Extracellular Vesicles in CNS Developmental Disorders

 , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
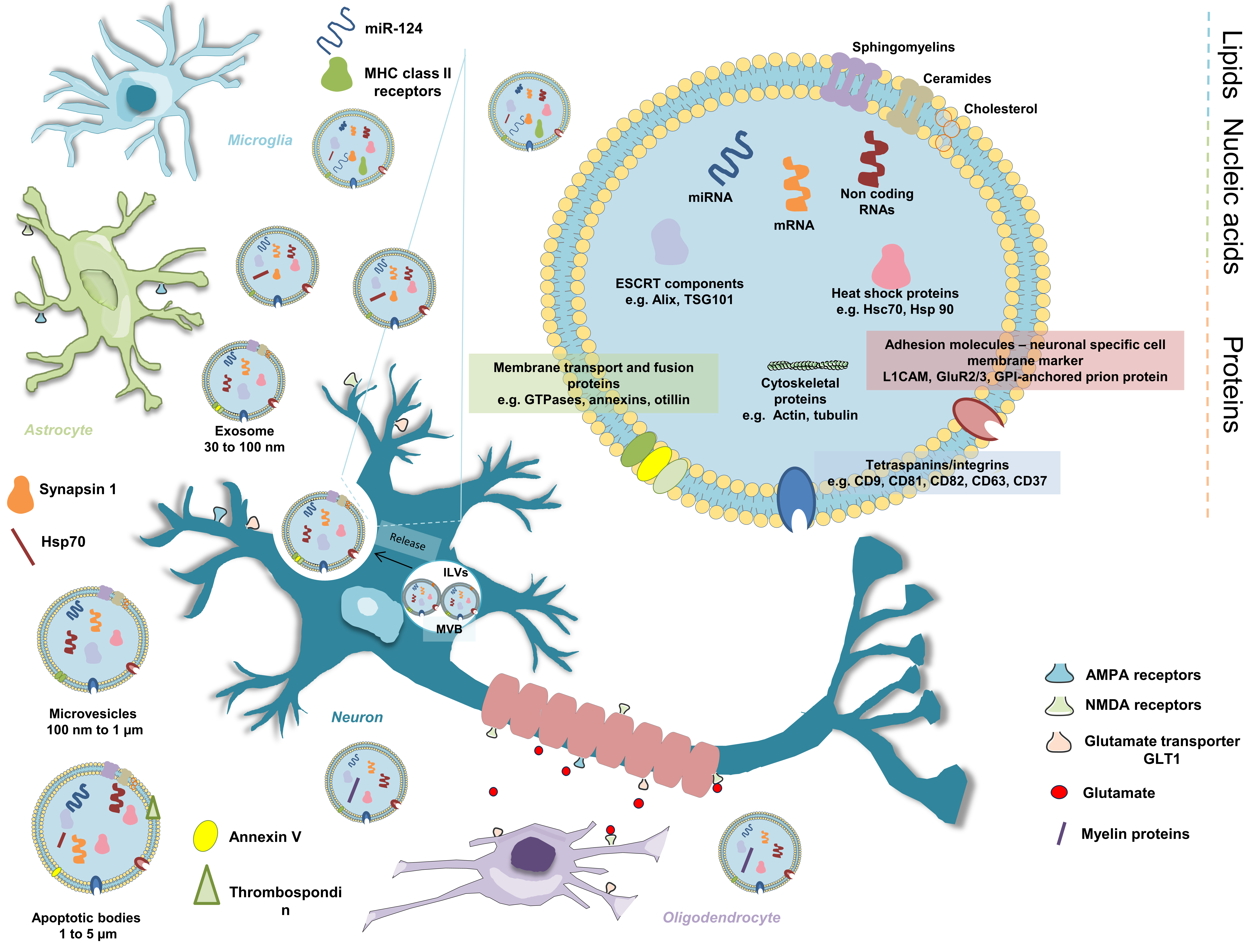
1.1. Extracellular Vesicles
1.2. Molecular Composition of EVs
2. EVs Mediate Communication in CNS—During and Post-Development
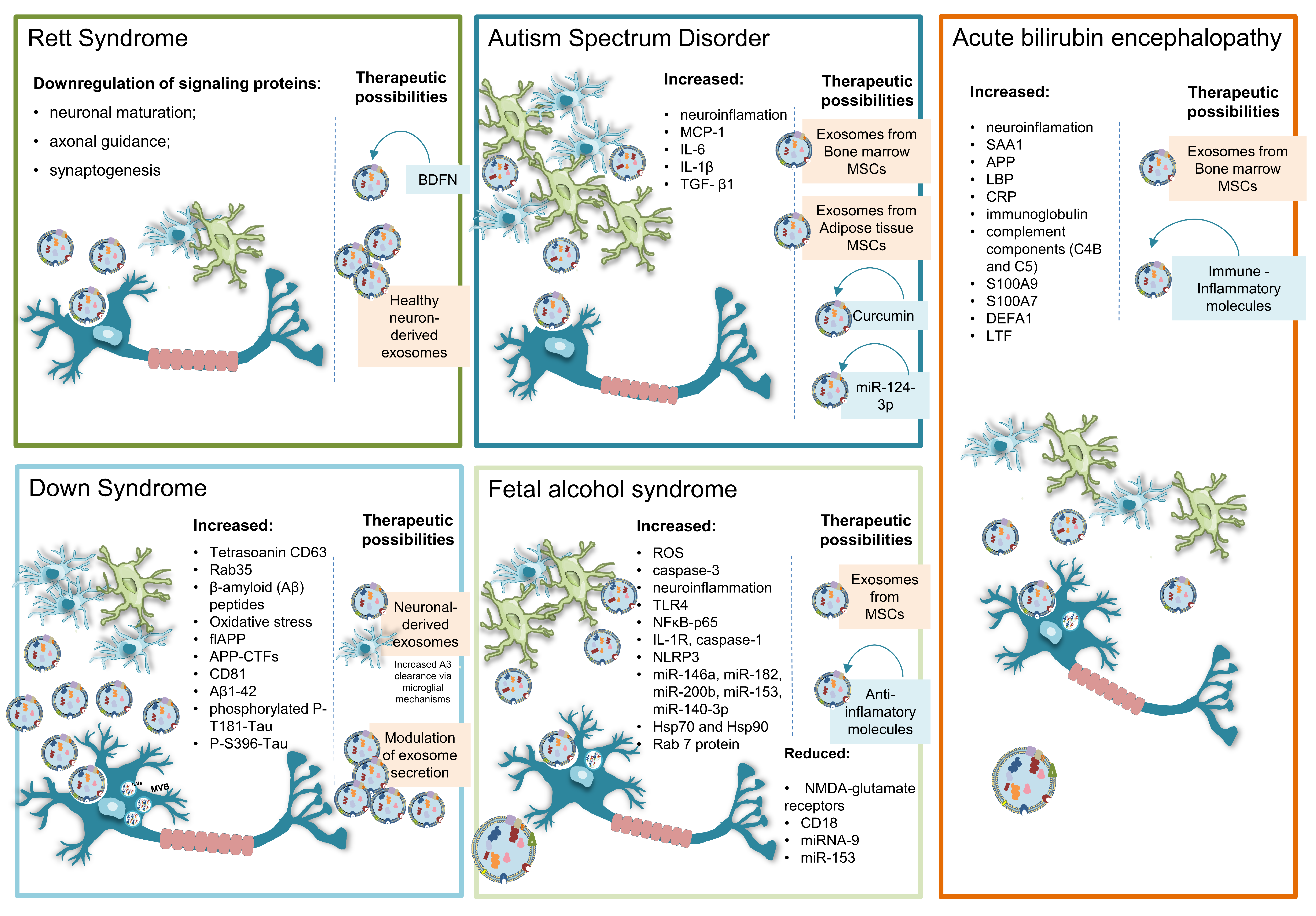
3. EVs in Developmental Pathology of the Nervous System
3.1. Rett Syndrome
3.2. Autism Spectrum Disorder
3.3. Down Syndrome
3.4. Fetal Alcohol Syndrome
3.5. Acute Bilirubin Encephalopathy
4. EVs in Therapy
5. Conclusions/Final Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABE | acute bilirubin encephalopathy |
| ASD | autism spectrum disorder |
| AD | Alzheimer’s disease |
| AMPA | 2-amino-3-(3-hydroxy-5-methyl-isoxazol-4-yl) propanoic acid |
| Aβ | β-amyloid |
| APP | β-amyloid precursor protein |
| APP-CTFs | APP carboxyl-terminal fragments |
| GABA | gamma-aminobutyric acid |
| BBB | blood–brain barrier |
| BDNF | brain-derived neurotrophic factor |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| DS | Down syndrome |
| DYRK1A | dual-specificity tyrosine-phosphorylation-regulated kinase 1A |
| E/I | excitatory/inhibitory imbalance |
| ESCRT | endosomal sorting complex required for transport |
| FAS | fetal alcohol syndrome |
| FASDs | fetal alcohol spectrum disorders |
| EVs | extracellular vesicles |
| flAPP | full-length APP |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| GLAST | glutamate aspartate transporter |
| GLT1 | glucose transporter type 1 |
| GFAP | glial fibrillary acidic protein |
| GluR2/3 | glutamate receptor subunit |
| GPI | glycosylphosphatidylinositol |
| hiPSCs | human induced pluripotent stem cells |
| Hsa21 | chromosome 21 |
| Hsp | heat-shock proteins |
| IC | isogenic control |
| IL | interleukin |
| ILVs | intraluminal vesicles |
| L1CAM L1 | cell adhesion molecule |
| LPS | lipopolysaccharides |
| MeCP2 | methyl-CpG-binding protein 2 |
| MeCP2LOF | methyl-CpG-binding protein 2 loss of function |
| miRNAs | micro RNAs |
| mRNAs | messenger RNA |
| MSCs | mesenchymal stem cells |
| mtDNA | mitochondrial DNA |
| MVs | microvesicles |
| MVBs | multivesicular bodies |
| NPCs | neural progenitor cells |
| NMDA | N-methyl-d-aspartate |
| RG | radial glia |
| RNA | ribonucleic acid |
| ROS | reactive oxygen species |
| RTT | Rett syndrome |
| siRNA | small interfering RNA |
| TGF- β1 | anti-inflammatory cytokine tumor growth factor–β1/transforming growth factor beta 1 |
| TLR4 | toll-like receptor 4 |
| TrkB | tropomyosin-related kinase B |
References
- Budnik, V.; Ruiz-Cañada, C.; Wendler, F. Extracellular vesicles round off communication in the nervous system. Nat. Rev. Neurosci. 2016, 17, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janas, A.M.; Sapoń, K.; Janas, T.; Stowell, M.H.B.; Janas, T. Exosomes and other extracellular vesicles in neural cells and neurodegenerative diseases. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Bond, V.C.; Borràs, F.E.; Breakefield, X.; Budnik, V.; et al. Vesiclepedia: A Compendium for Extracellular Vesicles with Continuous Community Annotation. PLoS Biol. 2012, 10, e1001450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegtel, D.M.; Peferoen, L.; Amor, S. Extracellular vesicles as modulators of cell-to-cell communication in the healthy and diseased brain. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Doyle, L.; Wang, M. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, D.K.; Hvam, M.L.; Primdahl-Bengtson, B.; Boysen, A.T.; Whitehead, B.; Dyrskjøt, L.; Ørntoft, T.F.; Howard, K.A.; Ostenfeld, M.S. Comparative analysis of discrete exosome fractions obtained by differential centrifugation. J. Extracell. Vesicles 2014, 3, 25011. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Jin, K.; Gao, L.; Zhang, Z.; Li, F.; Zhou, F.; Zhang, L. Methods and Technologies for Exosome Isolation and Characterization. Small Methods 2018, 2, 1800021. [Google Scholar] [CrossRef]
- Cocozza, F.; Grisard, E.; Martin-Jaular, L.; Mathieu, M.; Théry, C. SnapShot: Extracellular Vesicles. Cell 2020, 182, 262-262.e1. [Google Scholar] [CrossRef] [PubMed]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct RNA profiles in subpopulations of extracellular vesicles: Apoptotic bodies, microvesicles and exosomes. J. Extracell. Vesicles 2013, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Babst, M. MVB Vesicle Formation: ESCRT-Dependent, ESCRT-Independent and Everything in Between. Curr. Opin. Cell Biol. 2011, 23, 452–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brügger, B.; Simons, M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef] [PubMed]
- Marzano, M.; Bejoy, J.; Cheerathodi, M.R.; Sun, L.; York, S.B.; Zhao, J.; Kanekiyo, T.; Bu, G.; Meckes, D.G.; Li, Y. Differential Effects of Extracellular Vesicles of Lineage-Specific Human Pluripotent Stem Cells on the Cellular Behaviors of Isogenic Cortical Spheroids. Cells 2019, 8, 993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savina, A.; Fader, C.M.; Damiani, M.T.; Colombo, M.I. Rab11 promotes docking and fusion of multivesicular bodies in a calcium-dependent manner. Traffic 2005, 6, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Fauré, J.; Lachenal, G.; Court, M.; Hirrlinger, J.; Chatellard-Causse, C.; Blot, B.; Grange, J.; Schoehn, G.; Goldberg, Y.; Boyer, V.; et al. Exosomes are released by cultured cortical neurones. Mol. Cell. Neurosci. 2006, 31, 642–648. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Didiot, M.C.; Sapp, E.; Leszyk, J.; Shaffer, S.A.; Rockwell, H.E.; Gao, F.; Narain, N.R.; DiFiglia, M.; Kiebish, M.A.; et al. High-resolution proteomic and lipidomic analysis of exosomes and microvesicles from different cell sources. J. Extracell. Vesicles 2016, 5, 32570. [Google Scholar] [CrossRef]
- Record, M.; Silvente-poirot, S.; Poirot, M.; Wakelam, M.J.O. Extracellular vesicles: Lipids as key components of their biogenesis and functions. J. Lipid Res. 2018, 59, 1316–1324. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Boussac, M.; Véron, P.; Ricciardi-Castagnoli, P.; Raposo, G.; Garin, J.; Amigorena, S. Proteomic Analysis of Dendritic Cell-Derived Exosomes: A Secreted Subcellular Compartment Distinct from Apoptotic Vesicles. J. Immunol. 2001, 166, 7309–7318. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Liao, L.; Tian, W. Extracellular Vesicles Derived From Apoptotic Cells: An Essential Link between Death and Regeneration. Front. Cell Dev. Biol. 2020, 8, 573511. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, L.; Regan, M.; Higashimori, H.; Ng, S.K.; Esau, C.; Vidensky, S.; Rothstein, J.; Yang, Y. Neuronal exosomal miRNA-dependent translational regulation of astroglial glutamate transporter GLT1. J. Biol. Chem. 2013, 288, 7105–7116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Men, Y.; Yelick, J.; Jin, S.; Tian, Y.; Chiang, M.S.R.; Higashimori, H.; Brown, E.; Jarvis, R.; Yang, Y. Exosome reporter mice reveal the involvement of exosomes in mediating neuron to astroglia communication in the CNS. Nat. Commun. 2019, 10, 4236. [Google Scholar] [CrossRef] [Green Version]
- Xin, H.; Li, Y.; Buller, B.; Katakowski, M.; Zhang, Y.; Wang, X.; Shang, X.; Zhang, Z.G.; Chopp, M. Exosome-mediated transfer of miR-133b from multipotent mesenchymal stromal cells to neural cells contributes to neurite outgrowth. Stem Cells 2012, 30, 1556–1564. [Google Scholar] [CrossRef] [Green Version]
- Shaimardanova, A.; Solovyeva, V.; Chulpanova, D.; James, V.; Kitaeva, K.; Rizvanov, A. Extracellular vesicles in the diagnosis and treatment of central nervous system diseases. Neural Regen. Res. 2020, 15, 586–596. [Google Scholar]
- Yagi, Y.; Ohkubo, T.; Kawaji, H.; Machida, A.; Miyata, H.; Goda, S.; Roy, S.; Hayashizaki, Y.; Suzuki, H.; Yokota, T. Next-generation sequencing-based small RNA profiling of cerebrospinal fluid exosomes. Neurosci. Lett. 2017, 636, 48–57. [Google Scholar] [CrossRef]
- Marzesco, A.M.; Janich, P.; Wilsch-Bräuninger, M.; Dubreuil, V.; Langenfeld, K.; Corbeil, D.; Huttner, W.B. Release of extracellular membrane particles carrying the stem cell marker prominin-1 (CD133) from neural progenitors and other epithelial cells. J. Cell Sci. 2005, 118, 2849–2858. [Google Scholar] [CrossRef] [Green Version]
- Lachenal, G.; Pernet-Gallay, K.; Chivet, M.; Hemming, F.J.; Belly, A.; Bodon, G.; Blot, B.; Haase, G.; Goldberg, Y.; Sadoul, R. Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol. Cell. Neurosci. 2011, 46, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Bahrini, I.; Song, J.H.; Diez, D.; Hanayama, R. Neuronal exosomes facilitate synaptic pruning by up-regulating complement factors in microglia. Sci. Rep. 2015, 5, 7989. [Google Scholar] [CrossRef] [Green Version]
- Pastuzyn, E.D.; Day, C.E.; Kearns, R.B.; Kyrke-Smith, M.; Taibi, A.V.; McCormick, J.; Yoder, N.; Belnap, D.M.; Erlendsson, S.; Morado, D.R.; et al. The Neuronal Gene Arc Encodes a Repurposed Retrotransposon Gag Protein that Mediates Intercellular RNA Transfer. Cell 2018, 172, 275–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.R.; Robinson, M.B.; Gifondorwa, D.J.; Tytell, M.; Milligan, C.E. Regulation of Heat Shock Protein 70 Release in Astrocytes: Role of Signaling Kinases. Dev. Neurobiol. 2007, 67, 1815–1829. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cesca, F.; Loers, G.; Schweizer, M.; Buck, F.; Benfenati, F.; Schachner, M.; Kleene, R. Synapsin I is an oligomannose-carrying glycoprotein, acts as an oligomannose-binding lectin, and promotes neurite outgrowth and neuronal survival when released via glia-derived exosomes. J. Neurosci. 2011, 31, 7275–7290. [Google Scholar] [CrossRef] [PubMed]
- Frühbeis, C.; Fröhlich, D.; Kuo, W.P.; Amphornrat, J.; Thilemann, S.; Saab, A.S.; Kirchhoff, F.; Möbius, W.; Goebbels, S.; Nave, K.A.; et al. Neurotransmitter-Triggered Transfer of Exosomes Mediates Oligodendrocyte-Neuron Communication. PLoS Biol. 2013, 11, e1001604. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Stoica, B.A.; Loane, D.J.; Yang, M.; Abulwerdi, G.; Khan, N.; Kumar, A.; Thom, S.R.; Faden, A.I. Microglial-derived microparticles mediate neuroinflammation after traumatic brain injury. J. Neuroinflammation 2017, 14, 47. [Google Scholar] [CrossRef] [Green Version]
- Antonucci, F.; Turola, E.; Riganti, L.; Caleo, M.; Gabrielli, M.; Perrotta, C.; Novellino, L.; Clementi, E.; Giussani, P.; Viani, P.; et al. Microvesicles released from microglia stimulate synaptic activity via enhanced sphingolipid metabolism. EMBO J. 2012, 31, 1231–1240. [Google Scholar] [CrossRef]
- Glebov, K.; Löchner, M.; Jabs, R.; Lau, T.; Merkel, O.; Schloss, P.; Steinhäuser, C.; Walter, J. Serotonin stimulates secretion of exosomes from microglia cells. Glia 2015, 63, 626–634. [Google Scholar] [CrossRef]
- Schiera, G.; Di Liegro, C.M.; Di Liegro, I. Cell-to-cell communication in learning and memory: From neuro-and glio-transmission to information exchange mediated by extracellular vesicles. Int. J. Mol. Sci. 2020, 21, 226. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Mesci, P.; Carromeu, C.; McClatchy, D.R.; Schiapparelli, L.; Yates, J.R.; Muotri, A.R.; Cline, H.T. Exosomes regulate neurogenesis and circuit assembly. Proc. Natl. Acad. Sci. USA 2019, 116, 16086–16094. [Google Scholar] [CrossRef] [Green Version]
- Tsilioni, I.; Theoharides, T.C. Extracellular vesicles are increased in the serum of children with autism spectrum disorder, contain mitochondrial DNA, and stimulate human microglia to secrete IL-1β. J. Neuroinflammation 2018, 15, 239. [Google Scholar] [CrossRef]
- D’Acunzo, P.; Hargash, T.; Pawlik, M.; Goulbourne, C.N.; Pérez-González, R.; Levy, E. Enhanced generation of intraluminal vesicles in neuronal late endosomes in the brain of a Down syndrome mouse model with endosomal dysfunction. Dev. Neurobiol. 2019, 79, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Pérez-González, R.; Gauthier, S.A.; Sharma, A.; Miller, C.; Pawlik, M.; Kaur, G.; Kim, Y.; Levy, E. A pleiotropic role for exosomes loaded with the amyloid β precursor protein carboxyl-terminal fragments in the brain of Down syndrome patients. Neurobiol. Aging 2019, 84, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Hamlett, E.D.; Ledreux, A.; Potter, H.; Chial, H.J.; Patterson, D.; Espinosa, J.M.; Bettcher, B.M.; Granholm, A.-C. Exosomal biomarkers in Down syndrome and Alzheimer’s disease. Free Radic. Biol. Med. 2018, 114, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, I.; Jarc, J.; Dassler, E.; Aronica, E.; Iyer, A.; Adle-Biassette, H.; Scharrer, A.; Reischer, T.; Hainfellner, J.A.; Kovacs, G.G. The physiological phosphorylation of tau is critically changed in fetal brains of individuals with Down syndrome. Neuropathol. Appl. Neurobiol. 2018, 44, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Hamlett, E.D.; Goetzl, E.J.; Vasilevko, V.; Heather, A.; Larosa, A.; Clark, D.; Carroll, S.L.; Iragui, M.C.; Mufson, E.J.; Sabbagh, M.; et al. Neuronal exosomes reveal Alzheimer’s disease biomarkers in Down syndrome. Alzheimer’s Dement. 2016, 13, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, F.; Montesinos, J.; Ureña-Peralta, J.R.; Guerri, C.; Pascual, M. TLR4 participates in the transmission of ethanol-induced neuroinflammation via astrocyte-derived extracellular vesicles. J. Neuroinflammation 2019, 16, 136. [Google Scholar] [CrossRef] [Green Version]
- Crenshaw, B.J.; Kumar, S.; Bell, C.R.; Jones, L.B.; Williams, S.D.; Saldanha, S.N.; Joshi, S.; Sahu, R.; Sims, B.; Matthews, Q.L. Alcohol Modulates the Biogenesis and Composition of Microglia-Derived Exosomes. Biology 2019, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Tseng, A.M.; Chung, D.D.; Pinson, M.R.; Salem, N.A.; Eaves, S.E.; Miranda, R.C. Ethanol Exposure Increases miR-140 in Extracellular Vesicles: Implications for Fetal Neural Stem Cell Proliferation and Maturation. Alcohol. Clin. Exp. Res. 2019, 43, 1414–1426. [Google Scholar] [CrossRef]
- Tan, N.; Hu, S.; Hu, Z.; Wu, Z.; Wang, B. Quantitative proteomic characterization of microvesicles/exosomes from the cerebrospinal fluid of patients with acute bilirubin encephalopathy. Mol. Med. Rep. 2020, 22, 1257–1268. [Google Scholar] [CrossRef]
- Perets, N.; Hertz, S.; London, M.; Offen, D. Intranasal administration of exosomes derived from mesenchymal stem cells ameliorates autistic-like behaviors of BTBR mice. Mol. Autism 2018, 9, 57. [Google Scholar] [CrossRef]
- Geffen, Y.; Perets, N.; Horev, R.; Yudin, D.; Oron, O.; Elliott, E.; Marom, E.; Danon, U.; Offen, D. Exosomes derived from adipose mesenchymal stem cells: A potential non-invasive intranasal treatment for autism. Cytotherapy 2020, 22, S49. [Google Scholar] [CrossRef]
- Geffen, Y.; Horev, R.; Perets, N.; Marom, E.; Danon, U.; Offen, D. Immuno-Modulation and neuroprotection mediate the therapeutic effect of exosomes in mice model of autism. Cytotherapy 2020, 22, S49–S50. [Google Scholar] [CrossRef]
- Hamlett, E.D.; LaRosa, A.; Mufson, E.J.; Fortea, J.; Ledreux, A.; Granholm, A.C. Exosome release and cargo in Down syndrome. Dev. Neurobiol. 2019, 79, 639–655. [Google Scholar] [CrossRef] [PubMed]
- Goetzl, L.; Goetzl, E. Purification, Extraction and Analyses of Fetal Neurally-Derived Exosomes in Maternal Blood and Neonatal Neurally-Derived Exosomes from Neonatal Blood. U.S. Patent Application No. 16/068,738, 24 January 2019. [Google Scholar]
- Ip, J.P.K.; Mellios, N.; Sur, M. Rett syndrome: Insights into genetic, molecular and circuit mechanisms. Nat. Rev. Neurosci. 2018, 19, 368–382. [Google Scholar] [CrossRef] [PubMed]
- Chahrour, M.; Zoghbi, H.Y. Review the Story of Rett Syndrome: From Clinic to Neurobiology. Neuron 2007, 56, 422–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetto, M.C.N.; Carromeu, C.; Acab, A.; Yu, D.; Yeo, G.W.; Mu, Y.; Chen, G.; Gage, F.H.; Muotri, A.R. A Model for Neural Development and Treatment of Rett Syndrome Using Human Induced Pluripotent Stem Cells. Cell 2010, 143, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Kim, J.; Zhou, L.; Wengert, E.; Zhang, L.; Wu, Z.; Carromeu, C.; Muotri, A.R. KCC2 rescues functional deficits in human neurons derived from patients with Rett syndrome. Proc. Natl. Acad. Sci. USA 2016, 113, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.J.; Tsytsarev, V.; Erzurumlu, R.S. Structural and functional differences in the barrel cortex of Mecp2 null mice. J. Comp. Neurol. 2017, 525, 3951–3961. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, T.; Vaz, S.; Silva, T.; Bekman, E.; Xapelli, S.; Duarte, S.; Ghazvini, M.; Gribnau, J.; Muotri, A.; et al. Modelling Rett Syndrome with Human Patient-Specific Forebrain Organoids. Front. Cell Dev. Biol. 2020, 8, 1–18. [Google Scholar] [CrossRef]
- Abuhatzira, L.; Makedonski, K.; Kaufman, Y.; Razin, A.; Shemer, R. MeCP2 deficiency in the brain decreases BDNF levels by REST/CoREST-mediated repression and increases TRKB production. Epigenetics 2007, 2, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Larimore, J.L.; Chapleau, C.A.; Kudo, S.; Theibert, A.; Percy, K.; Pozzo-Miller, L. Bdnf Overexpression in Hippocampal Neurons Prevents Dendritic Atrophy Caused by Rett-Associated MECP2 Mutations. Neurobiol. Dis. 2009, 34, 199–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Pozzo-Miller, L. BDNF deregulation in Rett syndrome. Neuropharmacology 2014, 76, 737–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suire, C.; Eitan, E.; Shaffer, N.C.; Tian, Q.; Studenski, S.; Mattson, M.P.; Kapogiannis, D. Walking Speed Decline in Older Adults is Associated with Elevated Pro-BDNF in Plasma Extracellular Vesicles. Exp. Gerontol. 2017, 98, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Pramparo, T.; Gazestani, V.H.; Lombardo, M.V.; Pierce, K.; Lewis, N.E. The ASD Living Biology: From cell proliferation to clinical phenotype. Mol. Psychiatry 2019, 24, 88–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariani, J.; Coppola, G.; Zhang, P.; Abyzov, A.; Provini, L.; Tomasini, L.; Amenduni, M.; Szekely, A.; Palejev, D.; Wilson, M.; et al. FOXG1-Dependent Dysregulation of GABA/Glutamate Neuron Differentiation in Autism Spectrum Disorders. Cell 2015, 162, 375–390. [Google Scholar] [CrossRef] [Green Version]
- Courchesne, E.; Karns, C.M.; Davis, H.R.; Ziccardi, R.; Carper, R.A.; Tigue, Z.D.; Chisum, H.J.; Moses, P.; Pierce, K.; Lord, C.; et al. Unusual brain growth patterns in early life in patients with autistic disorder: An MRI study. Neurology 2001, 57, 245–254. [Google Scholar] [CrossRef]
- Matta, S.M.; Hill-Yardin, E.L.; Crack, P.J. The influence of neuroinflammation in Autism Spectrum Disorder. Brain Behav. Immun. 2019, 79, 75–90. [Google Scholar] [CrossRef]
- Pascual, M.; Ibáñez, F.; Guerri, C. Exosomes as mediators of neuron-glia communication in neuroinflammation. Neural Regen. Res. 2020, 15, 796–801. [Google Scholar] [CrossRef]
- Vargas, D.L.; Nascimbene, C.; Krishnan, C.; Zimmerman, A.W.; Pardo, C.A. Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann. Neurol. 2005, 57, 67–81. [Google Scholar] [CrossRef]
- Zhang, B.; Angelidou, A.; Alysandratos, K.D.; Vasiadi, M.; Francis, K.; Asadi, S.; Theoharides, A.; Sideri, K.; Lykouras, L.; Kalogeromitros, D.; et al. Mitochondrial DNA and anti-mitochondrial antibodies in serum of autistic children. J. Neuroinflammation 2010, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- El-Mahalaway, A.M.; El-Azab, N.E.E. The potential neuroprotective role of mesenchymal stem cell-derived exosomes in cerebellar cortex lipopolysaccharide-induced neuroinflammation in rats: A histological and immunohistochemical study. Ultrastruct. Pathol. 2020, 3, 159–173. [Google Scholar] [CrossRef]
- Poleg, S.; Golubchik, P.; Offen, D.; Weizman, A. Cannabidiol as a suggested candidate for treatment of autism spectrum disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 89, 90–96. [Google Scholar] [CrossRef]
- Gabrielli, M.; Battista, N.; Riganti, L.; Prada, I.; Antonucci, F.; Cantone, L.; Lombardi, M.; Matteoli, M.; Maccarrone, M.; Verderio, C. Active endocannabinoids are secreted on the surface of microglial microvesicles. EMBO Rep. 2015, 16, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Ge, X.; Yu, J.; Han, Z.; Yin, Z.; Li, Y.; Chen, F.; Wang, H.; Zhang, J.; Lei, P. Increased miR-124-3p in microglial exosomes following traumatic brain injury inhibits neuronal inflammation and contributes to neurite outgrowth via their transfer into neurons. FASEB J. 2018, 32, 512–528. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.; Xiang, X.; Grizzle, W.; Sun, D.; Zhang, S.; Axtell, R.C.; Ju, S.; Mu, J.; Zhang, L.; Steinman, L.; et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol. Ther. 2011, 19, 1769–1779. [Google Scholar] [CrossRef]
- Renner, M.; Lancaster, M.A.; Bian, S.; Choi, H.; Chung, K.; Knoblich, J.A.; Ku, T.; Peer, A. Self-organized developmental patterning and differentiation in cerebral organoids. EMBO J. 2017, 36, 1316–1329. [Google Scholar] [CrossRef]
- Silva, T.P.; Bekman, E.; Fernandes, T.G.; Vaz, S.H.; Carlos, A.; Diogo, M.M.; Cabral, J.M.; Carmo-fonseca, M. Maturation of human pluripotent stem cell-derived cerebellar neurons in the absence of co-culture. Front. Bioeng. Biotechnol. 2020, 8, 70. [Google Scholar] [CrossRef]
- Holtzman, D.M.; Santucci, D.; Kilbridge, J.; Chua-Couzens, J.; Fontana, D.J.; Daniels, S.E.; Johnson, R.M.; Chen, K.; Sun, Y.; Carlson, E.; et al. Developmental abnormalities and age-related neurodegeneration in a mouse model of Down syndrome. Proc. Natl. Acad. Sci. USA 1996, 93, 13333–13338. [Google Scholar] [CrossRef] [Green Version]
- Hamlett, E.D.; Boger, H.A.; Ledreux, A.; Kelley, C.M.; Mufson, E.J.; Falangola, M.F.; Guilfoyle, D.N.; Nixon, R.A.; Patterson, D.; Duval, N.; et al. Cognitive Impairment, Neuroimaging, and Alzheimer Neuropathology in Mouse Models of Down Syndrome. Curr. Alzheimer Res. 2015, 13, 35–52. [Google Scholar] [CrossRef]
- Cataldo, A.M.; Peterhoff, C.M.; Troncoso, J.C.; Gomez-Isla, T.; Hyman, B.T.; Nixon, R.A. Endocytic pathway abnormalities precede amyloid β deposition in sporadic alzheimer’s disease and down syndrome: Differential effects of APOE genotype and presenilin mutations. Am. J. Pathol. 2000, 157, 277–286. [Google Scholar] [CrossRef]
- Gauthier, S.A.; Pérez-González, R.; Sharma, A.; Huang, F.K.; Alldred, M.J.; Pawlik, M.; Kaur, G.; Ginsberg, S.D.; Neubert, T.A.; Levy, E. Enhanced exosome secretion in Down syndrome brain—A protective mechanism to alleviate neuronal endosomal abnormalities. Acta Neuropathol. Commun. 2017, 5, 65. [Google Scholar] [CrossRef]
- Gyure, K.A.; Durham, R.; Stewart, W.F.; Smialek, J.E.; Troncoso, J.C. Intraneuronal Aβ-amyloid precedes development of amyloid plaques in Down syndrome. Arch. Pathol. Lab. Med. 2001, 125, 489–492. [Google Scholar]
- Yuyama, K.; Sun, H.; Usuki, S.; Sakai, S.; Hanamatsu, H.; Mioka, T.; Kimura, N.; Okada, M.; Tahara, H.; Furukawa, J.I.; et al. A potential function for neuronal exosomes: Sequestering intracerebral amyloid-β peptide. FEBS Lett. 2015, 589, 84–88. [Google Scholar] [CrossRef]
- Baden, R.; Retallack, R.; Hilbich, C.; Simms, G.; Multhaup, G.; Martins, R.; Hockey, A.; Montgomery, P.; Beyreuther, K.; Masters, C.L. Amyloid A4 and its precursor in Down’s syndrome and Alzheimer’s disease. N. Engl. J. Med. 1989, 320, 1446–1452. [Google Scholar]
- Schupf, N.; Patel, B.; Silverman, W.; Zigman, W.B.; Zhong, N.; Tycko, B.; Mehta, P.D.; Mayeux, R. Elevated plasma amyloid b-peptide 1 ± 42 and onset of dementia in adults with Down syndrome. Neurosci. Lett. 2001, 301, 199–203. [Google Scholar] [CrossRef]
- Refolo, L.M.; Sambamurti, K.; Efthimiopoulos, S.; Pappolla, M.A.; Robakis, N.K. Evidence that secretase cleavage of cell surface Alzheimer amyloid precursor occurs after normal endocytic internalization. J. Neurosci. Res. 1995, 40, 694–706. [Google Scholar] [CrossRef]
- Pérez-González, R.; Kim, Y.; Miller, C.; Pacheco-Quinto, J.; Eckman, E.A.; Levy, E. Extracellular vesicles: Where the amyloid precursor protein carboxyl-terminal fragments accumulate and amyloid-β oligomerizes. FASEB J. 2020, 34, 12922–12931. [Google Scholar] [CrossRef]
- Yin, X.; Jin, N.; Shi, J.; Zhang, Y.; Wu, Y.; Gong, C.X.; Iqbal, K.; Liu, F. Dyrk1A overexpression leads to increase of 3R-tau expression and cognitive deficits in Ts65Dn Down syndrome mice. Sci. Rep. 2017, 7, 619. [Google Scholar] [CrossRef]
- Winston, C.N.; Aulston, B.; Rockenstein, E.M.; Adame, A.; Prikhodko, O.; Dave, K.N.; Mishra, P.; Rissman, R.A.; Yuan, S.H. Neuronal exosome-derived human tau is toxic to recipient mouse neurons in vivo. J. Alzheimer’s Dis. 2019, 67, 541–553. [Google Scholar] [CrossRef]
- Podvin, S.; Jones, A.; Liu, Q.; Aulston, B.; Ransom, L.; Ames, J.; Shen, G.; Lietz, C.B.; Jiang, Z.; O’Donoghue, A.J.; et al. Dysregulation of exosome cargo by mutant tau expressed in human-induced pluripotent stem cell (iPSC) neurons revealed by proteomics analyses. Mol. Cell. Proteom. 2020, 19, 1017–1034. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Mitsutake, S.; Igarashi, Y. Sphingolipid-modulated exosome secretion promotes clearance of amyloid-β by microglia. J. Biol. Chem. 2012, 287, 10977–10989. [Google Scholar] [CrossRef] [Green Version]
- Mahnke, A.H.; Adams, A.M.; Wang, A.Z.; Miranda, R.C. Toxicant and teratogenic effects of prenatal alcohol. Curr. Opin. Toxicol. 2019, 14, 29–34. [Google Scholar] [CrossRef]
- Moore, E.M.; Migliorini, R.; Infante, M.A.; Riley, E.P. Fetal Alcohol Spectrum Disorders: Recent Neuroimaging Findings. Curr. Dev. Disord. Rep. 2014, 1, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Young, C.; Roth, K.A.; Klocke, B.J.; West, T.; Holtzman, D.M.; Labruyere, J.; Qin, Y.Q.; Dikranian, K.; Olney, J.W. Role of caspase-3 in ethanol-induced developmental neurodegeneration. Neurobiol. Dis. 2005, 20, 608–614. [Google Scholar] [CrossRef]
- Olney, J.W.; Tenkova, T.; Dikranian, K.; Qin, Y.Q.; Labruyere, J.; Ikonomidou, C. Ethanol-induced apoptotic neurodegeneration in the developing C57BL/6 mouse brain. Dev. Brain Res. 2002, 133, 115–126. [Google Scholar] [CrossRef]
- Light, K.E.; Brown, D.P.; Newton, B.W.; Belcher, S.M.; Kane, C.J.M. Ethanol-induced alterations of neurotrophin receptor expression on Purkinje cells in the neonatal rat cerebellum. Brain Res. 2002, 924, 71–81. [Google Scholar] [CrossRef]
- Mahnke, A.H.; Salem, N.A.; Tseng, A.M.; Chung, D.D.; Miranda, R.C. Nonprotein-coding RNAs in Fetal Alcohol Spectrum Disorders. Prog. Mol. Biol. Sci. 2018, 157, 299–342. [Google Scholar]
- Pappalardo-Carter, D.L.; Balaraman, S.; Sathyan, P.; Carter, E.S.; Chen, J.A.; Miranda, R.C. Suppression and epigenetic regulation of miR-9 contributes to ethanol teratology: Evidence from zebrafish and murine fetal neural stem cell models. Alcohol. Clin. Exp. Res. 2013, 37, 1657–1667. [Google Scholar] [CrossRef] [Green Version]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 2004, 5, R13. [Google Scholar] [CrossRef] [Green Version]
- Tsai, P.; Bake, S.; Balaraman, S.; Rawlings, J.; Holgate, R.R.; Dubois, D.; Miranda, R.C. MiR-153 targets the nuclear factor-1 family and protects against teratogenic effects of ethanol exposure in fetal neural stem cells. Biol. Open 2014, 3, 741–758. [Google Scholar] [CrossRef] [Green Version]
- Tal, T.L.; Franzosa, J.A.; Tilton, S.C.; Philbrick, K.A.; Iwaniec, U.T.; Turner, R.T.; Waters, K.M.; Tanguay, R.L. MicroRNAs control neurobehavioral development and function in zebrafish. FASED J. 2012, 26, 1452–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhutani, V.K.; Johnson-Hamerman, L. The clinical syndrome of bilirubin-induced neurologic dysfunction. Semin. Fetal Neonatal Med. 2015, 20, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Bahr, T.M.; Christensen, R.D.; Agarwal, A.M.; George, T.I.; Bhutani, V.K. The Neonatal Acute Bilirubin Encephalopathy Registry (NABER): Background, Aims, and Protocol. Neonatology 2019, 115, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Brites, D. The evolving landscape of neurotoxicity by unconjugated bilirubin: Role of glial cells and inflammation. Front. Pharmacol. 2012, 3, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watchko, J.F.; Tiribelli, C. Bilirubin-Induced Neurologic Damage—Mechanisms and Management Approaches. N. Engl. J. Med. 2013, 369, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Qaisiya, M.; Brischetto, C.; Jašprová, J.; Vitek, L.; Tiribelli, C.; Bellarosa, C. Bilirubin-induced ER stress contributes to the inflammatory response and apoptosis in neuronal cells. Arch. Toxicol. 2017, 91, 1847–1858. [Google Scholar] [CrossRef]
- Saeedi, S.; Israel, S.; Nagy, C.; Turecki, G. The emerging role of exosomes in mental disorders. Transl. Psychiatry 2019, 9, 122. [Google Scholar] [CrossRef]
- Kwon, S.; Shin, S.; Do, M.; Oh, B.H.; Song, Y.; Bui, V.D.; Lee, E.S.; Jo, D.-G.; Cho, Y.W.; Kim, D.-H.; et al. Engineering approaches for effective therapeutic applications based on extracellular vesicles. J. Control. Release 2020, 20. [Google Scholar] [CrossRef]
- Fu, S.; Wang, Y.; Xia, X.; Zheng, J.C. Exosome engineering: Current progress in cargo loading and targeted delivery. NanoImpact 2020, 20, 100261. [Google Scholar] [CrossRef]
- Rufino-Ramos, D.; Albuquerque, P.R.; Carmona, V.; Perfeito, R.; Nobre, R.J.; Pereira de Almeida, L. Extracellular vesicles: Novel promising delivery systems for therapy of brain diseases. J. Control. Release 2017, 262, 247–258. [Google Scholar] [CrossRef]
- De Almeida Fuzeta, M.; Bernardes, N.; Oliveira, F.; Costa, A.; Fernandes-Platzgummer, A.; Farinha, J.; Rodrigues, C.; Jung, S.; Tseng, R.-J.; Milligan, W.; et al. Scalable Production of Human Mesenchymal Stromal Cell-Derived Extracellular Vesicles Under Serum-/Xeno-Free Conditions in a Microcarrier-Based Bioreactor Culture System. Front. Cell Dev. Biol. 2020, 8, 553444. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Disease | EVs—Type and Source | EVs—Cargo Alterations in Disease Context | Reference |
|---|---|---|---|
| Rett syndrome (RTT) | Exosomes from hiPSC-derived neurons (from both isogenic control and MeCP2LOF-disesase cell lines) | Proteomic analysis revealed a downregulation in neurodevelopmental signaling proteins associated with neuronal maturation, axonal guidance and synaptogenesis. | [39] |
| Autism spectrum disorder (ASD) | EVs isolated from ASD children’s serum | Significant increase in total protein concentration and in the amount of mtDNA in EVs. | [40] |
| Down syndrome (DS) | Exosomes from a Ts2 mice model with DS-like phenotype | Increased levels of exosomes in DS models influenced by larger and more abundant number of MVBs and more ILVs per neuron; Neuronal exosomes with a homeostatic role for neurotoxic material release in response to chronic endosomal dysfunction. | [41] |
| Exosomes isolated from DS patients, Ts2 mouse brains and human DS fibroblasts | Enriched in APP carboxyl-terminal fragments (APP-CTFs) and in full-length APP (flAPP). | [42] | |
| Exosomes from blood samples from DS patients | Increased CD81 levels (more abundant neuronal exosomes secreted). Neuronal exosomes contained Aβ peptide products and hyper-phosphorylated species of Tau (P-Tau). | [43,44] | |
| Exosomes from blood samples of DS patients | DS neuronal exosomes showed higher levels of Aβ1-42, phosphorylated P-T181-Tau and P-S396-Tau. | [45] | |
| Fetal alcohol syndrome (FAS) | EVs from cultured neurons and astrocytes (ethanol-treated) | EVs from the treated condition are increased in number, with higher content of inflammatory-related proteins, such as TLR4, NFκB-p65, IL-1R, caspase-1 and NLRP3, as well as miRNAs (miR-146a, miR-182 and miR-200b). | [46] |
| Exosomes from microglia BV-2 cell lines (exposed to ethanol during biogenesis) | Decreased levels of CD18 (a microglial and immune cell marker). Both Hsp70 and Hsp90 were increased (preventing damaging pro-inflammatory responses). Decreased expression of Rab 7 protein, (important role in vesicle trafficking and exosome biogenesis). | [47] | |
| EVs from an in vitro model of NSCs (exposed to ethanol) | miR-140-3p was identified to be increased during ethanol treatment, which could influence neurogenesis inhibition and neuronal alterations. | [48] | |
| Acute bilirubin encephalopathy (ABE) | EVs isolated from the CSF of ABE patients | Differentially expressed proteins associated with immune-inflammatory response, such as SAA1, APP, LBP, CRP, immunoglobulin and complement components (C4B and C5). Altered levels of specific EVs cargo, mainly S100A9, S100A7, DEFA1 and LTF. | [49] |
| Disease | EVs—Type and Source | EVs—Culture/Administration | EVs—Therapeutic Effect | Reference |
|---|---|---|---|---|
| Rett syndrome (RTT) | Exosomes extracted from IC hiPSC-derived neurons | Exosomes cultured with MeCP2LOF hiPSC-derived neurons | Increased/improved: Puncta densities; Synaptogenesis; Neuronal activity (higher network synchronization); Proliferation; Neuronal fate in developing neural cultures. | [39] |
| Autism spectrum disorder (ASD) | EVs isolated from ASD children’s serum | EVs cultured with human microglia cells | Increased Secretion of IL-1β, a pro-inflammatory cytokine. | [40] |
| Exosomes secreted by mesenchymal stem cells (MSCs) | Exosomes were intranasally administrated in a BTBR mouse model (presents autistic-like behaviors and ASD symptoms) | Increased: Male to male social interaction; Reduced: Repetitive behaviors; miRNA-143 cargo (an immunomodulatory effector in the host cells). | [50] | |
| Exosomes from adipose-derived MSCs | Exosomes were intranasally and intravenously administrated into BTBR and Shank3 mice models (ASD cognitive and motor impairments) | Improved: ASD behavioral phenotype (mainly by non-invasive intranasal administration). | [51] | |
| Exosomes from adipose-derived MSCs | Exosomes cultured with primary neuronal cell cultures, prepared from a newborn SHANK3 homozygote mouse model of autism. | Upregulated: Proteins related to anti-inflammatory processes; Proteins related to immunomodulation; BDNF (neuroprotection and neurogenesis mediator). | [52] | |
| Down syndrome (DS) | Neuronal-derived exosomes purified from the blood of individuals with DS-AD and controls | Exosomes were injected into a control mouse model | Increased: P-Tau amount in pyramidal neurons and in the dentate gyrus of the hippocampus; Spread of toxic P-Tau species via exosome mediation (unpublished work) | [53] |
| Fetal alcohol syndrome (FAS) | EVs from cultured neurons and astrocytes (ethanol-treated) | Ethanol-treated EVs from astrocytes were incubated with cortical neural cultures | Increased: Levels of the inflammatory protein COX-2; miRNAs: miR-146a (regulation of genes related to inflammatory pathways) | [46] |
| Disease | EVs—Type and Source | EVs—Biomarker Potential | Reference |
|---|---|---|---|
| Down syndrome (DS) | Exosomes from DS patients’ blood samples | Increased levels of: CD81; Aβ peptide products; Hyper-phosphorylated species of Tau (P-Tau) | [43,44] |
| Neural origin exosomes extracted from DS patients’ blood samples | Elevated levels of: Aβ1-42; Phosphorylated P-T181-Tau; P-S396-Tau | [45] | |
| Fetal alcohol syndrome (FAS) | Fetal neural exosomes isolated from maternal plasma | Significantly lower levels of: HSF1; Bcl–XL; REST; Synaptophysin; Synaptotagmin; Synaptopodin; GAP–43 | [54] |
| Acute bilirubin encephalopathy (ABE) | EVs isolated from the CSF of ABE patients | Upregulated: LTF and DEFA1 C4B and C5 (complement-associated proteins) Downregulated: S100A7 and S100A9 | [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, A.R.; Sangani, N.B.; Fernandes, T.G.; Diogo, M.M.; Curfs, L.M.G.; Reutelingsperger, C.P. Extracellular Vesicles in CNS Developmental Disorders. Int. J. Mol. Sci. 2020, 21, 9428. https://doi.org/10.3390/ijms21249428
Gomes AR, Sangani NB, Fernandes TG, Diogo MM, Curfs LMG, Reutelingsperger CP. Extracellular Vesicles in CNS Developmental Disorders. International Journal of Molecular Sciences. 2020; 21(24):9428. https://doi.org/10.3390/ijms21249428
Chicago/Turabian StyleGomes, Ana Rita, Nasim Bahram Sangani, Tiago G. Fernandes, M. Margarida Diogo, Leopold M. G. Curfs, and Chris P. Reutelingsperger. 2020. "Extracellular Vesicles in CNS Developmental Disorders" International Journal of Molecular Sciences 21, no. 24: 9428. https://doi.org/10.3390/ijms21249428
APA StyleGomes, A. R., Sangani, N. B., Fernandes, T. G., Diogo, M. M., Curfs, L. M. G., & Reutelingsperger, C. P. (2020). Extracellular Vesicles in CNS Developmental Disorders. International Journal of Molecular Sciences, 21(24), 9428. https://doi.org/10.3390/ijms21249428