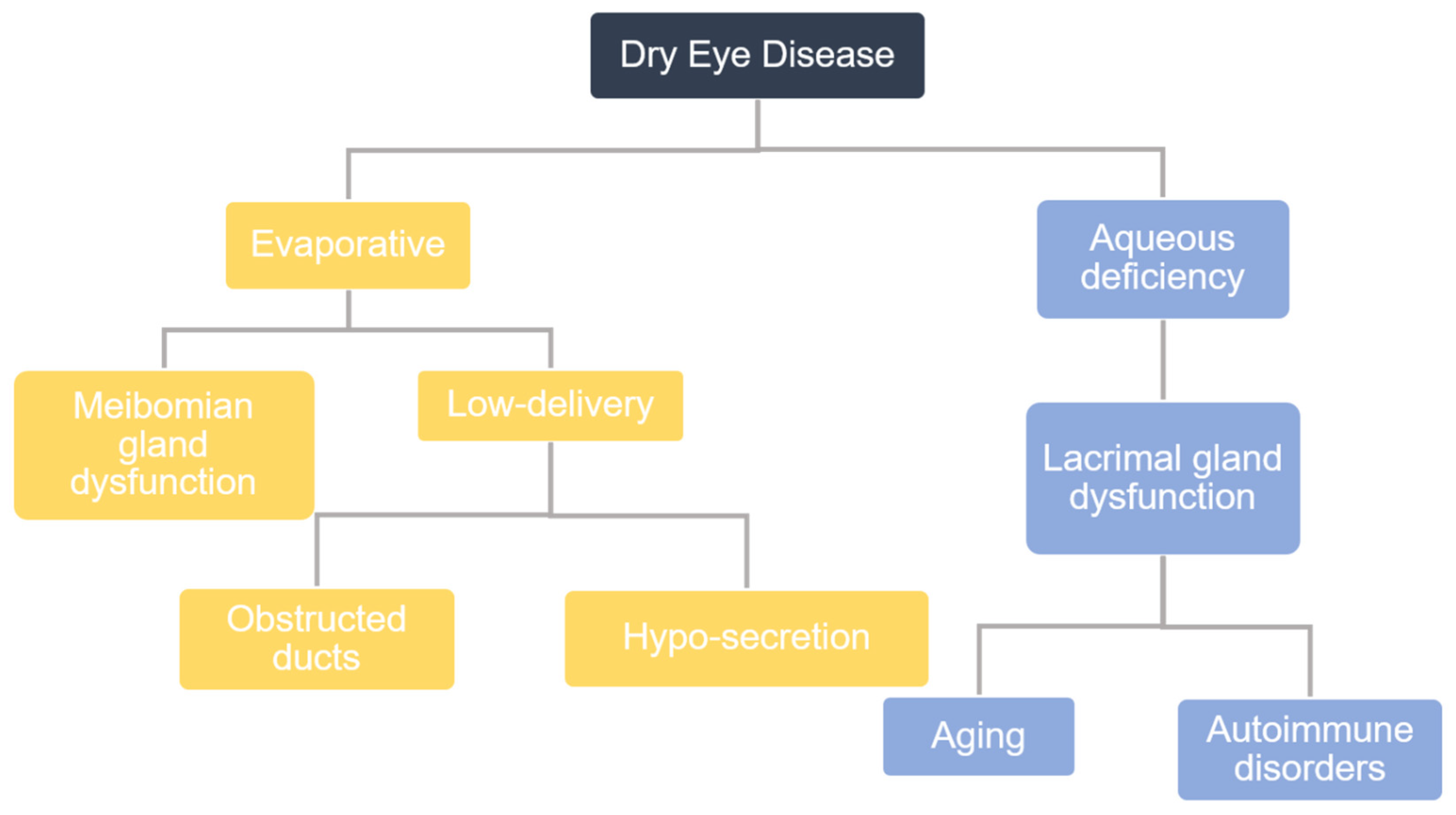
:1. Introduction
2. Meibomian Gland Structure and Function
3. Meibomian Gland Morphogenesis
4. Meibomian Gland Dysfunction and Pathology
5. Pathogenesis of MGD in Humans and Other Mammalian Species
6. Interspecies Variations in Meibum Composition
7. Mouse Models Used to Study DED and ARMGD
7.1. Aging Mice as a Model for ARMGD
7.2. Correlation between Hormone Levels and MGD
7.3. Role of PPARγ in MGD and ARMGD
7.4. Mouse Models Used to Study the Role of Oxidative Stress in MGD
7.5. Cautery-Induced Obstruction of Meibomian Gland Orifices and MGD
7.6. Desiccating Stress Induces Dry Eye-Like Symptoms and Associated Meibomian Gland Functional Changes
7.7. Association between Dyslipidemia and MGD
7.8. Role of Ectodysplasin A (EDA) in the Meibomian Gland and “Tabby” Mice
7.9. FGFR2 Signaling in Meibomian Gland Homeostasis and Potential Role in MGD
7.10. Gene Disruption of Fatty Acid (FA) Synthesis and MGD
7.11. Role of NF-κB in Meibomian Gland Development
7.12. Role of Hyaluronan in Meibomian Gland Development and Homeostasis
8. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DED | Dry eye disease |
| MG | Meibomian gland |
| MGD | Meibomian gland disease |
| ARMGD | Age-related Meibomian gland disease |
| E | Embryonic day |
| P | Post-natal day |
| SRS | Stimulated Raman scattering |
| ROS | Reactive oxygen species |
| St | Free sterols |
| FAl | Fatty alcohols |
| FA | Fatty acids |
| WE | Wax esters |
| CE | Cholesteryl esters |
| Chl | Free cholesterols |
| TAG | Triacylglycerols |
| FFA | Free fatty acids |
| PL | Phospholipids |
| Cer | Ceramides |
| VLC | Very long chains |
| ELC | Extremely long chains |
| OAHFA | (O)-acylated ω-hydroxy fatty acids |
| DiAD | Diacylated α,ω-diols |
| HE | Hematoxylin–eosin |
| NIM | Non-infrared Meibography |
| NLO | Nonlinear optical |
| GH | Growth hormone |
| IGF-1 | Insulin growth factor 1 |
| PPAR | Peroxisome proliferator-activated receptor |
| RXR | Retinoid X receptor |
| WAT | White adipose tissue |
| BAT | Brown adipose tissue |
| SOD | Superoxide dismutase |
| DS | Desiccating stress |
| APOE-KO | Apolipoprotein-E knockout |
| LDLR-KO | Low-density lipoprotein receptor knockout |
| ApoCIIIKI | Apolipoprotein CIII |
| ApoE | Apolipoprotein E |
| LDLR | Low-density lipoprotein receptor |
| VLDL | Very low-density lipoprotein |
| HDL | High-density lipoprotein |
| HFD | High-fat diet |
| OGD | Oregon green dextran |
| EDA | Ectodysplasin A |
| FGF | Fibroblast growth factor |
| FGR | Fibroblast growth receptor |
| VLCFA | Very long chain fatty acid |
| ELOVL | Elongation of very long chain fatty acid |
| TP | Tarsal plate |
| ELCFA | Extremely long chain fatty acid |
| KOMP | Knockout mouse project |
| HED | Hypohidrotic ectodermal dysplasia |
| GR | Glucocorticoid preceptor |
| K5 | Keratin 5 |
| HA | Hyaluronan |
| ECM | Extracellular matrix |
| HAS | Hyaluronan synthases |
References
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Fiona, S.; Schaumberg, D.; Uchino, M.; Vehof, J.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef] [PubMed]
- Nichols, K.K.; Foulks, G.N.; Bron, A.J.; Glasgow, B.J.; Dogru, M.; Tsubota, K.; Lemp, M.A.; Sullivan, D.A. The International Workshop on Meibomian Gland Dysfunction: Executive Summary. Investig. Opthalmol. Vis. Sci. 2011, 52, 1922–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhadva, P.; Goldhardt, R.; Galor, A. Meibomian Gland Disease. Ophthalmology 2017, 124, S20–S26. [Google Scholar] [CrossRef] [PubMed]
- Geerling, G.; Tauber, J.; Baudouin, C.; Goto, E.; Matsumoto, Y.; O’Brien, T.; Rolando, M.; Tsubota, K.; Nichols, K.K. The International Workshop on Meibomian Gland Dysfunction: Report of the Subcommittee on Management and Treatment of Meibomian Gland Dysfunction. Investig. Opthalmol. Vis. Sci. 2011, 52, 2050–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knop, E.; Knop, N.; Millar, T.; Obata, H.; Sullivan, D.A. The International Workshop on Meibomian Gland Dysfunction: Report of the Subcommittee on Anatomy, Physiology, and Pathophysiology of the Meibomian Gland. Investig. Opthalmol. Vis. Sci. 2011, 52, 1938–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jester, J.V.; Parfitt, G.J.; Brown, D.J. Meibomian gland dysfunction: Hyperkeratinization or atrophy? BMC Ophthalmol. 2015, 15, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Nien, C.J.; Paugh, J.R.; Massei, S.; Wahlert, A.J.; Kao, W.W.; Jester, J.V. Age-related changes in the meibomian gland. Exp. Eye Res. 2009, 89, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Parfitt, G.J.; Lewis, P.N.; Young, R.D.; Richardson, A.; Lyons, J.G.; Di Girolamo, N.; Jester, J.V. Renewal of the Holocrine Meibomian Glands by Label-Retaining, Unipotent Epithelial Progenitors. Stem Cell Rep. 2016, 7, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Nien, C.J.; Massei, S.; Lin, G.; Nabavi, C.; Tao, J.; Brown, D.J.; Paugh, J.R.; Jester, J.V. Effects of age and dysfunction on human meibomian glands. Arch. Ophthalmol. 2011, 129, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.S.; Parfitt, G.J.; Brown, D.J.; Jester, J.V. Meibocyte differentiation and renewal: Insights into novel mechanisms of meibomian gland dysfunction (MGD). Exp. Eye Res. 2017, 163, 37–45. [Google Scholar] [CrossRef]
- Den, S.; Shimizu, K.; Ikeda, T.; Tsubota, K.; Shimmura, S.; Shimazaki, J. Association Between Meibomian Gland Changes and Aging, Sex, or Tear Function. Cornea 2006, 25, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, B.D.; Evans, J.E.; Dana, M.R.; Sullivan, D.A. Influence of Aging on the Polar and Neutral Lipid Profiles in Human Meibomian Gland Secretions. Arch. Ophthalmol. 2006, 124, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.T.; Muntz, A.; Lim, J.; Kim, J.S.; Lacerda, L.; Arora, A.; Craig, J.P. Ageing and the natural history of dry eye disease: A prospective registry-based cross-sectional study. Ocul. Surf. 2020, 18, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Yeotikar, N.S.; Zhu, H.; Markoulli, M.; Nichols, K.K.; Naduvilath, T.J.; Papas, E. Functional and Morphologic Changes of Meibomian Glands in an Asymptomatic Adult Population. Investig. Opthalmol. Vis. Sci. 2016, 57, 3996–4007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchman, D.; Ramasubramanian, A.; Foulks, G.N. Human Meibum Cholesteryl and Wax Ester Variability with Age, Sex, and Meibomian Gland Dysfunction. Investig. Opthalmol. Vis. Sci. 2019, 60, 2286–2293. [Google Scholar] [CrossRef]
- Craig, J.P.; Wang, M.T.; Ambler, A.; Cheyne, K.R.; Wilson, G.A. Characterising the ocular surface and tear film in a population-based birth cohort of 45-year old New Zealand men and women. Ocul. Surf. 2020. [Google Scholar] [CrossRef]
- Amano, S.; Inoue, K. Estimation of Prevalence of Meibomian Gland Dysfunction in Japan. Cornea 2017, 36, 684–688. [Google Scholar] [CrossRef]
- Olami, Y.; Zajicek, G.; Cogan, M.; Gnessin, H.; Pe’Er, J. Turnover and Migration of Meibomian Gland Cells in Rats’ Eyelids. Ophthalmic Res. 2001, 33, 170–175. [Google Scholar] [CrossRef]
- Byun, T.H.; Kim, J.T.; Park, S.; Kim, W.K. Timetable for Upper Eyelid Development in Staged Human Embryos and Fetuses. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2011, 294, 789–796. [Google Scholar] [CrossRef]
- Nien, C.J.; Massei, S.; Lin, G.; Liu, H.; Paugh, J.R.; Liu, C.-Y.; Kao, W.W.-Y.; Brown, N.J.; Jester, J.V. The development of meibomian glands in mice. Mol. Vis. 2010, 16, 1132–1140. [Google Scholar]
- Kao, W.W.-Y.; Liu, H.; Zhang, J. Wakayama Symposium: Challenges of Future Research in Ocular Surface Cell Biology. Ocul. Surf. 2013, 11, 19–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A.; Borowiak, A.M.; Eule, J.C. Comparative HPLC-MSn analysis of canine and human meibomian lipidomes: Many similarities, a few differences. Sci. Rep. 2011, 1, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A. Lipidomic Analysis of Human Meibum Using HPLC–MSn. Methods Mol. Biol. 2009, 579, 221–246. [Google Scholar] [CrossRef] [PubMed]
- Arciniega, J.C.; Uchiyama, E.; Butovich, I.A. Disruption and Destabilization of Meibomian Lipid Films Caused by Increasing Amounts of Ceramides and Cholesterol. Investig. Opthalmol. Vis. Sci. 2013, 54, 1352–1360. [Google Scholar] [CrossRef] [Green Version]
- Harvey, D.; Tiffany, J. Identification of meibomian gland lipids by gas chromatography—mass spectrometry: Application to the meibomian lipids of the mouse. J. Chromatogr. A 1984, 301, 173–187. [Google Scholar] [CrossRef]
- Chen, J.; Green-Church, K.B.; Nichols, K.K. Shotgun Lipidomic Analysis of Human Meibomian Gland Secretions with Electrospray Ionization Tandem Mass Spectrometry. Investig. Opthalmol. Vis. Sci. 2010, 51, 6220–6231. [Google Scholar] [CrossRef]
- Nicolaides, N.; Kaitaranta, J.K.; Rawdah, T.N.; Macy, J.; Boswell, F.M.; Smith, R. Meibomian gland studies: Comparison of steer and human lipids. Investig. Ophthalmol. Vis. Sci. 1981, 20, 522–536. [Google Scholar]
- Suhalim, J.L.; Parfitt, G.J.; Xie, Y.; De Paiva, C.S.; Pflugfelder, S.C.; Shah, T.N.; Potma, E.O.; Brown, D.J.; Jester, J.V. Effect of desiccating stress on mouse meibomian gland function. Ocul. Surf. 2013, 12, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Morishige, N.; Arita, R.; Koh, S.; Sakimoto, T.; Shirakawa, R.; Miyata, K.; Ohashi, Y. Morphological changes in the meibomian glands of patients with phlyctenular keratitis: A multicenter cross-sectional study. BMC Ophthalmol. 2016, 16, 178. [Google Scholar] [CrossRef] [Green Version]
- Dogru, M.; Kojima, T.; Şimşek, C.; Tsubota, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Opthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [Green Version]
- DeNoyer, A.; Landman, E.; Trinh, L.; Faure, J.-F.; Auclin, F.; Baudouin, C. Dry Eye Disease after Refractive Surgery. Ophthalmology 2015, 122, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Viñas, M.; Maggio, F.; D’Anna, N.; Rabozzi, R.; Peruccio, C. Meibomian gland dysfunction (MGD), as diagnosed by non-contact infrared Meibography, in dogs with ocular surface disorders (OSD): A retrospective study. BMC Veter Res. 2019, 15, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, Y.; Arita, R.; Miwa, Y.; Iwashita, H.; Saito, A. Histopathologic changes associated with Meibomian Gland dropout in a dog. Veter Ophthalmol. 2020, 23, 575–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartori, R.; Peruccio, C. A Case of Sebaceous Adenitis and Concurrent Meibomian Gland Dysfunction in a Dog. Veter Sci. 2020, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; Lu, H.; McMahon, A.; Eule, J.C. Toward an Animal Model of the Human Tear Film: Biochemical Comparison of the Mouse, Canine, Rabbit, and Human Meibomian Lipidomes. Investig. Opthalmol. Vis. Sci. 2012, 53, 6881–6896. [Google Scholar] [CrossRef]
- Steagall, R.J.; Yamagami, H.; Wickham, L.A.; Sullivan, D.A. Androgen Control of Gene Expression in the Rabbit Meibomian Gland. Adv. Exp. Med. Biol. 2002, 506, 465–476. [Google Scholar] [CrossRef]
- Sullivan, D.A.; Jensen, R.V.; Suzuki, T.; Richards, S.M. Do sex steroids exert sex-specific and/or opposite effects on gene expression in lacrimal and meibomian glands? Mol. Vis. 2009, 15, 1553. [Google Scholar]
- Jester, J.V.; Rajagopalan, S.; Rodrigues, M. Meibomian gland changes in the rhino (hr(rh)hr(rh)) mouse. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1190–1194. [Google Scholar]
- Jester, J.V.; Potma, E.; Brown, D.J. PPARγ Regulates Mouse Meibocyte Differentiation and Lipid Synthesis. Ocul. Surf. 2016, 14, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Yoon, C.H.; Ryu, J.S.; Hwang, H.S.; Kim, M.K. Comparative Analysis of Age-Related Changes in Lacrimal Glands and Meibomian Glands of a C57BL/6 Male Mouse Model. Int. J. Mol. Sci. 2020, 21, 4169. [Google Scholar] [CrossRef]
- Butovich, I.A. Meibomian glands, meibum, and meibogenesis. Exp. Eye Res. 2017, 163, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Arita, R.; Morishige, N.; Koh, S.; Shirakawa, R.; Kawashima, M.; Sakimoto, T.; Suzuki, T.; Tsubota, K. Increased Tear Fluid Production as a Compensatory Response to Meibomian Gland Loss. Ophthalmology 2015, 122, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A. On the Presence and Role of Polar Lipids in Meibum. Investig. Opthalmol. Vis. Sci. 2010, 51, 6908–6910. [Google Scholar] [CrossRef] [PubMed]
- Butovich, I.A.; Wojtowicz, J.C.; Molai, M. Human tear film and meibum. Very long chain wax esters and (O-acyl)-omega-hydroxy fatty acids of meibum. J. Lipid Res. 2009, 50, 2471–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A.; Arciniega, J.C.; Lu, H.; Molai, M. Evaluation and Quantitation of Intact Wax Esters of Human Meibum by Gas-Liquid Chromatography-Ion Trap Mass Spectrometry. Investig. Opthalmol. Vis. Sci. 2012, 53, 3766–3781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pochi, P.E.; Strauss, J.S.; Downing, D.T. Age-related Changes in Sebaceous Gland Activity. J. Investig. Dermatol. 1979, 73, 108–111. [Google Scholar] [CrossRef] [Green Version]
- Zoubouliss, C.C.; Boschnakow, A. Chronological ageing and photoageing of the human sebaceous gland. Clin. Exp. Dermatol. 2001, 26, 600–607. [Google Scholar] [CrossRef]
- Obata, H. Anatomy and Histopathology of Human Meibomian Gland. Cornea 2002, 21, S70–S74. [Google Scholar] [CrossRef]
- Jester, B.E.; Nien, C.J.; Winkler, M.; Brown, D.J.; Jester, J.V. Volumetric Reconstruction of the Mouse Meibomian Gland Using High-Resolution Nonlinear Optical Imaging. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2010, 294, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Parfitt, G.J.; Xie, Y.; Reid, K.M.; Brown, D.J.; Jester, J.V. Three-Dimensional Immunohistochemical Multiplexed High-Resolution Macroscopy (IM-HRMac) of the Meibomian Gland. Investig. Ophthalmol. Vis. Sci. 2012, 53, 592. [Google Scholar]
- Parfitt, G.J.; Xie, Y.; Reid, K.M.; Dervillez, X.; Brown, D.J.; Jester, J.V. A Novel Immunofluorescent Computed Tomography (ICT) Method to Localise and Quantify Multiple Antigens in Large Tissue Volumes at High Resolution. PLoS ONE 2012, 7, e53245. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.S.; Mikula, E.; Xie, Y.; Brown, D.J.; Jester, J.V. A novel transillumination meibography device for in vivo imaging of mouse meibomian glands. Ocul. Surf. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.A.; Sullivan, B.D.; Ullman, M.D.; Rocha, E.M.; Krenzer, K.L.; Cermak, J.M.; Toda, I.; Doane, M.G.; Evans, J.E.; Wickham, L.A. Androgen influence on the meibomian gland. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3732–3742. [Google Scholar]
- Rocha, E.M. Identification of androgen receptor protein and 5alpha -reductase mRNA in human ocular tissues. Br. J. Ophthalmol. 2000, 84, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Yamagami, H.; Schirra, F.; Liu, M.; Richards, S.M.; Sullivan, B.D.; Sullivan, D.A. Androgen influence on gene expression in the meibomian gland. Atherosclerosis 2002, 506, 477–481. [Google Scholar] [CrossRef]
- Kopchick, J.J.; Andry, J.M. Growth Hormone (GH), GH Receptor, and Signal Transduction. Mol. Genet. Metab. 2000, 71, 293–314. [Google Scholar] [CrossRef]
- Ding, J.; Sullivan, D.A. The Effects of Insulin-like Growth Factor 1 and Growth Hormone on Human Meibomian Gland Epithelial Cells. JAMA Ophthalmol. 2014, 132, 593. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D. Evolutionary Medicine: From Dwarf Model Systems to Healthy Centenarians? Science 2003, 299, 1342–1346. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kam, W.R.; Ding, J.; Sullivan, D.A. Effect of azithromycin on lipid accumulation in immortalized human meibomian gland epithelial cells. JAMA Ophthalmol. 2014, 132, 226–228. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Knop, E.; Knop, N.; Sullivan, D.A.; List, E.O.; Kopchick, J.J.; Kam, W.R.; Ding, J. Growth Hormone Influence on the Morphology and Size of the Mouse Meibomian Gland. J. Ophthalmol. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Menendez-Gutierrez, M.P.; Röszer, T.; Ricote, M. Biology and therapeutic applications of peroxisome proliferator- activated receptors. Curr. Top. Med. Chem. 2012, 12, 548–584. [Google Scholar] [CrossRef] [PubMed]
- Barish, G.D.; Narkar, V.A.; Evans, R.M. PPAR: A dagger in the heart of the metabolic syndrome. J. Clin. Investig. 2006, 116, 590–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, S.G.; Laganà, A.S.; Nigro, A.; La Rosa, V.L.; Rossetti, P.; Rapisarda, A.M.C.; La Vignera, S.; Condorelli, R.A.; Corrado, F.; Buscema, M.; et al. Peroxisome Proliferator-Activated Receptor Modulation during Metabolic Diseases and Cancers: Master and Minions. PPAR Res. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, R.M.; Barrish, G.D.; Wang, Y.-X. PPARs and the complex journey to obesity. Nat. Med. 2004, 10, 355–361. [Google Scholar] [CrossRef]
- Sugii, S.; Evans, R.M. Epigenetic codes of PPARγ in metabolic disease. FEBS Lett. 2011, 585, 2121–2128. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Xie, Y.; Nguyen, P.Q.; Bui, V.T.; Huynh, K.; Kang, J.S.; Brown, D.J.; Jester, J.V. PPARγ regulates meibocyte differentiation and lipid synthesis of cultured human meibomian gland epithelial cells (hMGEC). Ocul. Surf. 2018, 16, 463–469. [Google Scholar] [CrossRef]
- Jester, J.V.; Brown, D.J. Wakayama Symposium: Peroxisome Proliferator-Activated Receptor-Gamma (PPARγ) and Meibomian Gland Dysfunction. Ocul. Surf. 2012, 10, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Rho, C.R.; Kim, J.; Xie, Y.; Prince, R.C.; Mustafa, K.; Potma, E.O.; Brown, D.J.; Jester, J.V. Eicosapentaenoic acid (EPA) activates PPARγ signaling leading to cell cycle exit, lipid accumulation, and autophagy in human meibomian gland epithelial cells (hMGEC). Ocul. Surf. 2020, 18, 427–437. [Google Scholar] [CrossRef]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Halliwell, B. Cell culture, oxidative stress, and antioxidants: Avoiding pitfalls. Biomed. J. 2014, 37, 99–105. [Google Scholar] [CrossRef]
- Crapo, J.D.; Oury, T.; Rabouille, C.; Slot, J.W.; Chang, L.Y. Copper, zinc superoxide dismutase is primarily a cytosolic protein in human cells. Proc. Natl. Acad. Sci. USA 1992, 89, 10405–10409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [PubMed]
- Imamura, Y.; Noda, S.; Hashizume, K.; Shinoda, K.; Yamaguchi, M.; Uchiyama, S.; Shimizu, T.; Mizushima, Y.; Shirasawa, T.; Tsubota, K. Drusen, choroidal neovascularization, and retinal pigment epithelium dysfunction in SOD1-deficient mice: A model of age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2006, 103, 11282–11287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, T.; Wakamatsu, T.H.; Dogru, M.; Ogawa, Y.; Igarashi, A.; Ibrahim, O.M.; Inaba, T.; Shimizu, T.; Noda, S.; Obata, H.; et al. Age-Related Dysfunction of the Lacrimal Gland and Oxidative Stress. Am. J. Pathol. 2012, 180, 1879–1896. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, O.M.; Dogru, M.; Matsumoto, Y.; Igarashi, A.; Kojima, T.; Wakamatsu, T.H.; Inaba, T.; Shimizu, T.; Shimazaki, J.; Tsubota, K. Oxidative Stress Induced Age Dependent Meibomian Gland Dysfunction in Cu, Zn-Superoxide Dismutase-1 (Sod1) Knockout Mice. PLoS ONE 2014, 9, e99328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbard, J.P.; Rossi, S.R.; Heyda, K.G. Tear Film and Ocular Surface Changes after Closure of the Meibomian Gland Orifices in the Rabbit. Ophthalmology 1989, 96, 1180–1186. [Google Scholar] [CrossRef]
- Nichols, K.K.; Hanlon, S.D.; Nichols, J.J. A Murine Model for Characterizing Glandular Changes in Obstructive Meibomian Gland Dysfunction. Investig. Ophthalmol. Vis. Sci. 2014, 55, 14. [Google Scholar]
- Dong, Z.-Y.; Ying, M.; Zheng, J.; Hu, L.-J.; Xie, J.-Y.; Ma, Y. Evaluation of a rat meibomian gland dysfunction model induced by closure of meibomian gland orifices. Int. J. Ophthalmol. 2018, 11, 1077–1083. [Google Scholar] [CrossRef]
- Foulks, G.N.; Bron, A.J. Meibomian Gland Dysfunction: A Clinical Scheme for Description, Diagnosis, Classification, and Grading. Ocul. Surf. 2003, 1, 107–126. [Google Scholar] [CrossRef]
- Dursun, D.; Wang, M.; Monroy, D.; Li, D.-Q.; Lokeshwar, B.L.; Stern, M.; Pflugfelder, S.C. Experimentally induced dry eye produces ocular surface inflammation and epithelial disease. Atherosclerosis 2002, 506, 647–655. [Google Scholar] [CrossRef]
- Dursun, D.; Wang, M.; Monroy, D.; Li, D.-Q.; Lokeshwar, B.L.; Stern, M.E.; Pflugfelder, S.C. A mouse model of keratoconjunctivitis sicca. Investig. Ophthalmol. Vis. Sci. 2002, 43, 632–638. [Google Scholar]
- Stern, M.E.; Beuerman, R.W.; Fox, R.I.; Gao, J.; Mircheff, A.K.; Pflugfelder, S.C. The Pathology of Dry Eye. Cornea 1998, 17, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Suhalim, J.L.; Nien, C.L.; Miljković, M.D.; Diem, M.; Jester, J.V.; Potma, E.O. Picosecond spectral coherent anti-Stokes Raman scattering imaging with principal component analysis of meibomian glands. J. Biomed. Opt. 2011, 16, 21104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniappan, C.K.; Schütt, B.S.; Bräuer, L.; Schicht, M.; Millar, T.J. Effects of Keratin and Lung Surfactant Proteins on the Surface Activity of Meibomian Lipids. Investig. Opthalmol. Vis. Sci. 2013, 54, 2571–2581. [Google Scholar] [CrossRef] [Green Version]
- Dao, A.H.; Spindle, J.D.; Harp, B.A.; Jacob, A.; Chuang, A.Z.; Yee, R.W. Association of Dyslipidemia in Moderate to Severe Meibomian Gland Dysfunction. Am. J. Ophthalmol. 2010, 150, 371–375.e1. [Google Scholar] [CrossRef]
- Braich, P.S.; Howard, M.K.; Singh, J.S. Dyslipidemia and its association with meibomian gland dysfunction. Int. Ophthalmol. 2015, 36, 469–476. [Google Scholar] [CrossRef]
- Módulo, C.M.; Filho, E.B.M.; Malki, L.T.; Dias, A.C.; De Souza, J.C.; Oliveira, H.C.F.; Jorge Ítalo, C.; Gomes, I.B.S.; Meyrelles, S.S.; Rocha, E.M. The Role of Dyslipidemia on Ocular Surface, Lacrimal and Meibomian Gland Structure and Function. Curr. Eye Res. 2012, 37, 300–308. [Google Scholar] [CrossRef]
- Osae, E.A.; Steven, P.; Redfern, R.; Hanlon, S.; Smith, C.W.; Rumbaut, R.E.; Burns, A.R. Dyslipidemia and Meibomian Gland Dysfunction: Utility of Lipidomics and Experimental Prospects with a Diet-Induced Obesity Mouse Model. Int. J. Mol. Sci. 2019, 20, 3505. [Google Scholar] [CrossRef] [Green Version]
- Bu, J.; Wu, Y.; Cai, X.; Jiang, N.; Jeyalatha, M.V.; Yu, J.; He, X.; He, H.; Guo, Y.; Zhang, M.; et al. Hyperlipidemia induces meibomian gland dysfunction. Ocul. Surf. 2019, 17, 777–786. [Google Scholar] [CrossRef]
- Blue, M.L.; Williams, D.L.; Zucker, S.W.; Khan, S.A.; Blum, C.B. Apolipoprotein E synthesis in human kidney, adrenal gland, and liver. Proc. Natl. Acad. Sci. USA 1983, 80, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Osada, J.; Joven, J.; Maeda, N. The value of apolipoprotein E knockout mice for studying the effects of dietary fat and cholesterol on atherogenesis. Curr. Opin. Lipidol. 2000, 11, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Pinna, A.; Piccinini, P.; Carta, F. Effect of Oral Linoleic and γ-Linolenic Acid on Meibomian Gland Dysfunction. Cornea 2007, 26, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Tomozawa, J.; Mizutani, N.; Nabe, T.; Danno, K.; Kohno, S. Atopic dermatitis-like pruritic skin inflammation caused by feeding a special diet to HR-1 hairless mice. Exp. Dermatol. 2005, 14, 460–468. [Google Scholar] [CrossRef]
- Miyake, H.; Oda, T.; Katsuta, O.; Seno, M.; Nakamura, M. Meibomian Gland Dysfunction Model in Hairless Mice Fed a Special Diet With Limited Lipid Content. Investig. Opthalmol. Vis. Sci. 2016, 57, 3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Wu, J.; Bu, J.; Tang, L.; Yang, Y.; Ouyang, W.; Lin, X.; Liu, Z.; Huang, C.; Quantock, A.J.; et al. High-fat diet induces dry eye-like ocular surface damages in murine. Ocul. Surf. 2020, 18, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Pispa, J.; Hartung, A.J.; Du, Y.; Ezer, S.; Jenks, T.; Shimada, T.; Pekkanen, M.; Mikkola, M.L.; Ko, M.S.H.; et al. The Tabby phenotype is caused by mutation in a mouse homologue of the EDA gene that reveals novel mouse and human exons and encodes a protein (ectodysplasin-A) with collagenous domains. Proc. Natl. Acad. Sci. USA 1997, 94, 13069–13074. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.-Y.; Smith, J.A.; Schlessinger, D.; Chan, C.-C. X-Linked Anhidrotic Ectodermal Dysplasia Disruption Yields a Mouse Model for Ocular Surface Disease and Resultant Blindness. Am. J. Pathol. 2005, 167, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-C.; Li, S.; Chen, X.; Ma, B.; He, H.; Liu, T.; Yu, J.; Zhang, L.; Chen, Y.; Liu, Z.; et al. Meibomian Gland Absence Related Dry Eye in Ectodysplasin A Mutant Mice. Am. J. Pathol. 2016, 186, 32–42. [Google Scholar] [CrossRef]
- Yun, Y.-R.; Won, J.E.; Jeon, E.; Lee, S.; Kang, W.; Jo, H.; Jang, J.-H.; Shin, U.S.; Kim, H.-W. Fibroblast Growth Factors: Biology, Function, and Application for Tissue Regeneration. J. Tissue Eng. 2010, 1, 218142. [Google Scholar] [CrossRef]
- Korsensky, L.; Ron, D. Regulation of FGF signaling: Recent insights from studying positive and negative modulators. Semin. Cell Dev. Biol. 2016, 53, 101–114. [Google Scholar] [CrossRef]
- Scalinci, S.Z.; Scorolli, L.; Meduri, A.; Grenga, P.L.; Corradetti, G. Effect of basic fibroblast growth factor and cytochrome c peroxidase combination in transgenic mice corneal epithelial healing process after excimer laser photoablation. Clin. Ophthalmol. 2011, 5, 215–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meduri, A.; Scalinci, S.Z.; Morara, M.; Ceruti, P.; Grenga, P.L.; Zigiotti, G.L.; Scorolli, L. Effect of Basic Fibroblast Growth Factor in Transgenic Mice: Corneal Epithelial Healing Process after Excimer Laser Photoablation. Ophthalmologica 2008, 223, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Grose, R.P.; Fantl, V.; Werner, S.; Chioni, A.-M.; Jarosz, M.; Rudling, R.; Cross, B.; Hart, I.R.; Dickson, C. The role of fibroblast growth factor receptor 2b in skin homeostasis and cancer development. EMBO J. 2007, 26, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Reneker, L.W.; Wang, L.; Irlmeier, R.T.; Huang, A.J.W. Fibroblast Growth Factor Receptor 2 (FGFR2) Is Required for Meibomian Gland Homeostasis in the Adult Mouse. Investig. Opthalmol. Vis. Sci. 2017, 58, 2638–2646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bron, A.J.; Tomlinson, A.; Foulks, G.N.; Pepose, J.S.; Baudouin, C.; Geerling, G.; Nichols, K.K.; Lemp, M.A. Rethinking Dry Eye Disease: A Perspective on Clinical Implications. Ocul. Surf. 2014, 12, S1–S31. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Suto, S.; Yamanaka, M.; Mizutani, Y.; Mitsutake, S.; Igarashi, Y.; Sassa, T.; Kihara, A. ELOVL1 production of C24 acyl-CoAs is linked to C24 sphingolipid synthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 18439–18444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassa, T.; Tadaki, M.; Kiyonari, H.; Kihara, A. Very long-chain tear film lipids produced by fatty acid elongase ELOVL1 prevent dry eye disease in mice. FASEB J. 2018, 32, 2966–2978. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; Wilkerson, A.; Bhat, N.; McMahon, A.; Yuksel, S. On the pivotal role of Elovl3 /ELOVL3 in meibogenesis and ocular physiology of mice. FASEB J. 2019, 33, 10034–10048. [Google Scholar] [CrossRef]
- Hopiavuori, B.R.; Anderson, R.E.; Agbaga, M.-P. ELOVL4: Very long-chain fatty acids serve an eclectic role in mammalian health and function. Prog. Retin. Eye Res. 2018, 69, 137–158. [Google Scholar] [CrossRef]
- McMahon, A.; Lu, H.; Butovich, I.A. A Role for ELOVL4 in the Mouse Meibomian Gland and Sebocyte Cell Biology. Investig. Opthalmol. Vis. Sci. 2014, 55, 2832–2840. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.B.; Russell, D.W. Mammalian Wax Biosynthesis. J. Biol. Chem. 2004, 279, 37789–37797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundberg, J.P.; Dadras, S.S.; Silva, K.A.; Kennedy, V.E.; Garland, G.; Murray, S.A.; Sundberg, B.A.; Schofield, P.N.; Pratt, C.H. Systematic screening for skin, hair, and nail abnormalities in a large-scale knockout mouse program. PLoS ONE 2017, 12, e0180682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundberg, J.P.; Shen, T.; Fiehn, O.; Rice, R.H.; Silva, K.A.; Kennedy, V.E.; Gott, N.E.; Dionne, L.A.; Bechtold, L.S.; Murray, S.A.; et al. Sebaceous gland abnormalities in fatty acyl CoA reductase 2 (Far2) null mice result in primary cicatricial alopecia. PLoS ONE 2018, 13, e0205775. [Google Scholar] [CrossRef] [PubMed]
- Niemann, C.; Horsley, V. Development and homeostasis of the sebaceous gland. Semin. Cell Dev. Biol. 2012, 23, 928–936. [Google Scholar] [CrossRef] [Green Version]
- Lamartine, J. Towards a new classification of ectodermal dysplasias. Clin. Exp. Dermatol. 2003, 28, 351–355. [Google Scholar] [CrossRef]
- Mikkola, M.L.; Thesleff, I. Ectodysplasin signaling in development. Cytokine Growth Factor Rev. 2003, 14, 211–224. [Google Scholar] [CrossRef]
- Schmidt-Ullrich, R.; Aebischer, T.; Hülsken, J.; Birchmeier, W.; Klemm, U.; Scheidereit, C. Requirement of NF-κB/Rel for the development of hair follicles and other epidermal appendices. Development 2001, 19, 3843–3853. [Google Scholar]
- Pérez, P.; Cascallana, J.L. Disruption of eyelid and cornea development by targeted overexpression of the glucocorticoid receptor. Int. J. Dev. Biol. 2003, 47, 59–64. [Google Scholar] [CrossRef]
- Cascallana, J.L.; Bravo, A.; Donet, E.; Leis, H.; Lara, M.F.; Paramio, J.M.; Jorcano, J.L.; Pérez, P. Ectoderm-Targeted Overexpression of the Glucocorticoid Receptor Induces Hypohidrotic Ectodermal Dysplasia. Endocrinology 2005, 146, 2629–2638. [Google Scholar] [CrossRef] [Green Version]
- Nelson, G.; Wilde, G.J.C.; Spiller, D.G.; Kennedy, S.M.; Ray, D.W.; Sullivan, E.; Unitt, J.F.; White, M.R.H. NF-kappaB signalling is inhibited by glucocorticoid receptor and STAT6 via distinct mechanisms. J. Cell Sci. 2003, 116, 2495–2503. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Puri, S.; Parfitt, G.J.; Mutoji, N.; Coulson-Thomas, V.J. Hyaluronan Regulates Eyelid and Meibomian Gland Morphogenesis. Investig. Opthalmol. Vis. Sci. 2018, 59, 3713–3727. [Google Scholar] [CrossRef] [PubMed]
- Gesteira, T.F.; Sun, M.; Coulson-Thomas, Y.M.; Yamaguchi, Y.; Yeh, L.-K.; Hascall, V.; Coulson-Thomas, V.J. Hyaluronan Rich Microenvironment in the Limbal Stem Cell Niche Regulates Limbal Stem Cell Differentiation. Investig. Opthalmol. Vis. Sci. 2017, 58, 4407–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Model | Description | Animal | MGD Features | Other Defects |
|---|---|---|---|---|
| Yagyu, H. et al. 2000. | Elimination of Acyl-CoA:cholesterol acyltransferase gene 1 (ACAT-1). | ACAT-1 null (ACAT-1−/−) mice | Narrow eye fissures and Meibomian gland atrophy. | Lipid-depleted adrenal glands and corneal erosion. |
| Pikus et al. 2004. | Modulation of bone morphogenic protein (BMP). | K14-Noggin transgenic mouse | Formation of pilosebaceous units at the expense of Meibomian glands/suppression of the induction of Meibomian glands. | Abnormal sweat glands, ectopic cilia, distal limb agenesis, hyperpigmentation of claws, interdigital webbing, and reduced footpads. |
| Cascallana et al. 2005. | Overexpression of glucocorticoid receptor (GR). | keratin 5 (K5)-GR mice | Lack of Meibomian glands. | Underdeveloped sweat glands and preputial glands, and abnormal hair follicles, teeth, and palate. |
| Cui, C.Y. et al. 2005; Cui, C.Y. et al. 2010; Wang, Y.C. et al. 2016; Li, S. et al. 2018. | Mutation of X-linked anhidrotic ectodermal dysplasia (EDA) gene. | Eda mutant (Eda−/−) Tabby mice | Lack of Meibomian glands, reduced tear break up times, and blepharitis. | Corneal neovascularization, ulceration, keratinization, reduced corneal epithelial microvilli, and conjunctivitis. |
| Vauclair, S. et al. 2007. | Skin-specific inactivation of the Notch1 gene. | Notch1K14Cre mice; Notch1 K5CreERT mice | Meibomian gland dysfunction, abnormal morphology of Meibomian glands, lack of lipids in meibocytes. | Eye irritations, corneal opacity and keratinization. |
| Chang et al. 2009. | Elevation of EDAR signaling. | EdarTg951 heterozygous transgenic and EdarTg951/Tg951 homozygous transgenic mice | Enlarged Meibomian glands. | Excessively branched mammary and salivary glands. |
| Tukel, T. et al. 2010. | Homozygous TWIST2 (MIM 607556) nonsense mutations. | Twist2 knockout (KO) mice | Absence of or hypoplastic Meibomian glands and decreased eyelash follicles. | Bitemporal lesions, narrow snout, pointed chin, and sparse or absent eyelashes. |
| Kenchegowda, D. et al. 2011. | Conditional deletion of Krüppel-like factor (KLF) 5. | Klf5-conditional null (Klf5CN) mice | Malformed Meibomian glands with disorganized acini, lipid accumulation in the Meibomian ducts. | Smaller eyeballs; translucent and thicker corneas with a defective epithelial basement membrane and hypercellular stroma; conjunctivas lacking goblet cells. |
| Tsau, C. et al. 2011. | Deletion of the sequences encoding the homeodomain and C-terminal region of Barx2. | Barx2-null mice | ~50% of Barx2–/– mice have eyelid fusion problems, smaller eyes, and defects in Meibomian gland development and structure including Meibomian gland drop out or smaller glands. | Defective lacrimal gland morphogenesis, and absence of the harderian gland. |
| Lin et al. 2013. | Mice lacking fatty acid transport protein (FATP) 4. | Tg (IVL-Fatp4) transgenic mice | Underdeveloped Meibomian glands with thickened ducts. | Abnormal sebaceous glands, and thick skin with defective barrier. |
| McMahon et al. 2014. | Mutation in ELOVL4 resulted in abnormal synthesis of extremely long-chain fatty acids. | Heterozygous Stgd3 mice, on a mixed 129SvEv and C57BL6 background | Protruding Meibomian gland orifice, intragland anatomical changes, toothpaste-like meibum, and intense staining for ELOVL4 in glands. | Inability to open eyes fully, and increased blink rates. |
| Chen, Z.Y. et al. 2014. | Conditional deletion of Sox9. | Sox9CKO mice | Reduced number of Meibomian glands, 40% fewer glands in the upper and lower eyelids, and most Meibomian glands have fewer acini. | Lacrimal gland (LG) deficiency and abnormal LG morphogenesis, absence of harderian glands, and hair loss in mouse eyelids and facial skin. |
| Meng, Q. et al. 2014. | Targeted gene ablation that inactivated distinct signaling pathways. | Map3k1-null and Dkk2-null mice, K14rtTA/tet-O-cre/Shp2F/F mice, c-JunΔ/ΔOSE and EgfrΔ/ΔOSE mice, Map3k1/Jnk1 and Map3k1/Rhoa compound mutants | Eyelid fusion problems and severe hypoplastic Meibomian glands. | Corneal erosion/ulceration, harderian gland hypoplasia, misplacement of extraocular muscles and eyes open at birth. |
| Ibrahim, O.M. et al. 2014; Ikeda et al. 2018. | Deficient in Cu/Zn superoxide dismutase (SOD1) leading to accumulation of reactive oxygen species (ROS). | Sod1−/− mice | Age-related Meibomian gland abnormalities including an increase in periglandular inflammatory infiltrates, decrease in Meibomian gland glandular acinar density, and increase in periglandular fibrosis. | Corneal fluorescein and lissamine staining evidencing corneal erosions and reduced tear secretion. |
| Dong, F. et al. 2015. | Conditional inducible ablation of TGFα in the eyelid. | Bi-transgenic Kera-rtTA/tetO-TGFα (KR/TG) mice | Abnormal Meibomian glandmorphogenesis, Meibomian gland atrophy, and eyelid tendon and tarsal plate malformation. | Precocious eye opening, swollen eyelids, and conjunctival eyelid epithelial hyperplasia. |
| Sima, J. et al. 2016; Cui, C.Y. et al. 2010. | Evaluation of Dickkopf 4 (DKK4) regulated signaling pathway. | Skin-specific Dkk4 transgenic mice (WTDk4TG, wildtype background) and TaDk4TG (Tabby background) | Meibomian gland formation defects in Dkk4Tg mice, similar with Tabby mice. | Cataracts, corneal blindness and rough hair coat. |
| Miyake et al. 2016. | HR-AD diet (a special diet with limited lipid content). | HR-1 hairless mice (fed with HR-AD diet) | Plugged Meibomian gland orifice, Meibomian gland ductal epithelial hyperkeratinization and acinar atrophy. | No other phenotypes recorded. |
| Reneker, L.W. et al. 2017; Chen, Z.Y. et al. 2014. | Conditional inducible ablation of FGFR2. | Inducible Fgfr2CKO mice | Macerated periorbital hairs and eyelids, Meibomian gland orifice obstruction, reduced volume of meibum and Meibomian gland acinar atrophy. | Ocular irritation and rubbing. |
| Yu, D. et al. 2018. | Deletion of epithelial sodium channel (ENaC) functional subunits in the Meibomian gland. | Conditional βENaC Meibomian gland knockout (KO) mouse (βENaCfl/fl;Shh-Cre+/−) | Age-dependent, female-predominant Meibomian gland dysfunction (obstruction of Meibomian gland orifices and Meibomian gland acinar atrophy). | Increased tear secretion and severe ocular surface damage (corneal opacity, ulceration, neovascularization and ectasia). |
| Swirski, S. et al. 2018. | Mutations in the gene encoding Gasdermin A3 (Gsdma3). | C+/H− Mice | Degeneration of Meibomian glands, increased eyelid tissue formation and eyelid closure. | Progressive hair loss, hyperkeratinization of the skin, degeneration of sebaceous glands, corneal opacity, and corenal vascularization. |
| Sassa, T. et al. 2018. | Gene disruption of Elovl1 resulted in insufficient elongation of FAs. | Elovl1−/− Tg(IVL-Elovl1) mice | Evaporative dry eye phenotype with increased eye-blink frequency, together with partially closed eyes and excessive tear production. | Corneal opacity, vascularization and epidermalization in aged mice. |
| Sun, M. et al. 2018; Gesteira, T.F. et al. 2017. | Targeted knockout of Hyaluronan synthase (Has) genes 1, 2 and 3. | Combined Has1−/−;Has3−/− mice; conditional Has2Δ/ΔMG mice | Meibomian gland hyperplasia with more branched, longer and wider acini. | Dysmorphic eyelids with conjunctival epithelial hyperplasia and thinner corneal thickness. |
| Swirski, S. et al. 2018; Miyazaki, M. et al. 2001. | Targeted knockout of Stearoyl-CoA desaturase (SCD) 1. | SCD-1 knockout mice (SCD-1−/−) | Narrow eye fissures, atrophy or loss of Meibomian glands and depletion of meibum lipids. | Cutaneous abnormalities including thinner hair coat and hair loss, with atrophic sebaceous gland abnormalities and compensatory increases in tear volume and mucin levels. |
| Sundberg, J.P. et al. 2018. | Null mice for fatty acyl CoA reductase 2 (Far2) gene altering saturated 16:0 and 18:0 carbon fatty acids. | Homozygous mutant Far2tm2b(KOMP)Wtsi/2J (Far2−/−; null) | Markedly dilated Meibomian gland ducts, and abnormal sebocytes with clustered eosinophilic remnants. | Patchy alopecia on the dorsal trunk and diffuse hair thinning on the ventral body surface. |
| Butovich, I.A. et al. 2019. | Loss of ELOVL3 altering the synthesis of C21:0-C29:0 fatty acids. | Elovl3 knockout (E3hom) mice | Delayed eye opening, weeping eyes, crusty eyelids, eyelid edema, highly vascularized cornea and tarsal plates (TPs), slit eye, and excessive tearing. | Hairless pups and greasy fur in adults. |
| Wu, L. et al. 2019. | Null SDR16C5 and SDR16C6 mice with altered retinoic acid (RA) biosynthesis. | Sdr16c5/Sdr16c6 double-knockout mice (DKO) | Both the upper and lower eyelids are thicker and longer (eyelid expansion) with larger Meibomian glands. | Accelerated hair regrowth. |
| Widjaja-Adhi, M.A.K. et al. 2020. | Ablation of Acyl-CoA wax alcohol acyltransferase 2 (AWAT2) enzymatic activity. | Awat2-knockout mice (Awat2−/−) | Obstruction of the orifice and excretory duct of Meibomian glands, hyperkeratinization of the epithelium, absence of wax esters and overproduction of cholesteryl esters. | Shorter tear film break-up time, deterioration of the corneal surface, granular deposits in the corneal stromal layer, corneal neovascularization, blepharitis, Iritis, scaly and dry skin, and smaller sebaceous glands. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Moreno, I.Y.; Dang, M.; Coulson-Thomas, V.J. Meibomian Gland Dysfunction: What Have Animal Models Taught Us? Int. J. Mol. Sci. 2020, 21, 8822. https://doi.org/10.3390/ijms21228822
Sun M, Moreno IY, Dang M, Coulson-Thomas VJ. Meibomian Gland Dysfunction: What Have Animal Models Taught Us? International Journal of Molecular Sciences. 2020; 21(22):8822. https://doi.org/10.3390/ijms21228822
Chicago/Turabian StyleSun, Mingxia, Isabel Y. Moreno, Michelle Dang, and Vivien J. Coulson-Thomas. 2020. "Meibomian Gland Dysfunction: What Have Animal Models Taught Us?" International Journal of Molecular Sciences 21, no. 22: 8822. https://doi.org/10.3390/ijms21228822
APA StyleSun, M., Moreno, I. Y., Dang, M., & Coulson-Thomas, V. J. (2020). Meibomian Gland Dysfunction: What Have Animal Models Taught Us? International Journal of Molecular Sciences, 21(22), 8822. https://doi.org/10.3390/ijms21228822