Nitrogen Starvation and Stationary Phase Lipophagy Have Distinct Molecular Mechanisms



Abstract
:1. Introduction
2. Results
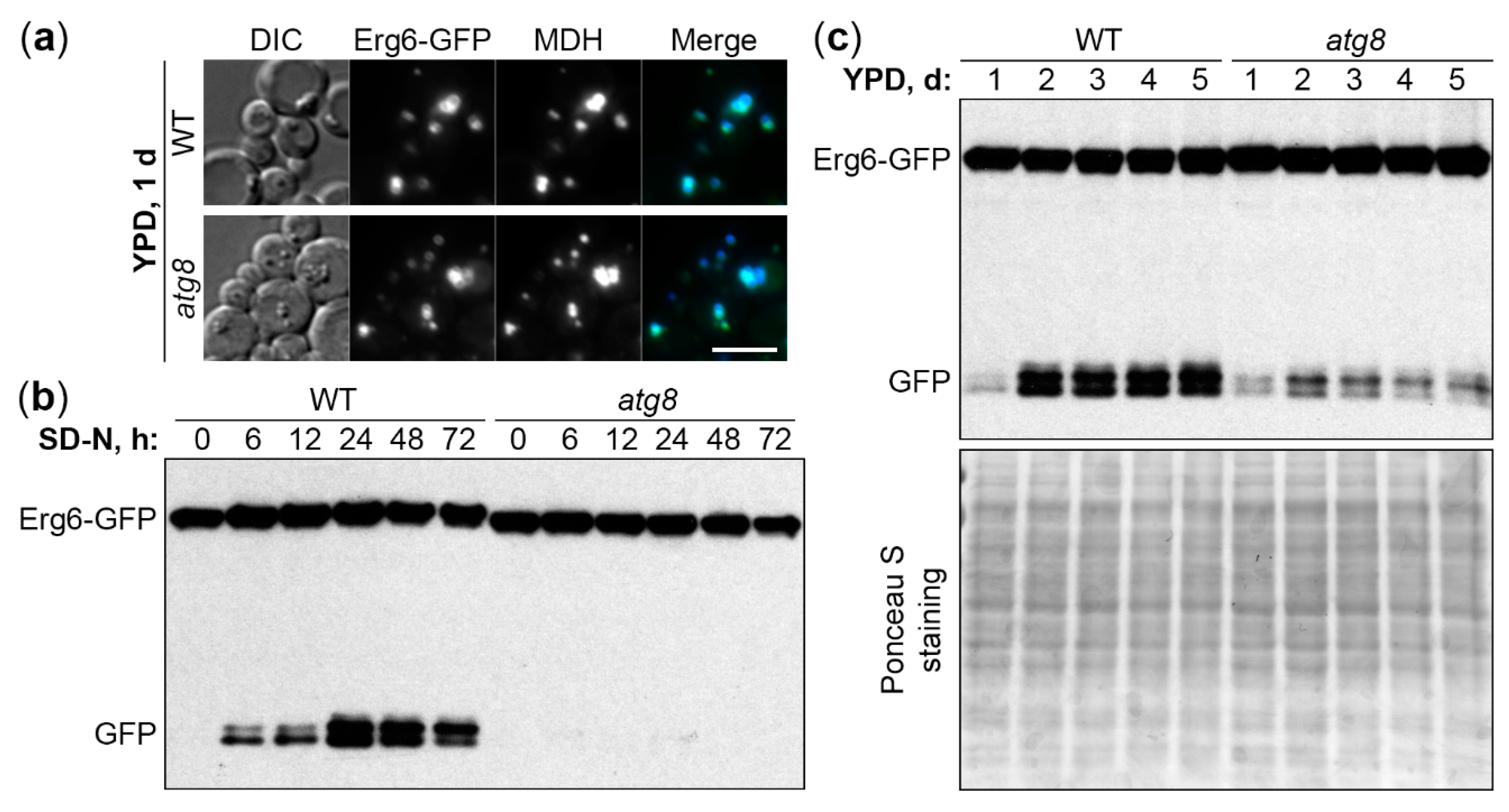
2.1. K. phaffii Is a Good Model for Both N-Starvation and S-Phase Lipophagy
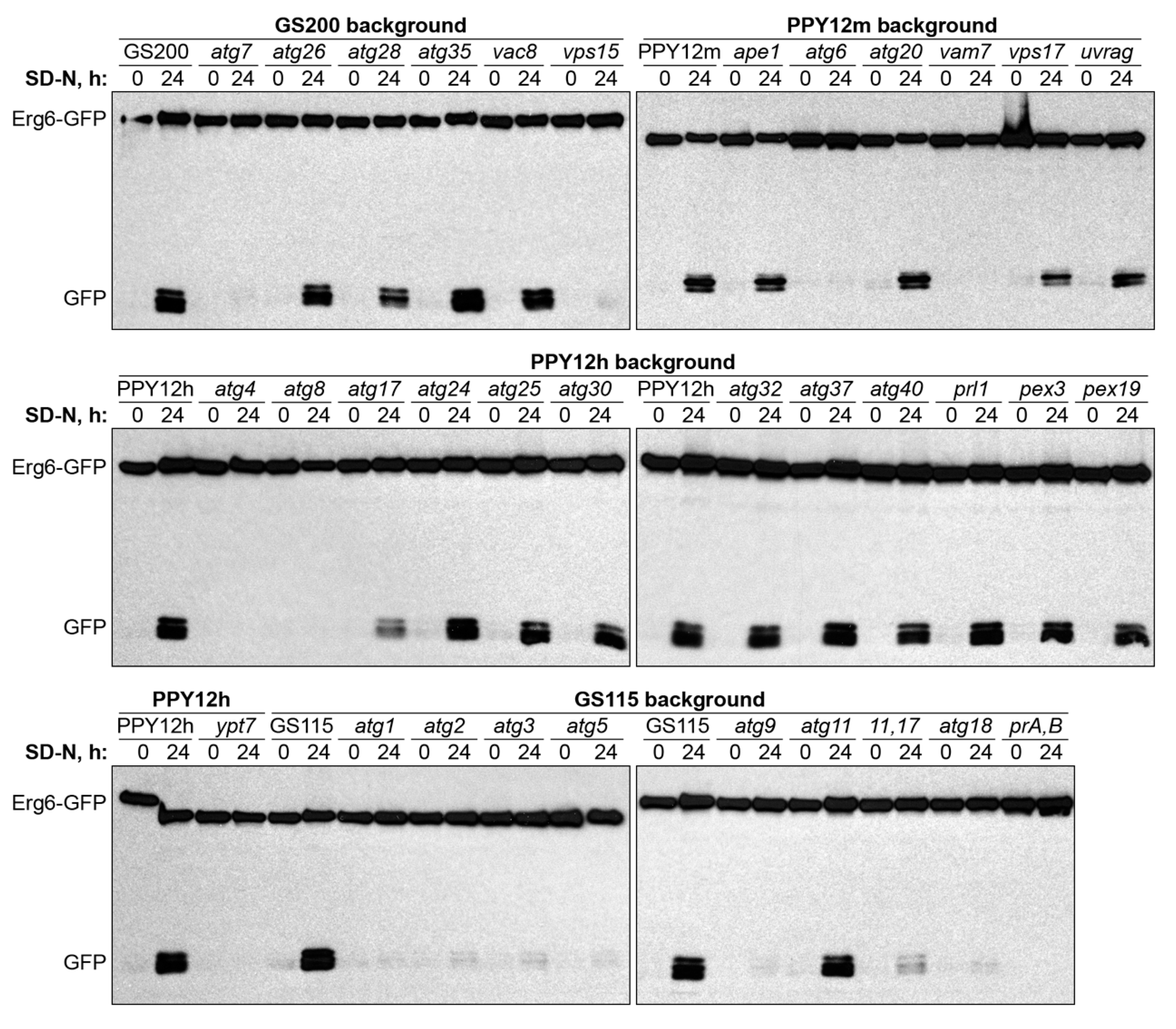
2.2. Molecular Requirements of N-Starvation and S-Phase Lipophagy in K. phaffii
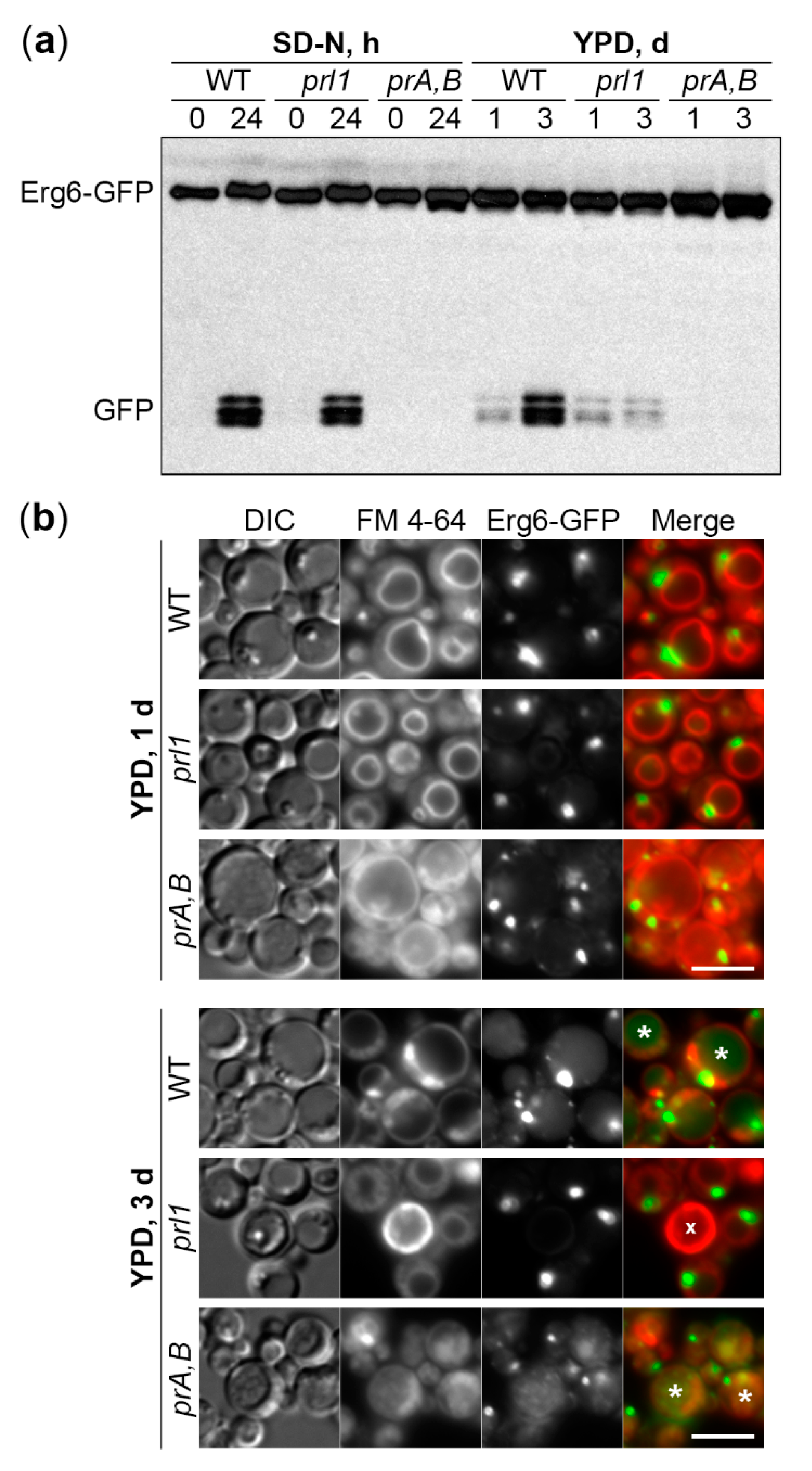
2.3. Prl1 Is Essential for the Delivery of LDs to the Vacuole in S-Phase
3. Discussion
4. Materials and Methods
4.1. Strains and Plasmids
4.2. Fluorescence Microscopy
4.3. Biochemical Studies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Atg | autophagy-related |
| C | carbon |
| DIC | differential interference contrast |
| ESCRT | endosomal sorting complex required for transport |
| GFP | green fluorescent protein |
| LD | lipid droplet |
| MDH | monodansylpentane |
| N | nitrogen |
| Prl1 | positive regulator of lipophagy 1 |
| prA,B | proteinases A and B |
| S | stationary |
| WT | wild-type |
Appendix A


References
- Klionsky, D.J. Autophagy: From phenomenology to molecular understanding in less than a decade. Nat. Rev. Mol. Cell Biol. 2007, 8, 931–937. [Google Scholar] [CrossRef]
- Ohsumi, Y. Historical landmarks of autophagy research. Cell Res. 2014, 24, 9–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshige, K.; Baba, M.; Tsuboi, S.; Noda, T.; Ohsumi, Y. Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. J. Cell Biol. 1992, 119, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Sutter, B.M.; Wu, X.; Laxman, S.; Tu, B.P. Methionine inhibits autophagy and promotes growth by inducing the SAM-responsive methylation of PP2A. Cell 2013, 154, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Marchesini, G.; Brizi, M.; Bianchi, G.; Tomassetti, S.; Bugianesi, E.; Lenzi, M.; McCullough, A.J.; Natale, S.; Forlani, G.; Melchionda, N. Nonalcoholic fatty liver disease: A feature of the metabolic syndrome. Diabetes 2001, 50, 1844–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Cuervo, A.M. Autophagy in the cellular energetic balance. Cell Metab. 2011, 13, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yao, Z.; Chen, Y.; Qian, L.; Jiang, S.; Zhou, J.; Shao, J.; Chen, A.; Zhang, F.; Zheng, S. Lipophagy and liver disease: New perspectives to better understanding and therapy. Biomed. Pharmacother. 2018, 97, 339–348. [Google Scholar] [CrossRef]
- Zhou, K.; Yao, P.; He, J.; Zhao, H. Lipophagy in nonliver tissues and some related diseases: Pathogenic and therapeutic implications. J. Cell. Physiol. 2019, 234, 7938–7947. [Google Scholar] [CrossRef]
- Van Zutphen, T.; Todde, V.; de Boer, R.; Kreim, M.; Hofbauer, H.F.; Wolinski, H.; Veenhuis, M.; van der Klei, I.J.; Kohlwein, S.D. Lipid droplet autophagy in the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2014, 25, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, T.; Fujimoto, M.; Tatematsu, T.; Cheng, J.; Orii, M.; Takatori, S.; Fujimoto, T. Niemann-Pick type C proteins promote microautophagy by expanding raft-like membrane domains in the yeast vacuole. Elife 2017, 6. [Google Scholar] [CrossRef]
- Seo, A.Y.; Lau, P.W.; Feliciano, D.; Sengupta, P.; Gros, M.A.L.; Cinquin, B.; Larabell, C.A.; Lippincott-Schwartz, J. AMPK and vacuole-associated Atg14p orchestrate µ-lipophagy for energy production and long-term survival under glucose starvation. Elife 2017, 6. [Google Scholar] [CrossRef]
- Wang, C.W.; Miao, Y.H.; Chang, Y.S. A sterol-enriched vacuolar microdomain mediates stationary phase lipophagy in budding yeast. J. Cell Biol. 2014, 206, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Farre, J.C.; Subramani, S. Mechanistic insights into selective autophagy pathways: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 2016, 17, 537–552. [Google Scholar] [CrossRef]
- Yang, H.J.; Hsu, C.L.; Yang, J.Y.; Yang, W.Y. Monodansylpentane as a blue-fluorescent lipid-droplet marker for multi-color live-cell imaging. PLoS ONE 2012, 7, e32693. [Google Scholar] [CrossRef] [Green Version]
- Shintani, T.; Klionsky, D.J. Cargo proteins facilitate the formation of transport vesicles in the cytoplasm to vacuole targeting pathway. J. Biol. Chem. 2004, 279, 29889–29894. [Google Scholar] [CrossRef] [Green Version]
- Cregg, J.M.; Barringer, K.J.; Hessler, A.Y.; Madden, K.R. Pichia pastoris as a host system for transformations. Mol. Cell. Biol. 1985, 5, 3376–3385. [Google Scholar] [CrossRef]
- Waterham, H.R.; de Vries, Y.; Russel, K.A.; Xie, W.; Veenhuis, M.; Cregg, J.M. The Pichia pastoris PER6 gene product is a peroxisomal integral membrane protein essential for peroxisome biogenesis and has sequence similarity to the Zellweger syndrome protein PAF-1. Mol. Cell. Biol. 1996, 16, 2527–2536. [Google Scholar] [CrossRef] [Green Version]
- Gould, S.J.; McCollum, D.; Spong, A.P.; Heyman, J.A.; Subramani, S. Development of the yeast Pichia pastoris as a model organism for a genetic and molecular analysis of peroxisome assembly. Yeast 1992, 8, 613–628. [Google Scholar] [CrossRef]
- Farre, J.C.; Vidal, J.; Subramani, S. A cytoplasm to vacuole targeting pathway in P. pastoris. Autophagy 2007, 3, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Stromhaug, P.E.; Bevan, A.; Dunn, W.A., Jr. GSA11 encodes a unique 208-kDa protein required for pexophagy and autophagy in Pichia pastoris. J. Biol. Chem. 2001, 276, 42422–42435. [Google Scholar] [CrossRef] [Green Version]
- Mukaiyama, H.; Oku, M.; Baba, M.; Samizo, T.; Hammond, A.T.; Glick, B.S.; Kato, N.; Sakai, Y. Paz2 and 13 other PAZ gene products regulate vacuolar engulfment of peroxisomes during micropexophagy. Genes Cells 2002, 7, 75–90. [Google Scholar] [CrossRef]
- Farre, J.C.; Mathewson, R.D.; Manjithaya, R.; Subramani, S. Roles of Pichia pastoris Uvrag in vacuolar protein sorting and the phosphatidylinositol 3-kinase complex in phagophore elongation in autophagy pathways. Autophagy 2010, 6, 86–99. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Stromhaug, P.E.; Dunn, W.A., Jr. Glucose-induced autophagy of peroxisomes in Pichia pastoris requires a unique E1-like protein. Mol. Biol. Cell 1999, 10, 1353–1366. [Google Scholar] [CrossRef] [Green Version]
- Nazarko, T.Y.; Farre, J.C.; Subramani, S. Peroxisome size provides insights into the function of autophagy-related proteins. Mol. Biol. Cell 2009, 20, 3828–3839. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kamada, Y.; Stromhaug, P.E.; Guan, J.; Hefner-Gravink, A.; Baba, M.; Scott, S.V.; Ohsumi, Y.; Dunn, W.A., Jr.; Klionsky, D.J. Cvt9/Gsa9 functions in sequestering selective cytosolic cargo destined for the vacuole. J. Cell. Biol. 2001, 153, 381–396. [Google Scholar] [CrossRef] [Green Version]
- Ano, Y.; Hattori, T.; Oku, M.; Mukaiyama, H.; Baba, M.; Ohsumi, Y.; Kato, N.; Sakai, Y. A sorting nexin PpAtg24 regulates vacuolar membrane dynamics during pexophagy via binding to phosphatidylinositol-3-phosphate. Mol. Biol. Cell 2005, 16, 446–457. [Google Scholar] [CrossRef]
- Stasyk, O.V.; Nazarko, T.Y.; Stasyk, O.G.; Krasovska, O.S.; Warnecke, D.; Nicaud, J.M.; Cregg, J.M.; Sibirny, A.A. Sterol glucosyltransferases have different functional roles in Pichia pastoris and Yarrowia lipolytica. Cell Biol. Int. 2003, 27, 947–952. [Google Scholar] [CrossRef]
- Stasyk, O.V.; Stasyk, O.G.; Mathewson, R.D.; Farre, J.C.; Nazarko, V.Y.; Krasovska, O.S.; Subramani, S.; Cregg, J.M.; Sibirny, A.A. Atg28, a novel coiled-coil protein involved in autophagic degradation of peroxisomes in the methylotrophic yeast Pichia pastoris. Autophagy 2006, 2, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Farre, J.C.; Burkenroad, A.; Burnett, S.F.; Subramani, S. Phosphorylation of mitophagy and pexophagy receptors coordinates their interaction with Atg8 and Atg11. EMBO Rep. 2013, 14, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Nazarko, V.Y.; Nazarko, T.Y.; Farre, J.C.; Stasyk, O.V.; Warnecke, D.; Ulaszewski, S.; Cregg, J.M.; Sibirny, A.A.; Subramani, S. Atg35, a micropexophagy-specific protein that regulates micropexophagic apparatus formation in Pichia pastoris. Autophagy 2011, 7, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Nazarko, T.Y.; Ozeki, K.; Till, A.; Ramakrishnan, G.; Lotfi, P.; Yan, M.; Subramani, S. Peroxisomal Atg37 binds Atg30 or palmitoyl-CoA to regulate phagophore formation during pexophagy. J. Cell. Biol. 2014, 204, 541–557. [Google Scholar] [CrossRef] [Green Version]
- Tuttle, D.L.; Dunn, W.A., Jr. Divergent modes of autophagy in the methylotrophic yeast Pichia pastoris. J. Cell Sci. 1995, 108 (Pt 1), 25–35. [Google Scholar]
- Wiemer, E.A.; Luers, G.H.; Faber, K.N.; Wenzel, T.; Veenhuis, M.; Subramani, S. Isolation and characterization of Pas2p, a peroxisomal membrane protein essential for peroxisome biogenesis in the methylotrophic yeast Pichia pastoris. J. Biol. Chem. 1996, 271, 18973–18980. [Google Scholar] [CrossRef] [Green Version]
- Snyder, W.B.; Faber, K.N.; Wenzel, T.J.; Koller, A.; Luers, G.H.; Rangell, L.; Keller, G.A.; Subramani, S. Pex19p interacts with Pex3p and Pex10p and is essential for peroxisome biogenesis in Pichia pastoris. Mol. Biol. Cell 1999, 10, 1745–1761. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.; Schroder, L.A.; Thomson, J.M.; Klocman, A.S.; Tomasini, A.J.; Stromhaug, P.E.; Dunn, W.A., Jr. PpATG9 encodes a novel membrane protein that traffics to vacuolar membranes, which sequester peroxisomes during pexophagy in Pichia pastoris. Mol. Biol. Cell 2005, 16, 4941–4953. [Google Scholar] [CrossRef] [Green Version]
- Manjithaya, R.; Anjard, C.; Loomis, W.F.; Subramani, S. Unconventional secretion of Pichia pastoris Acb1 is dependent on GRASP protein, peroxisomal functions, and autophagosome formation. J. Cell. Biol. 2010, 188, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Stasyk, O.V.; van der Klei, I.J.; Bellu, A.R.; Shen, S.; Kiel, J.A.; Cregg, J.M.; Veenhuis, M. A Pichia pastoris VPS15 homologue is required in selective peroxisome autophagy. Curr. Genet. 1999, 36, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Obara, K.; Ohsumi, Y. PtdIns 3-kinase orchestrates autophagosome formation in yeast. J. Lipids 2011, 2011, 498768. [Google Scholar] [CrossRef] [Green Version]
- Toulmay, A.; Prinz, W.A. Direct imaging reveals stable, micrometer-scale lipid domains that segregate proteins in live cells. J. Cell. Biol. 2013, 202, 35–44. [Google Scholar] [CrossRef]
- Oku, M.; Maeda, Y.; Kagohashi, Y.; Kondo, T.; Yamada, M.; Fujimoto, T.; Sakai, Y. Evidence for ESCRT-and clathrin-dependent microautophagy. J. Cell. Biol. 2017, 216, 3263–3274. [Google Scholar] [CrossRef] [Green Version]
- Cregg, J.M.; Russell, K.A. Transformation. Methods Mol. Biol. 1998, 103, 27–39. [Google Scholar] [CrossRef]
- Baerends, R.J.; Faber, K.N.; Kram, A.M.; Kiel, J.A.; van der Klei, I.J.; Veenhuis, M. A stretch of positively charged amino acids at the N terminus of Hansenula polymorpha Pex3p is involved in incorporation of the protein into the peroxisomal membrane. J. Biol. Chem. 2000, 275, 9986–9995. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant Name | Strain Name | Background | Genotype and Plasmid | Source |
|---|---|---|---|---|
| WT | GS115 | GS115 | his4 | [17] |
| WT | GS200 | GS200 | arg4 his4 | [18] |
| WT | PPY12h | PPY12h | arg4 his4 | [19] |
| WT | PPY12m | PPY12m | arg4 his4 | [19] |
| ape1 | SJCF434 | PPY12m | ∆ape1::GeneticinR arg4 his4 | [20] |
| atg1 | R12 | GS115 | atg1-1::ZeocinR his4 | [21] |
| atg2 | WDK011 | GS115 | ∆atg2::ZeocinR his4 | [21] |
| atg3 | gsa20 | GS115 | atg3::ZeocinR his4 | [21] |
| atg4 | PPM408 | PPY12h | atg4::ZeocinR arg4 his4 | [22] |
| atg5 | SJCF2320 | GS115 | ∆atg5::ZeocinR his4 | SL 1 |
| atg6 | SRDM006 | PPY12m | ∆atg6::GeneticinR arg4 his4 | [23] |
| atg7 | WDK07 | GS200 | ∆atg7::ScARG4 arg4 his4 | [24] |
| atg8 | SJCF925 | PPY12h | ∆atg8::GeneticinR arg4 his4 | [25] |
| atg9 | R19 | GS115 | atg9-1::ZeocinR his4 | [21] |
| atg11 | R8 | GS115 | atg11-2::ZeocinR his4 | [26] |
| atg17 | SJCF929 | PPY12h | ∆atg17::GeneticinR arg4 his4 | [25] |
| atg11 atg17 | SJCF948 | GS115 | atg11-2::ZeocinR ∆atg17::GeneticinR his4 | [25] |
| atg18 | R2 | GS115 | atg18-1::ZeocinR his4 | [21] |
| atg20 | SRDM020 | PPY12m | ∆atg20::GeneticinR arg4 his4 | SL |
| atg24 | paz16 | PPY12h | atg24::ZeocinR arg4 his4 | [27] |
| atg25 | SJCF1231 | PPY12h | ∆atg25::GeneticinR arg4 his4 | [23] |
| atg26 | ∆pdg3 | GS200 | ∆atg26::ScARG4 arg4 his4 | [28] |
| atg28 | ∆pdg2 | GS200 | ∆atg28::ScARG4 arg4 his4 | [29] |
| atg30 | SJCF936 | PPY12h | ∆atg30::ZeocinR arg4 his4 | [25] |
| atg32 | SJCF1715 | PPY12h | ∆atg32::GeneticinR arg4 his4 | [30] |
| atg35 | SVN1 | GS200 | ∆atg35::ScARG4 arg4 his4 | [31] |
| atg37 | STN96 | PPY12h | ∆atg37::GeneticinR arg4 his4 | [32] |
| atg40 | SRK2 | PPY12h | ∆atg40::ZeocinR (pRK4) arg4 his4 | This study |
| pep4 prb1 | SMD1163 | GS115 | pep4 prb1 his4 | [33] |
| pex3 | SEW1 | PPY12h | ∆pex3::PpARG4 arg4 his4 | [34] |
| pex19 | SKF13 | PPY12h | ∆pex19::ZeocinR arg4 his4 | [35] |
| prl1 | SRK3 | PPY12h | prl1::ZeocinR (pRK6) arg4 his4 | This study |
| vac8 | WDY53 | GS200 | ∆vac8::ZeocinR arg4 his4 | [36] |
| vam7 | SRDM050 | PPY12m | ∆vam7::ZeocinR arg4 his4 | [37] |
| vps15 | OP5 | GS200 | ∆vps15::ScARG4 arg4 his4 | [38] |
| vps17 | SRDM122 | PPY12m | ∆vps17::GeneticinR arg4 his4 | [23] |
| uvrag | SRDM083 | PPY12m | ∆uvrag::ZeocinR arg4 his4 | [23] |
| ypt7 | SRRM197 | PPY12h | ∆ypt7::GeneticinR arg4 his4 | [37] |
| Strain (Kp/Sc) | Kp (This Study) | Sc [10] | Sc [13] | Sc [12] | ||
|---|---|---|---|---|---|---|
| SD-N | S-Phase | SD-N | SD-N | S-Phase | SD-D (0.4%) | |
| Erg6-GFP | Erg6-GFP | Erg6-GFP | Faa4-GFP | BODIPY | Erg6-DsRed | |
| WT | + | + | + | + | + | + |
| ape1 | + | + | ND 1 | ND | ND | ND |
| atg1 | − | +/− | − | − | − | − |
| atg2 | − | +/− | ND | ND | − | − |
| atg3 | − | +/− | − | − | − | − |
| atg4 | − | +/− | − | − | − | ND |
| atg5 | − | +/− | − | − | − | − |
| atg6 | − | − | − | − | − | − |
| atg7 | − | +/− | − | − | − | − |
| atg8 | − | +/− | − | − | − | − |
| atg9 | − | +/− | − | − | − | − |
| atg11 | + | + | ND | +/− | + | + |
| atg17 | +/− | +/− | ND | − | − | +/− |
| atg11 atg17 | +/− | +/− | ND | ND | ND | ND |
| atg18 | − | +/− | − | − | − | − |
| atg20 | + | + | ND | + | + | + |
| atg24 | + | + | ND | ND | + | + |
| atg25 | + | + | NA 2 | NA | NA | NA |
| atg26 | + | +/− | ND | ND | + | + |
| atg28/atg29,31 | +/− | +/− | ND | ND | −, − | +/−, +/− |
| atg30/atg36 | + | + | ND | ND | + | + |
| atg32 | + | + | ND | ND | − | +/− |
| atg35 | + | + | NA | NA | NA | NA |
| atg37 | + | + | NA | NA | NA | NA |
| atg40 | + | +/− | ND | ND | ND | ND |
| pep4 prb1 | − | − | ND | ND | − | ND |
| pex3 | + | +/− | ND | ND | ND | ND |
| pex19 | + | +/− | ND | ND | ND | ND |
| prl1 | + | − | NA | NA | NA | NA |
| vac8 | + | + | ND | − | ND | ND |
| vam7 | − | +/− | − | − | ND | ND |
| vps15 | − | +/− | ND | ND | ND | ND |
| vps17 | + | + | ND | ND | ND | ND |
| uvrag/vps38 | + | + | +/− | +/− | ND | ND |
| ypt7 | − | +/− | − | − | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, R.; Rahman, M.A.; Nazarko, T.Y. Nitrogen Starvation and Stationary Phase Lipophagy Have Distinct Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 9094. https://doi.org/10.3390/ijms21239094
Kumar R, Rahman MA, Nazarko TY. Nitrogen Starvation and Stationary Phase Lipophagy Have Distinct Molecular Mechanisms. International Journal of Molecular Sciences. 2020; 21(23):9094. https://doi.org/10.3390/ijms21239094
Chicago/Turabian StyleKumar, Ravinder, Muhammad Arifur Rahman, and Taras Y. Nazarko. 2020. "Nitrogen Starvation and Stationary Phase Lipophagy Have Distinct Molecular Mechanisms" International Journal of Molecular Sciences 21, no. 23: 9094. https://doi.org/10.3390/ijms21239094
APA StyleKumar, R., Rahman, M. A., & Nazarko, T. Y. (2020). Nitrogen Starvation and Stationary Phase Lipophagy Have Distinct Molecular Mechanisms. International Journal of Molecular Sciences, 21(23), 9094. https://doi.org/10.3390/ijms21239094