Acetaminophen-Induced Rat Hepatotoxicity Based on M1/M2-Macrophage Polarization, in Possible Relation to Damage-Associated Molecular Patterns and Autophagy

 ,
,
Abstract
:1. Introduction
2. Results
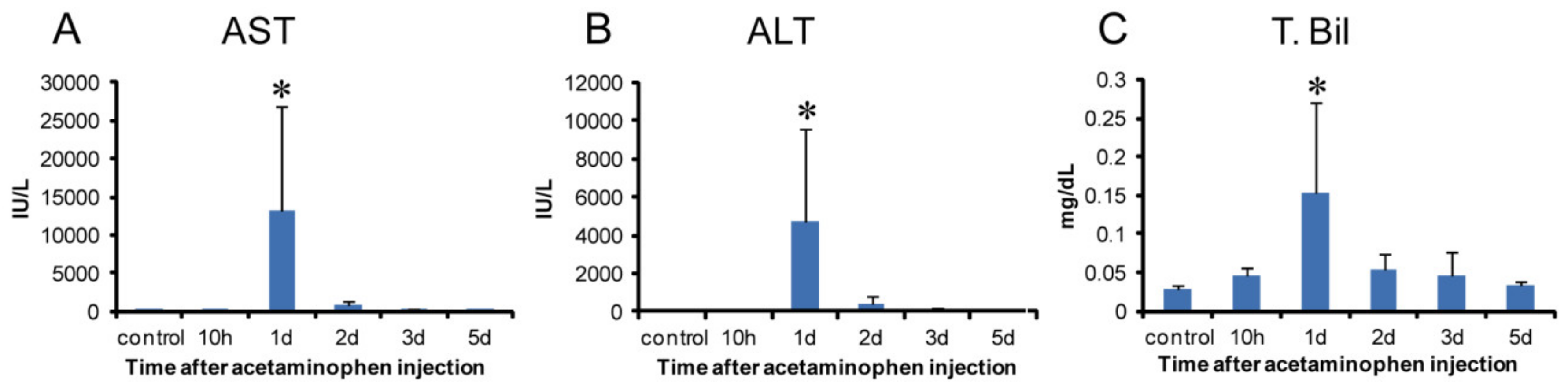
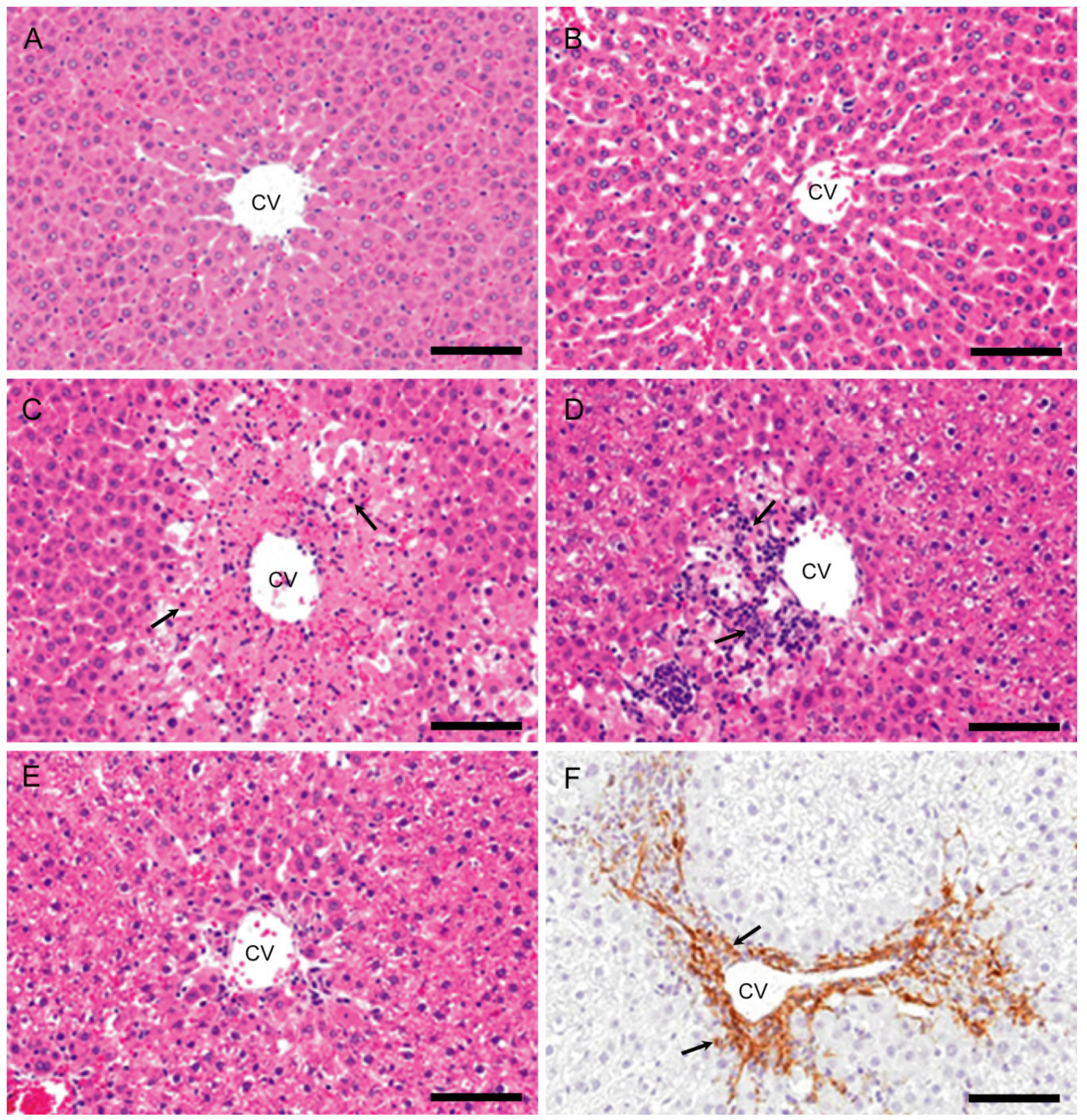
2.1. Gross and Histopathological Findings, and Serum Biochemistry
2.2. M1-/M2-Macrophages
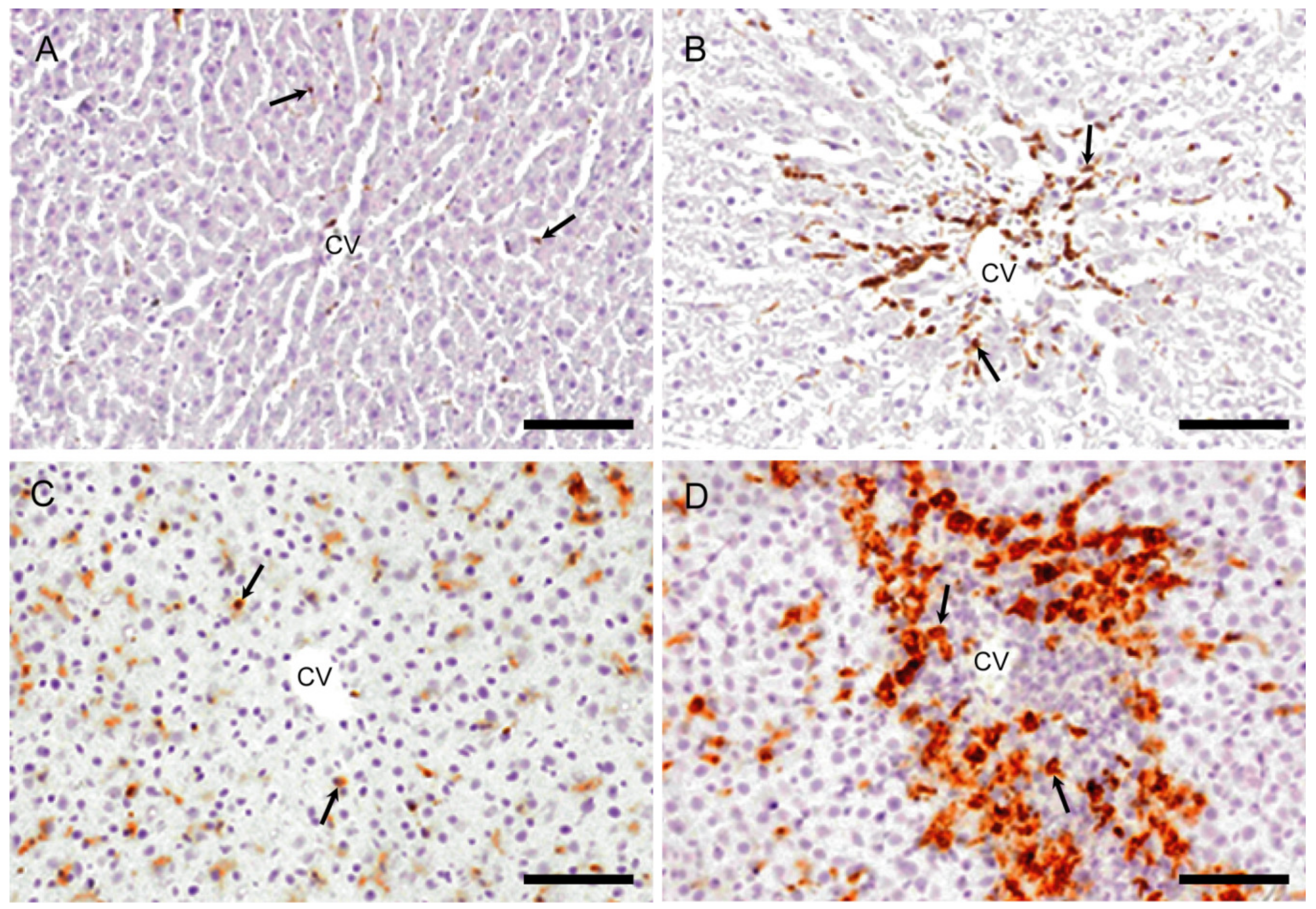
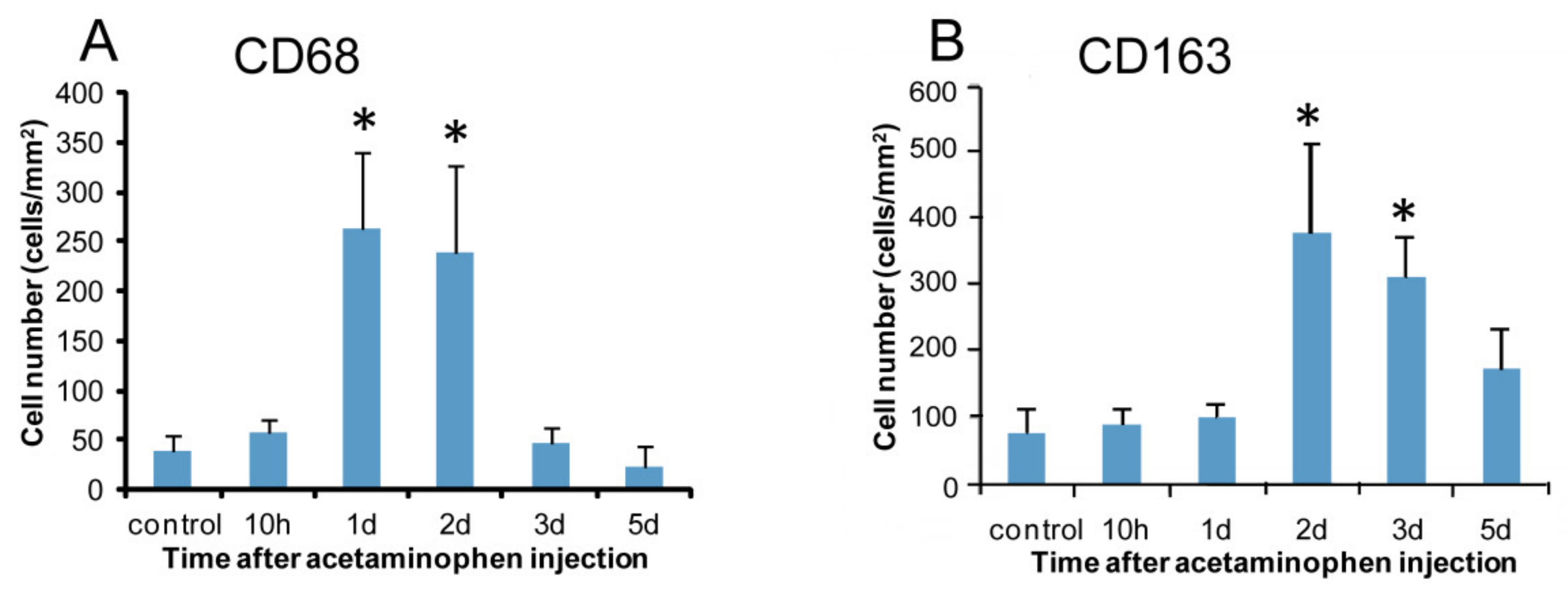
2.2.1. CD68 Immunostaining for M1-Macrophages
2.2.2. CD163 Immunostaining for M2-Macrophages
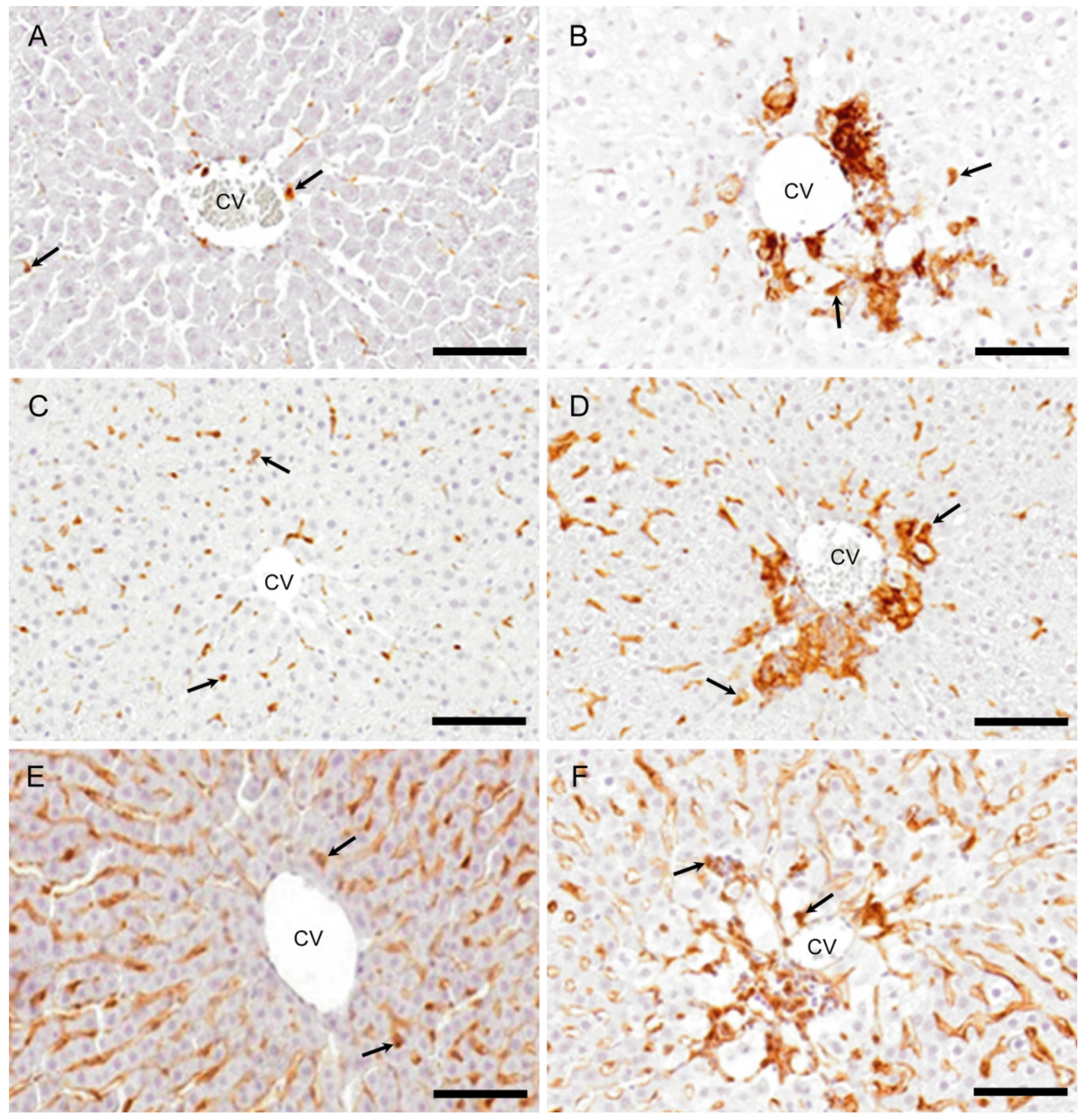
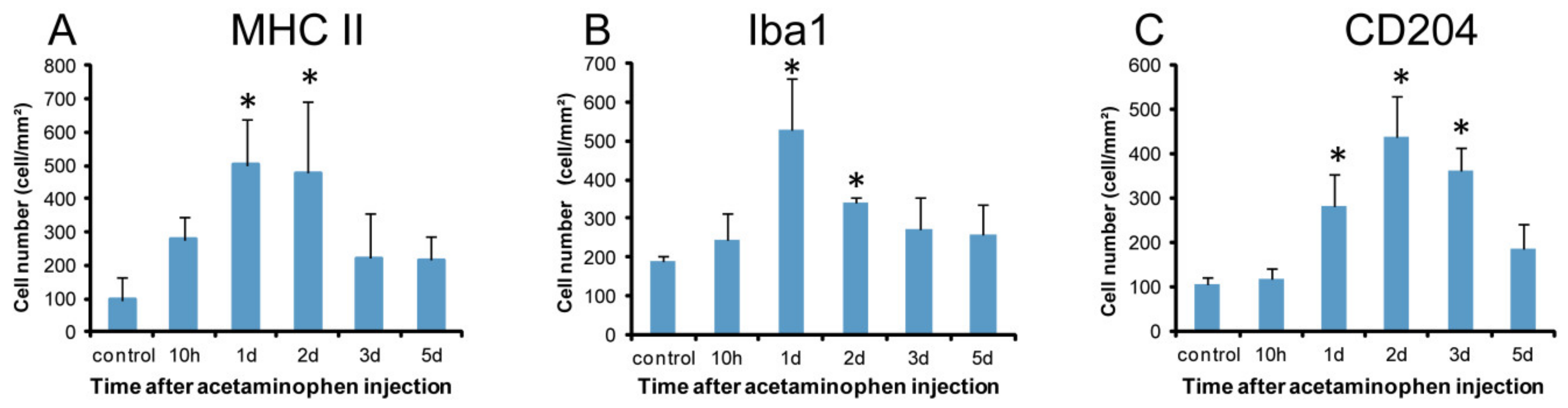
2.3. Macrophages with Immunophenotypes for MHC Class II, Iba1 and CD204
2.3.1. MHC Class II Immunostaining
2.3.2. Iba1 Immunostaining
2.3.3. CD204 Immunostaining
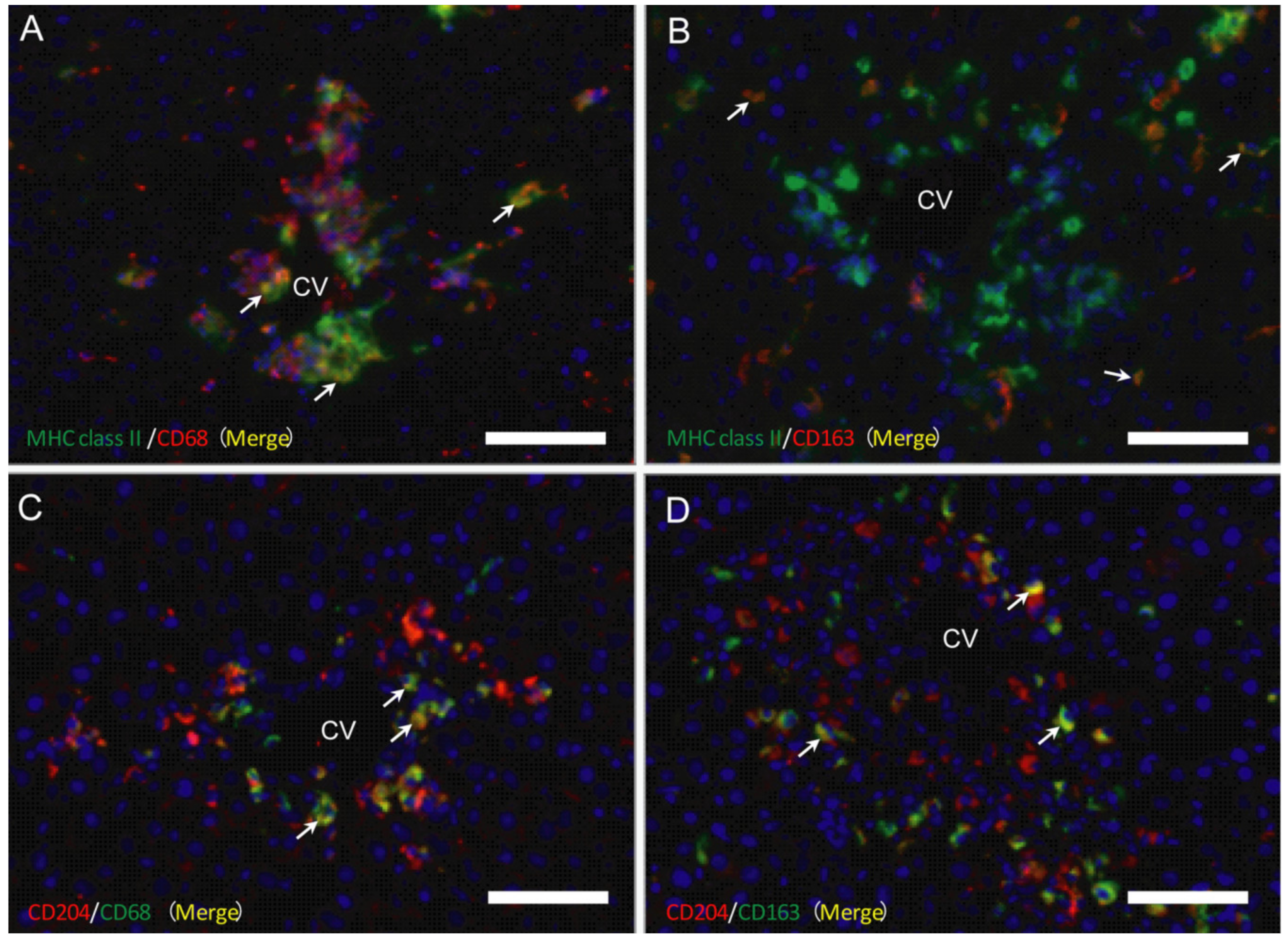
2.4. Double Immunofluorescence for Macrophage Markers
2.4.1. MHC Class II Positive Macrophages for M1-/M2-Polarization
2.4.2. CD204 Positive Macrophages for M1-/M2-Polarization
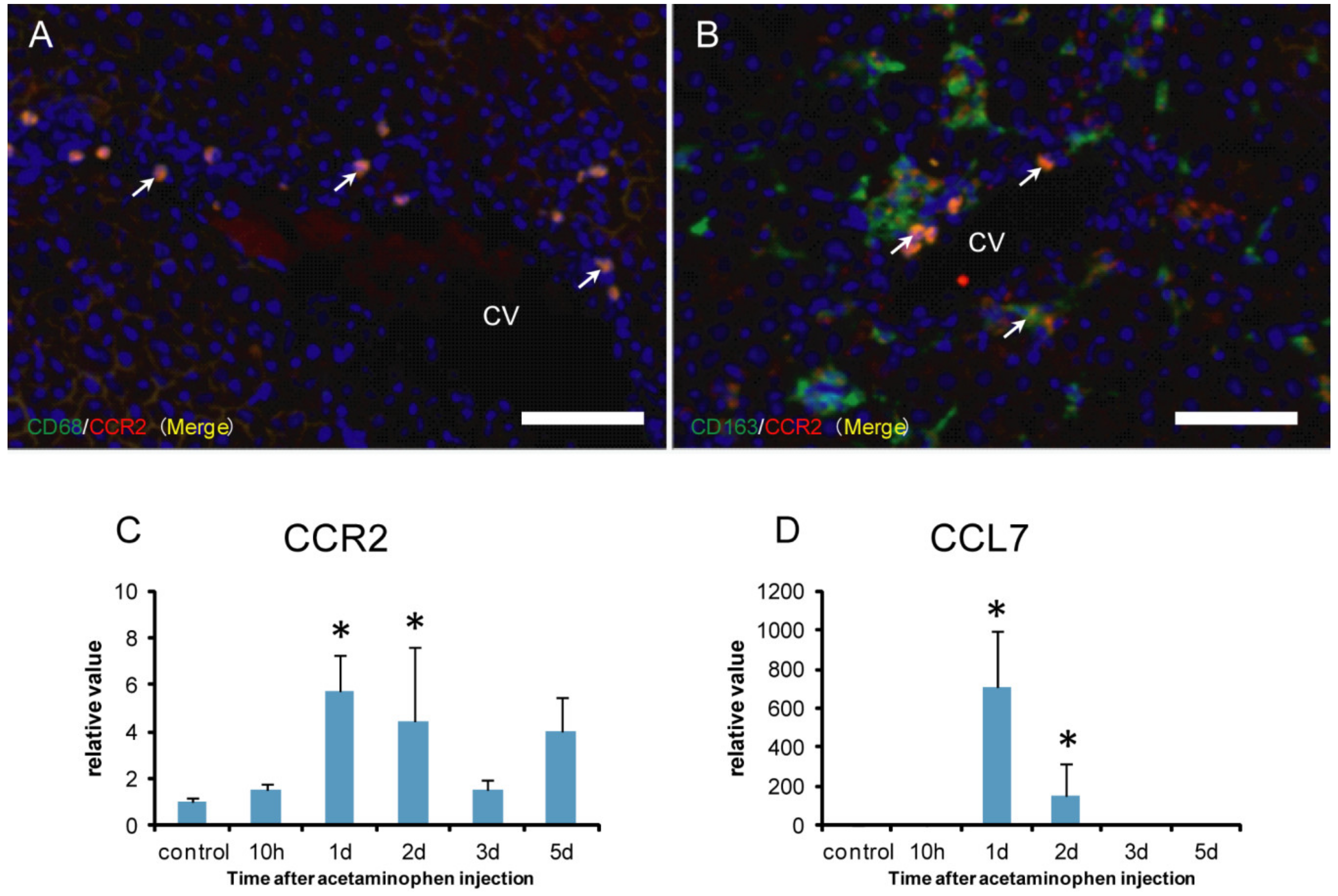
2.5. CCR2 Positive Macrophages for M1-/M2-Polarization
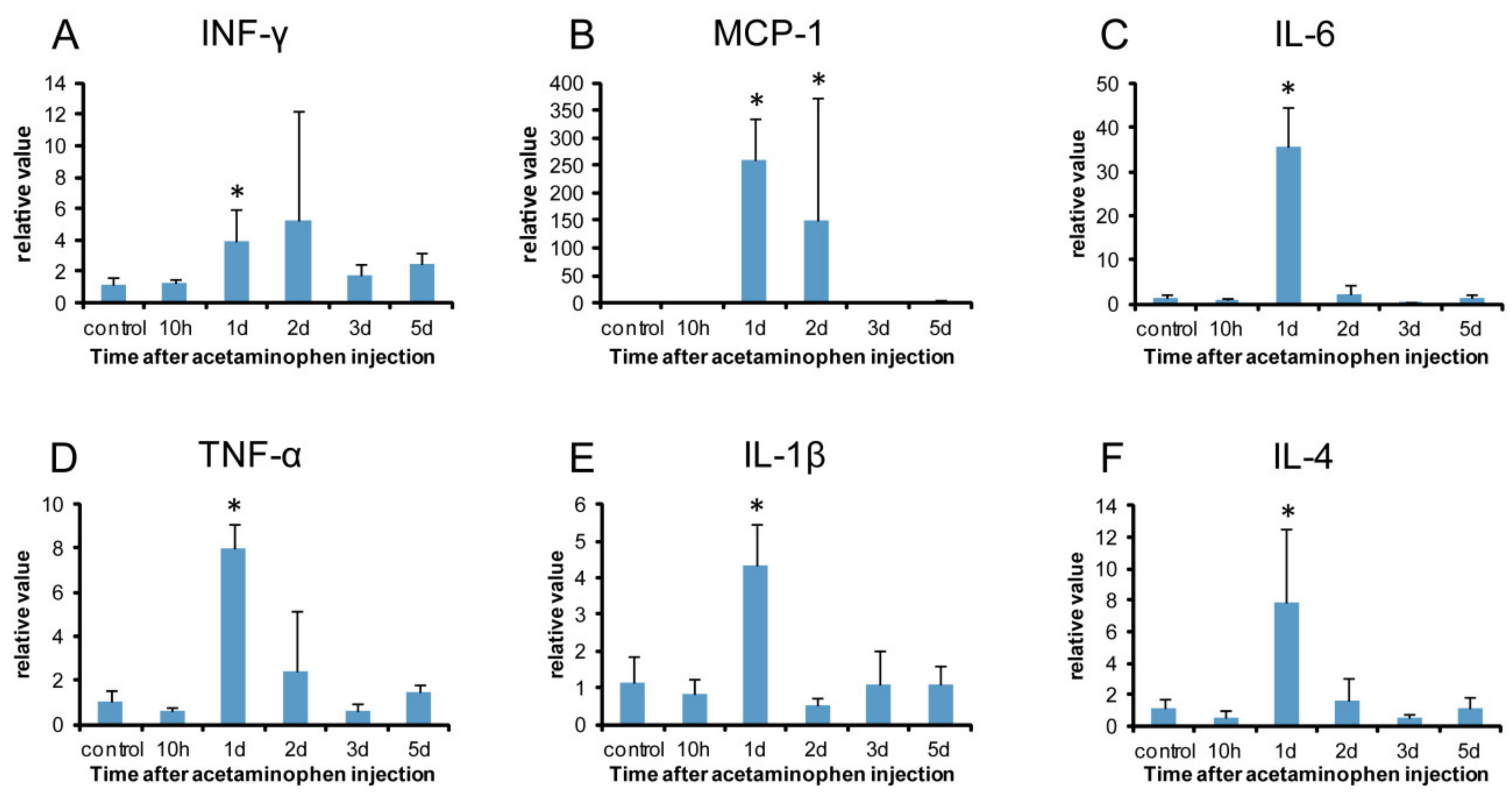
2.6. Analysis of M1- and M2-Macrophage-Related Factors
2.7. Analysis of DAMPs
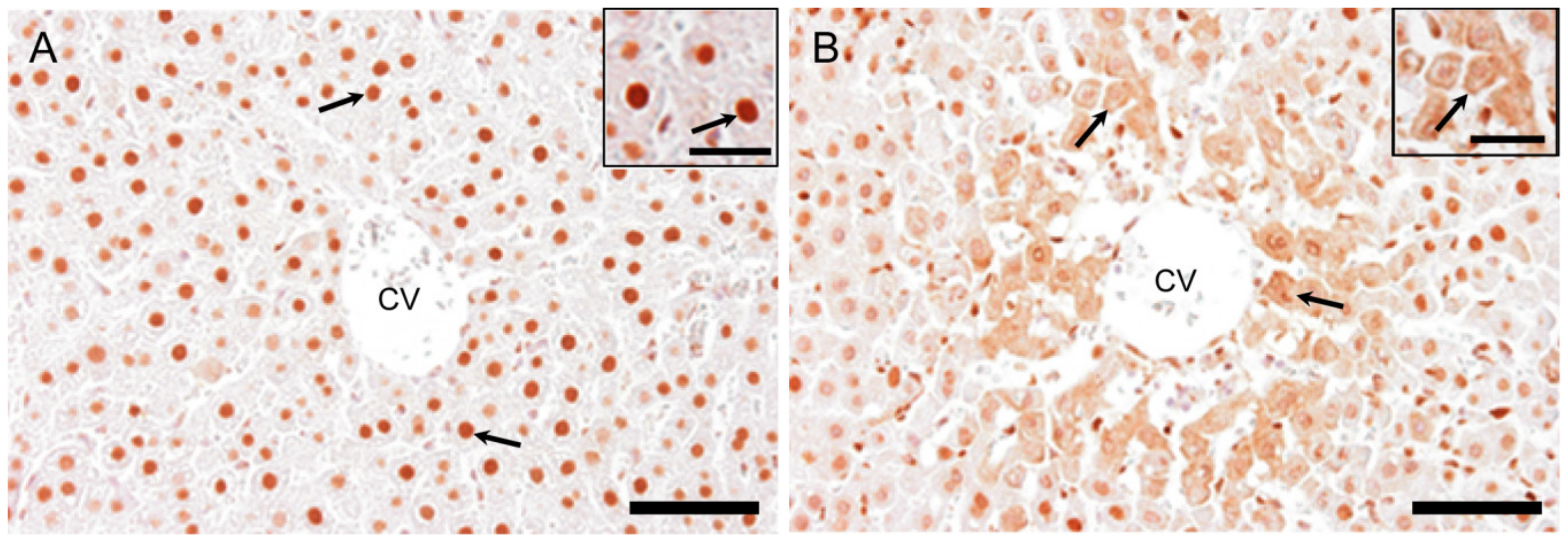
2.7.1. HMGB1 Immunostaining
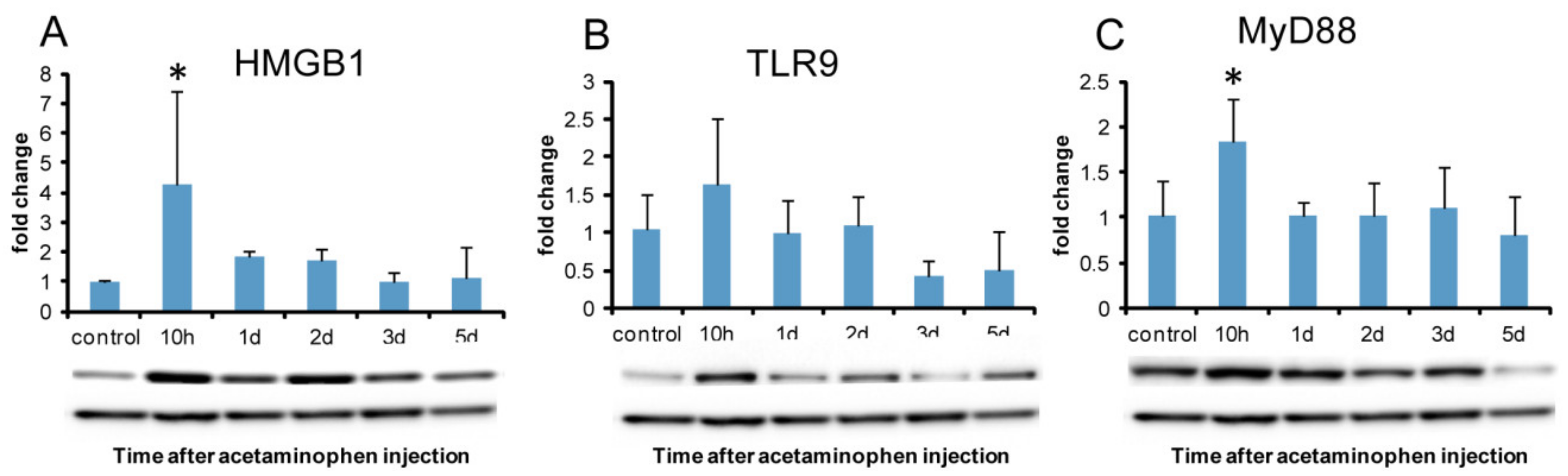
2.7.2. Western Blotting Analysis of HMGB1 and DAMPs Receptors
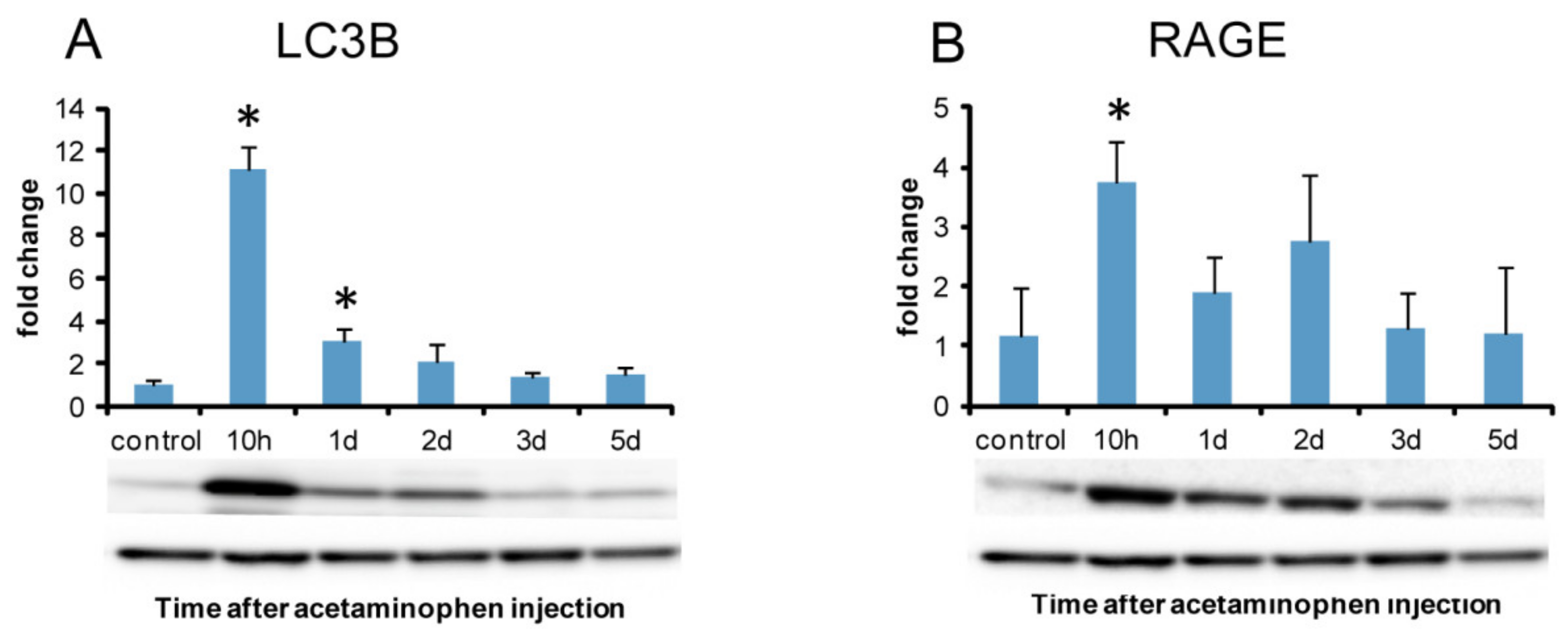
2.8. Analysis of Autophagy
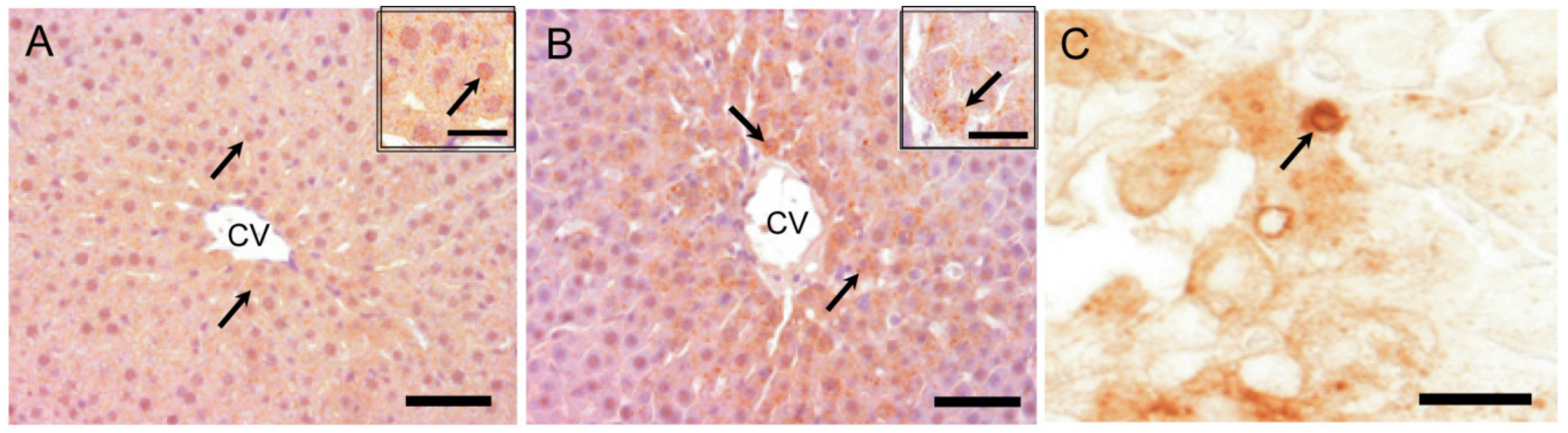
2.8.1. LC3B Immunostaining for Autophagosome Marker
2.8.2. Western Blotting Analysis of Autophagy Marker LC3B and Autophagy-Related Receptor (RAGE)
3. Discussion
3.1. M1-/M2-Macrophages
3.2. DAMPs
3.3. Autophagy
4. Materials and Methods
4.1. Animals
4.2. Serum Biochemistry
4.3. Histopathology and Immunohistochemistry
4.3.1. Histopathological Examination
4.3.2. Immunohistochemistry
4.3.3. Double Immunofluorescence
4.4. Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.5. Cell Counts
4.6. Western Blot
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Animal rights
Abbreviations
| T. Bil | Total bilirubin |
| ALT | Alanine transaminase |
| APAP | Acetaminophen |
| AST | Aspartate transaminase |
| CCL | Chemokine ligand |
| CCR | Chemokine receptor |
| CD | Cluster of differentiation |
| CV | Central vein |
| DAMP | Damage associated molecular pattern |
| ECMs | Extracellular matrices |
| HMGB | High mobility group box |
| Iba | Ionized calcium binding adaptor |
| IFN | Interferon |
| IL | Interleukin |
| MCP | Monocyte chemoattractant protein |
| MHC | Major histocompatibility complex |
| MyD | Myeloid Differentiation |
| PLP | Periodate-lysine-paraformaldehyde |
| RAGE | Receptor for Advanced Glycation End-products |
| RT-PCR | Reverse transcriptase polymerase chain reaction |
| SMA | Smooth muscle actin |
| TAA | Thioacetamide |
| TGF | Transforming growth factor |
| TLR | Toll-like receptor |
| TNF | Tumor necrosis factor |
References
- Maher, J.J. DAMPs ramp up drug toxicity. J. Clin. Invest. 2009, 119, 246–249. [Google Scholar] [CrossRef]
- McGill, M.R.; Williams, C.D.; Xie, Y.; Ramachandran, A.; Jaeschke, H. Acetaminophen-induced liver injury in rats and mice: Comparison of protein adducts, mitochondrial dysfunction, and oxidative stress in the mechanism of toxicity. Toxicol. Appl. Pharmacol. 2012, 264, 387–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGill, M.R.; Jaeschke, H. Metabolism and disposition of acetaminophen: Recent advances in relation to hepatotoxicity and diagnosis. Pharm. Res. 2013, 30, 2174–2187. [Google Scholar] [CrossRef] [Green Version]
- Ni, H.M.; McGill, M.R.; Chao, X.; Du, K.; Williams, J.A.; Xie, Y.; Jaeschke, H.; Ding, W.X. Removal of acetaminophen protein adducts by autophagy protects against acetaminophen-induced liver injury in mice. J. Hepatol. 2016, 65, 354–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolbright, B.L.; Jaeschke, H. Role of the inflammasome in acetaminophen-induced liver injury and acute liver failure. J. Hepatol. 2017, 66, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.Y.; Wu, L.Q.; Zhang, T.; Han, Y.F.; Lin, X. Autophagy-mediated HMGB1 release promotes gastric cancer cell survival via RAGE activation of extracellular signal-regulated kinases 1/2. Oncol. Rep. 2015, 33, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Melchior, W.B., Jr.; Wu, Y.; Guo, L. Autophagy in drug-induced liver toxicity. J. Food Drug Anal. 2014, 22, 161–168. [Google Scholar] [CrossRef]
- Dambach, D.M.; Watson, L.M.; Gray, K.R.; Durham, S.K.; Laskin, D.L. Role of CCR2 in macrophage migration into the liver during acetaminophen-induced hepatotoxicity in the mouse. Hepatology 2002, 35, 1093–1103. [Google Scholar] [CrossRef]
- Holt, M.P.; Cheng, L.; Ju, C. Identification and characterization of infiltrating macrophages in acetaminophen-induced liver injury. J. Leukoc. Biol. 2008, 84, 1410–1421. [Google Scholar] [CrossRef]
- Laskin, D.L.; Gardner, C.R.; Price, V.F.; Jollow, D.J. Modulation of macrophage functioning abrogates the acute hepatotoxicity of acetaminophen. Hepatology 1995, 21, 1045–1050. [Google Scholar] [CrossRef]
- Golbar, H.M.; Izawa, T.; Wijesundera, K.K.; Bondoc, A.; Tennakoon, A.H.; Kuwamura, M.; Yamate, J. Depletion of hepatic macrophages aggravates liver lesion induced in rats by thioacetamide (TAA). Toxicol. Pathol. 2016, 44, 246–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, D.M.; Justin, P.E. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 10, 958–969. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Invest. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Invernizzi, P.; Mantovani, A. Macrophage plasticity and polarization in liver homeostasis and pathology. Hepatology 2014, 59, 2034–2042. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 3, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesundera, K.K.; Izawa, T.; Tennakoon, A.H.; Golbar, H.M.; Tanaka, M.; Kuwamura, M.; Yamate, J. M1- and M2-Macrophage polarization in rat liver cirrhosis induced by thioacetamide ( TAA ), focusing on Iba1 and galectin-3. Exp. Mol. Pathol. 2014, 96, 382–392. [Google Scholar] [CrossRef]
- Yamate, J.; Izawa, T.; Kuwamura, M. Histopathological analysis of rat hepatotoxicity based on macrophage functions: In particular, an analysis for thioacetamide-induced hepatic lesions. Food Saf. 2016, 4, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Hori, O.; Brett, J.; Slattery, T.; Cao, R.; Zhang, J.; Chen, J.X.; Nagashima, M.; Lundh, E.R.; Vijay, S.; Nitecki, D.; et al. The Receptor for advanced glycation end products (RAGE) is a cellular binding site for amphoterin. J. Biol. Chem. 1995, 270, 25752–25761. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. TLR signaling. Semin. Immunol. 2007, 19, 24–32. [Google Scholar] [CrossRef]
- Ye, Z.; Ting, J.P.Y. NLR, the nucleotide-binding domain leucine-rich repeat containing gene family. Curr. Opin. Immunol. 2008, 20, 3–9. [Google Scholar] [CrossRef]
- Leung, C.S. Endogenous antigen presentation of MHC class II epitopes through non-autophagic pathways. Front. Immunol. 2015, 464, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: Signal 0s that spur autophagy and immunity. Immunol. Rev. 2012, 249, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. DAMPs, PAMPs and alarmins: All we need to know about danger. J. Leukoc. Biol. 2006, 81, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Jaeschke, H.; Williams, C.D.; Ramachandran, A.; Bajt, M.L. Acetaminophen hepatotoxicity and repair: The role of sterile inflammation and innate immunity. Liver Int. 2012, 32, 8–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsung, A.; Klune, J.R.; Zhang, X.; Jeyabalan, G.; Cao, Z.; Peng, X.; Stolz, D.B.; Geller, D.A.; Rosengart, M.R.; Billiar, T.R. HMGB1 release induced by liver ischemia involves Toll-like receptor 4–dependent reactive oxygen species production and calcium-mediated signaling. J. Exp. Med. 2007, 204, 2913–2923. [Google Scholar] [CrossRef] [PubMed]
- Kurahashi, T.; Lee, J.; Nabeshima, A.; Homma, T.; Kang, E.S.; Saito, Y.; Yamada, S.; Nakayama, T.; Yamada, K.; Miyata, S.; et al. Ascorbic acid prevents acetaminophen-induced hepatotoxicity in mice by ameliorating glutathione recovery and autophagy. Arch. Biochem. Biophys. 2016, 604, 36–46. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Livesey, K.M.; Cheh, C.W.; Farkas, A.; Loughran, P.; Hoppe, G.; Bianchi, M.E.; Tracey, K.J.; Zeh, H.J., 3rd; et al. Endogenous HMGB1 regulates autophagy. J. Cell Biol. 2010, 190, 881–892. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Cheh, C.W.; Livesey, K.M.; Liang, X.; Schapiro, N.E.; Benschop, R.; Sparvero, L.J.; Amoscato, A.A.; Tracey, K.J.; et al. HMGB1 release and redox regulates autophagy and apoptosis in cancer cells. Cancer 2011, 29, 5299–5310. [Google Scholar] [CrossRef] [Green Version]
- Ide, M.; Kuwamura, M.; Kotani, T.; Sawamoto, O.; Yamate, J. Effects of gadolinium chloride (GdCl3) on the appearance of macrophage populations and fibrogenesis in the thioacetamide induced rat hepatic lesions. J. Comp. Pathol. 2005, 133, 92–102. [Google Scholar] [CrossRef]
- Polfliet, M.M.; Fabriek, B.O.; Daniëls, W.P.; Dijkstra, C.D.; van den Berg, T.K. The rat macrophage scavenger receptor CD163: Expression, regulation and role in inflammatory mediator production. Immunobiology 2006, 211, 419–425. [Google Scholar] [CrossRef]
- Bajpai, G.; Schneider, C.; Wong, N.; Bredemeyer, A.; Hulsmans, M.; Nahrendorf, M.; Epelman, S.; Kreisel, D.; Liu, Y.; Itoh, A.; et al. The human heart contains distinct macrophage subsets with divergent origins and functions. Nat. Med. 2018, 4, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F. Review targeting hepatic macrophages to treat liver diseases. J. Hepatol. 2017, 66, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Ilavenil, S.; Al-Dhabi, N.A.; Srigopalram, A.; Kim, Y.O.; Agastian, P.; Baru, R.; Choi, K.C.; Arasu, M.V. Acetaminophen induced hepatotoxicity in wistar rats--a proteomic approach. Mol. 2016, 21, 161. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y. M1 and M2 polarization of macrophages: A mini-review. Med Biol Sci Eng. 2019, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Golbar, H.M.; Izawa, T.; Murai, F.; Kuwamura, M.; Yamate, J. Immunohistochemical analyses of the kinetics and distribution of macrophages, hepatic stellate cells and bile duct epithelia in the developing rat liver. Exp. Toxicol. Pathol. 2012, 64, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Kelemen, S.E.; Autieri, M.V. Inhibition of AIF-1 expression by constitutive SiRNA expression reduces macrophage migration, proliferation, and signal transduction initiated by atherogenic stimuli. Am. J. Physiol. Cell Physiol. 2006, 290, C1083–C1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomokiyo, R.; Jinnouchi, K.; Honda, M.; Wada, Y.; Hanada, N.; Hiraoka, T.; Suzuki, H.; Kodama, T.; Takahashi, K.; Takeya, M. Production, characterization, and interspecies reactivities of monoclonal antibodies against human class A macrophage scavenger receptors. Atherosclerosis 2002, 161, 123–132. [Google Scholar] [CrossRef]
- Ricardo, S.D.; van Goor, H.; Eddy, A.A. Macrophage diversity in renal injury and repair. J. Clin. Invest. 2008, 118, 3522–3530. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.M.; Liu, K.; Liu, J.H.; Jiang, X.L.; Wang, X.L.; Chen, Y.Z.; LI, S.G.; Zou, H.; Pang, L.J.; Liu, C.X.; et al. CD163 as a marker of M2 macrophage, contribute to predict aggressiveness and prognosis of Kazakh esophageal squamous cell carcinoma. Oncotarget 2017, 8, 21526–21538. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Harris, D.C.V.H. Macrophages in renal disease. J. Am. Soc. Nephrol. 2011, 22, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Yamate, J.; Sato, K.; Ide, M.; Nakanishi, M.; Kuwamura, M.; Sakuma, S.; Nakatsuji, S. Participation of different macrophage populations and myofibroblastic cells in chronically developed renal interstitial fibrosis after cisplatin-induced renal injury in rats. Vet. Pathol. 2002, 39, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Yamate, J.; Machida, Y.; Ide, M.; Kuwamura, M.; Kotani, T.; Sawamoto, O.; LaMarre, J. Cisplatin-induced renal interstitial fibrosis in neonatal rats, developing as solitary nephron unit lesions. Toxicol. Pathol. 2005, 33, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.H.; Sanders, C.; Johns, E.W. A new group of chromatin-associated proteins with a high content of acidic and basic amino acids. Eur. J. Biochem. 2000, 19, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Guan, X.; Zuo, X.; Wang, J.; Yin, W. The role of high mobility group box 1 (HMGB1) in the pathogenesis of kidney diseases. Acta Pharm. Sin. B. 2016, 6, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Chen, X. Expression of High-Mobility Group Box 1 protein (HMGB1) and Toll-Like Receptor 9 (TLR9) in retinas of diabetic rats. Med. Sci. Monit. 2017, 23, 3115–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudaliar, H.; Pollock, C.; Ma, J.; Wu, H.; Chadban, S.; Panchapakesan, U. The Role of TLR2 and 4-mediated inflammatory pathways in endothelial cells exposed to high glucose. Plos One 2014, 9, 1–14. [Google Scholar] [CrossRef]
- Tsung, A.; Tohme, S.; Billiar, T.R. High-Mobility Group Box-1 in sterile inflammation. J. Intern. Med. 2014, 276, 425–443. [Google Scholar] [CrossRef]
- Dinakaran, D.; Bristow, E.; Armanious, H.; Garros, D.; Yap, J.; Noga, M.; Sergi, C. Co-ingestion of willow bark tea and acetaminophen associated with fatal infantile fulminant liver failure. Pediatr. Int. 2017, 59, 743–745. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Zhang, J.; Dai, C.; Liu, X.; Wang, J.; Gao, Z.; Guo, H.; Wang, R.; Lu, S.; et al. S100A4 promotes liver fibrosis via activation of hepatic stellate cells. J. Hepatol. 2015, 62, 156–164. [Google Scholar] [CrossRef]
- Karvellas, C.J.; Speiser, J.L.; Tremblay, M.; Lee, W.M.; Rose, C.F.; US Acute Liver Failure Study Group. Elevated FABP1 serum levels are associated with poorer survival in acetaminophen-induced acute liver failure. Hepatology 2017, 65, 938–949. [Google Scholar] [CrossRef]
- Mariño, G.; Madeo, F.; Kroemer, G. Autophagy for tissue homeostasis and neuroprotection. Curr. Opin. Cell Biol. 2011, 23, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Lu, J. Autophagy and macrophage functions: Inflammatory response and phagocysstosis. Cells 2020, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.; Zhao, E.; Ilyas, G.; Lalazar, G.; Lin, Y.; Haseeb, M.; Tanaka, K.E.; Czaja, M.J. Impaired macrophage autophagy increases the immune response in obese mice by promoting proinflammatory macrophage polarization. Autophagy 2015, 11, 271–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, J.; El-benna, J. Macrophage autophagy protects against liver fibrosis in mice. Autophagy 2015, 11, 1280–1292. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| control | 10 h | 1 d | 2 d | 3 d | 5 d |
|---|---|---|---|---|---|
| ± | + | ++ | ± | ± | ± |
| Antibody (Clone) | Host | Fixative | Dilution | Pretreatment | Source |
|---|---|---|---|---|---|
| CD68 (ED-1) | Mouse Monoclonal | PLP solution | 1/500 | 10 µg/mL proteinase K, RT, 10 min | Millipore, Massachusetts, USA |
| CD163 (ED-2) | Mouse Monoclonal | Acetone/methanol (1:1) | 1/300 | ― | AbD Serotec, Oxford, UK |
| MHC class II (OX-6) | Mouse Monoclonal | PLP solution | 1/300 | Microwave in Citrate buffer, 20 min | AbD Serotec, Oxford, UK |
| CD204 (SRA-E5) | Mouse Monoclonal | PLP solution | 1/500 | Microwave in Citrate buffer, 20 min | Transgenic Inc, Kumamoto, Japan |
| Iba1 | Rabbit Polyclonal | Formalin | 1/500 | Microwave in Citrate buffer, 20 min | Wako, Osaka, Japan |
| α-SMA (1A4) | Mouse Monoclonal | Formalin | 1/1,000 | ― | Dako, Glostrup, Denmark |
| HMGB1 | Rabbit Polyclonal | PLP solution | 1/200 | Microwave in Citrate buffer, 20 min | Abcam, Cambridge, UK |
| LC3B | Rabbit Polyclonal | PLP solution | 1/1,000 | Microwave in Citrate buffer, 20 min | SIGMA, St. Louis, USA |
| Probe | Assay ID | Probe | Assay ID |
|---|---|---|---|
| IL-6 | Rn01410330_m1 | IL-10 | Rn00563409_m1 |
| IFN-γ | Rn00594078_m1 | TGF-β1 | Rn00572010_m1 |
| MCP-1 | Rn00580555_m1 | CCR2 | Rn01637698_s1 |
| TNF-α | Rn01525859_g1 | CCL7 | Rn01467286_m1 |
| IL-1β | Rn00580432_ml | Ribosomal 18s | Hs99999901_s1 |
| IL-4 | Rn01456866_m1 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuji, Y.; Kuramochi, M.; Golbar, H.M.; Izawa, T.; Kuwamura, M.; Yamate, J. Acetaminophen-Induced Rat Hepatotoxicity Based on M1/M2-Macrophage Polarization, in Possible Relation to Damage-Associated Molecular Patterns and Autophagy. Int. J. Mol. Sci. 2020, 21, 8998. https://doi.org/10.3390/ijms21238998
Tsuji Y, Kuramochi M, Golbar HM, Izawa T, Kuwamura M, Yamate J. Acetaminophen-Induced Rat Hepatotoxicity Based on M1/M2-Macrophage Polarization, in Possible Relation to Damage-Associated Molecular Patterns and Autophagy. International Journal of Molecular Sciences. 2020; 21(23):8998. https://doi.org/10.3390/ijms21238998
Chicago/Turabian StyleTsuji, Yuka, Mizuki Kuramochi, Hossain M. Golbar, Takeshi Izawa, Mitsuru Kuwamura, and Jyoji Yamate. 2020. "Acetaminophen-Induced Rat Hepatotoxicity Based on M1/M2-Macrophage Polarization, in Possible Relation to Damage-Associated Molecular Patterns and Autophagy" International Journal of Molecular Sciences 21, no. 23: 8998. https://doi.org/10.3390/ijms21238998
APA StyleTsuji, Y., Kuramochi, M., Golbar, H. M., Izawa, T., Kuwamura, M., & Yamate, J. (2020). Acetaminophen-Induced Rat Hepatotoxicity Based on M1/M2-Macrophage Polarization, in Possible Relation to Damage-Associated Molecular Patterns and Autophagy. International Journal of Molecular Sciences, 21(23), 8998. https://doi.org/10.3390/ijms21238998