Many Different LINE-1 Retroelements Are Activated in Bladder Cancer

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
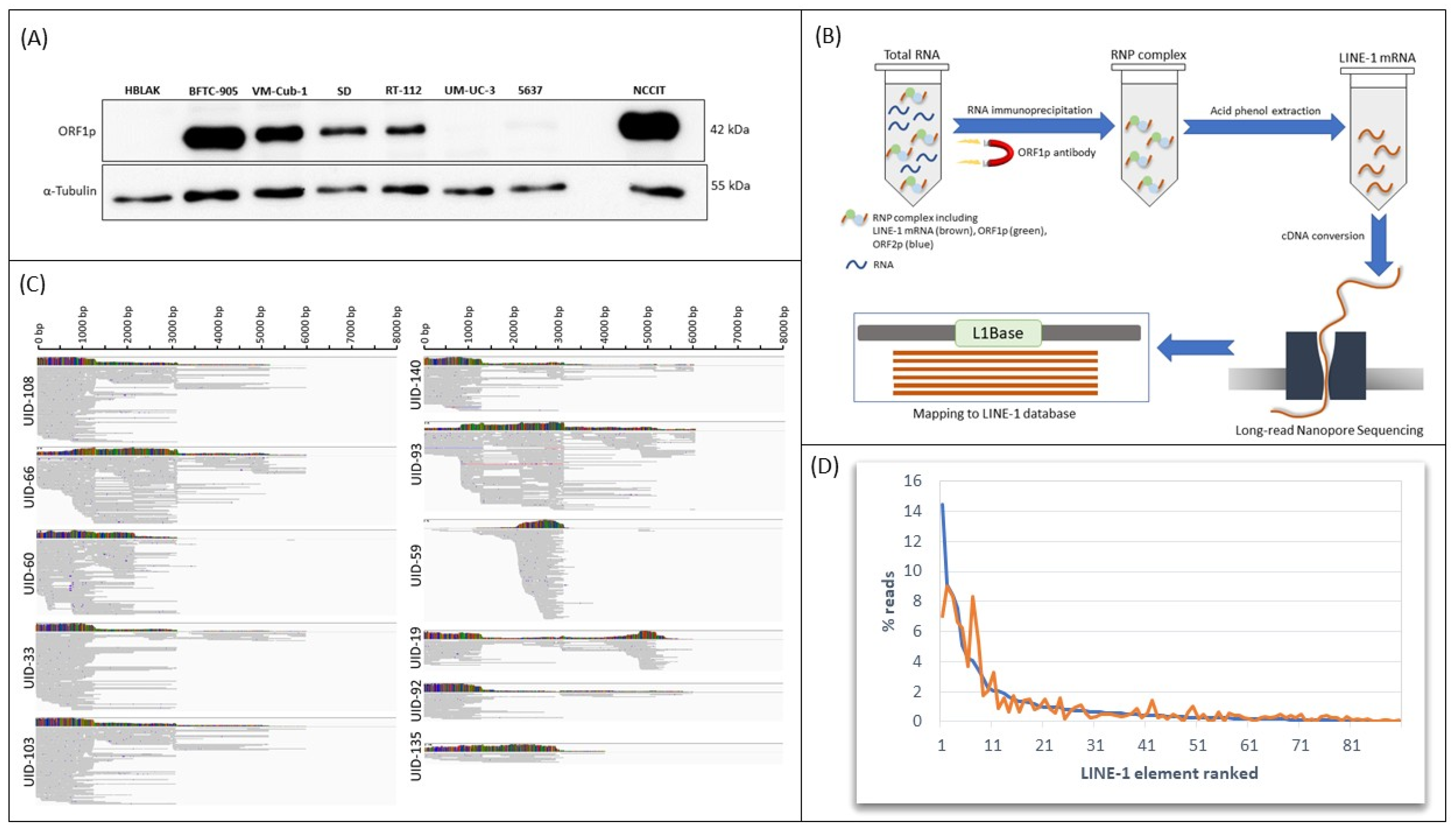
2.1. Delineation of L1 Expression by RNA Immunoprecipitation and Nanopore Sequencing
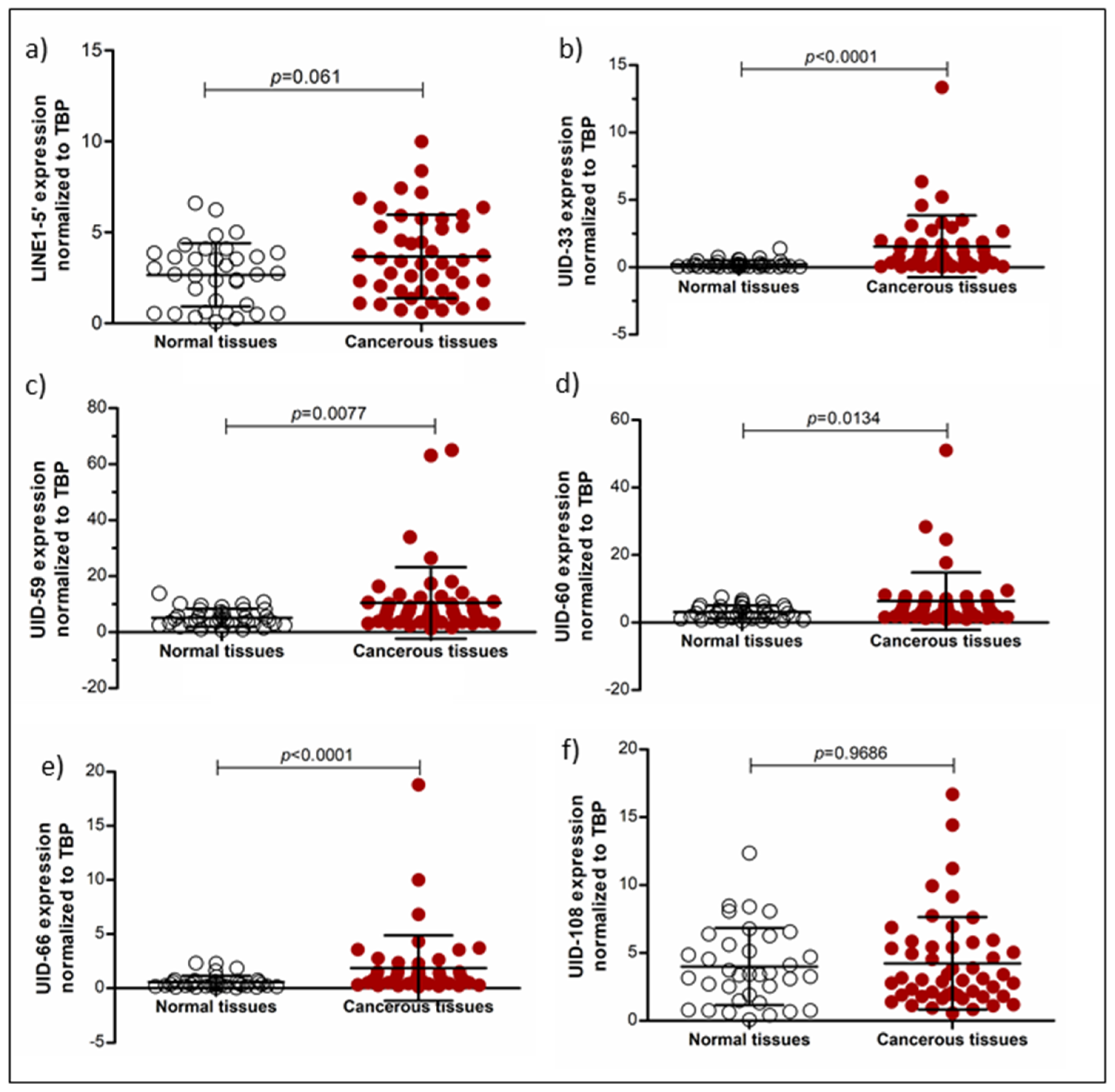
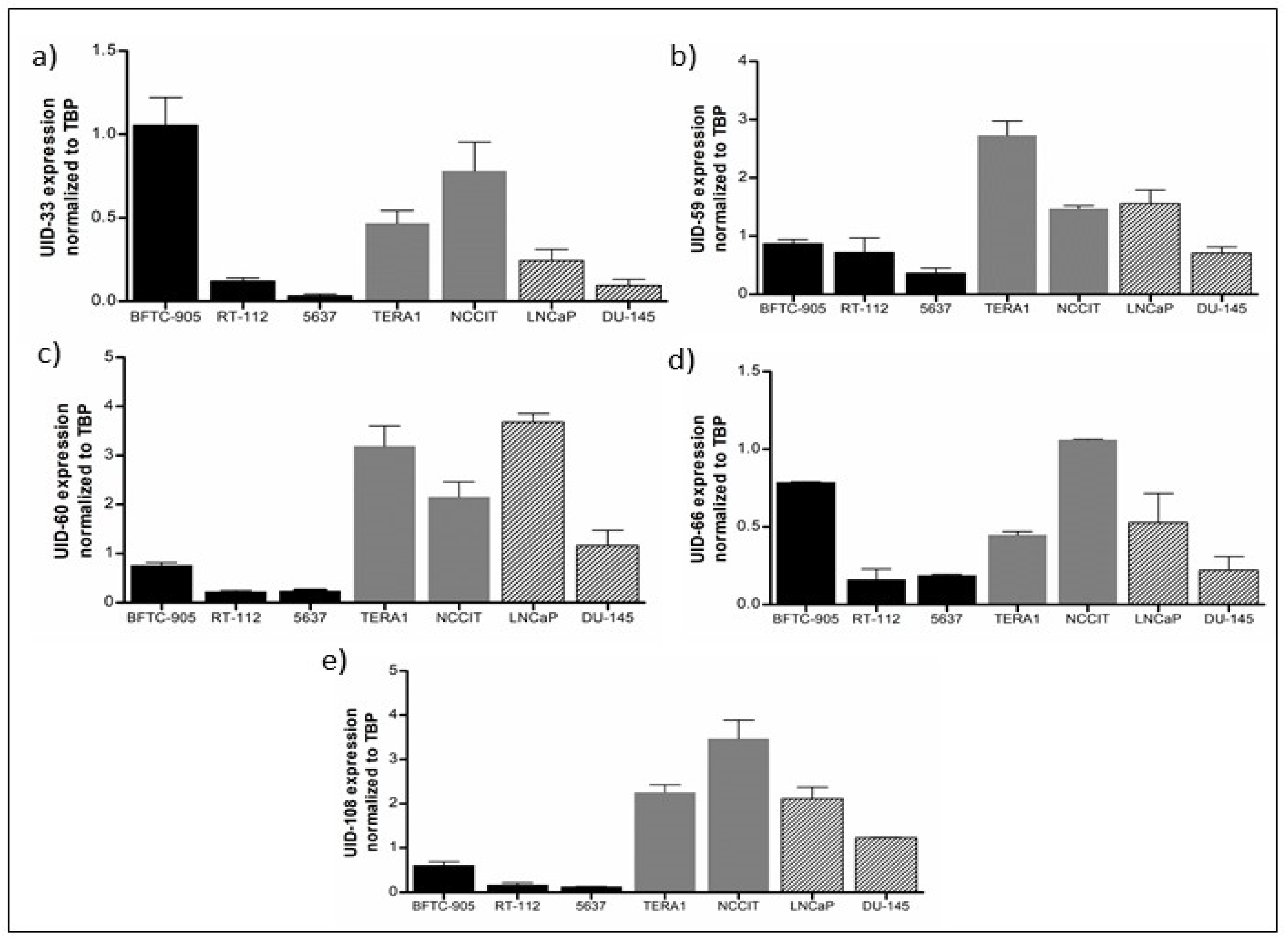
2.2. Analysis of L1 Expression by qRT-PCR in Cancer Cell Lines and Tissues
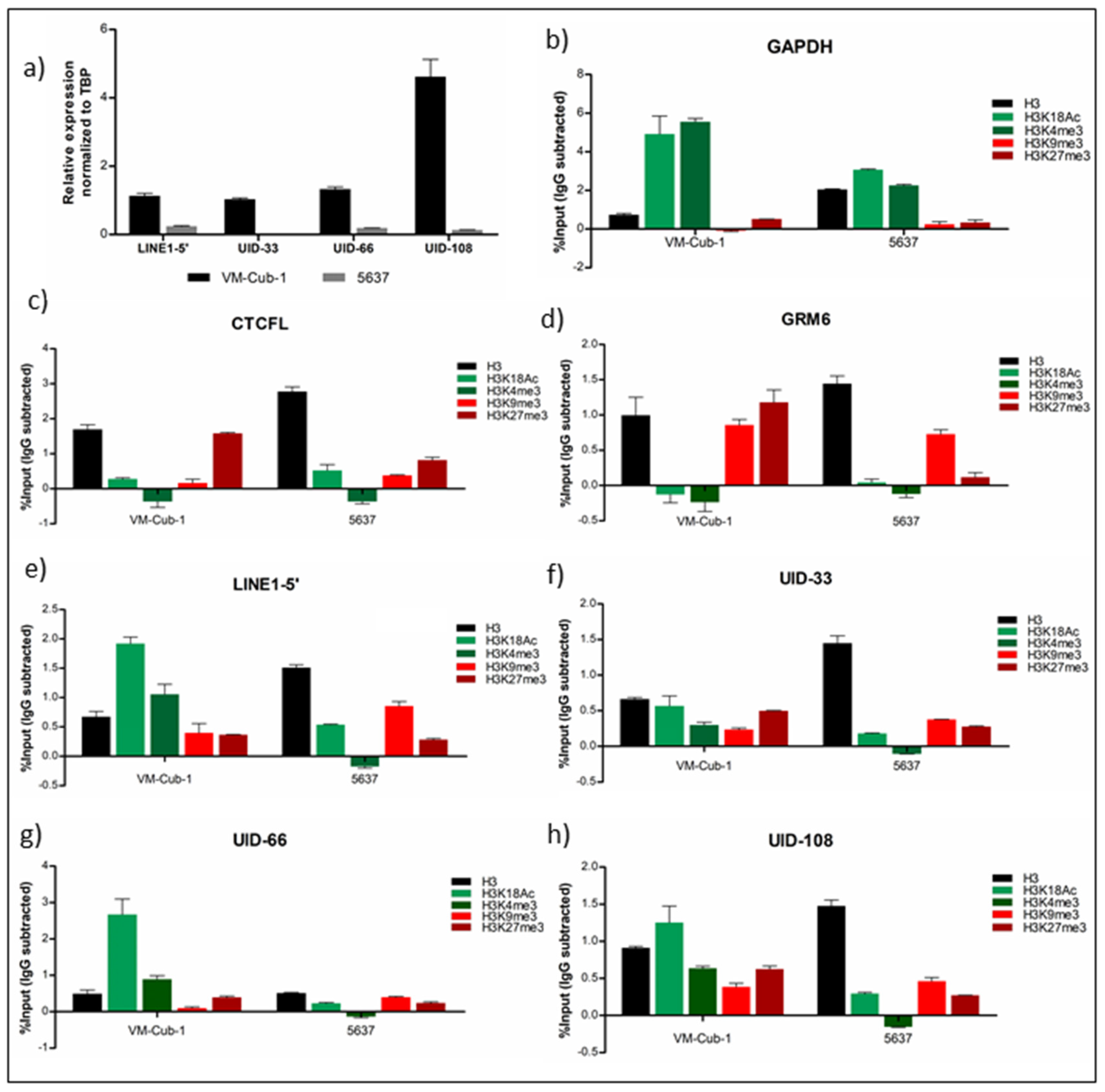
2.3. Analysis of Histone Modifications at Individual L1s
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Cell Lines and Cell Culture
4.3. RNA Immunoprecipitation via ORF1p
4.4. Nanopore Sequencing and Data Evaluation
4.5. RNA Extraction and Reverse Transcription
4.6. Quantitative Reverse Transcription PCR (qRT-PCR)
4.7. Chromatin Immunoprecipitation
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ChIP | Chromatin immunoprecipitation |
| flL1 | Full-le Ngth LINE-1 |
| L1 | LINE-1, long interspersed element 1 |
| ORF | Open reading frame |
| PBS | Phosphate-buffered saline |
| qRT-PCR | Quantitative reverse transcription polymerase chain reaction |
| RIP | RNA immunoprecipitation |
| UC | Urothelial carcinoma |
References
- Burns, K.H. Transposable elements in cancer. Nat. Rev. Cancer 2017, 17, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Goodier, J.L.; Kazazian, H.H., Jr. Retrotransposons Revisited: The Restraint and Rehabilitation of Parasites. Cell 2008, 135, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Lee, C.H.; Swigut, T.; Grow, E.; Gu, B.; Bassik, M.C.; Wysocka, J. Selective silencing of euchromatic L1s revealed by genome-wide screens for L1 regulators. Nature 2018, 553, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Jönsson, M.E.; Brattås, P.L.; Gustafsson, C.; Petri, R.; Yudovich, D.; Pircs, K.; Verschuere, S.; Madsen, S.; Hansson, J.; Larsson, J.; et al. Activation of neuronal genes via LINE-1 elements upon global DNA demethylation in human neural progenitors. Nat. Commun. 2019, 10, 3182. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Luque, F.J.; Kempen, M.-J.H.; Gerdes, P.; Vargas-Landin, D.B.; Richardson, S.R.; Troskie, R.-L.; Jesuadian, J.S.; Cheetham, S.W.; Carreira, P.E.; Salvador-Palomeque, C.; et al. LINE-1 Evasion of Epigenetic Repression in Humans. Mol. Cell 2019, 75, 590–604.e12. [Google Scholar] [CrossRef]
- Tubio, J.M.C.; Li, Y.; Ju, Y.S.; Martincorena, I.; Cooke, S.L.; Tojo, M.; Gundem, G.; Pipinikas, C.P.; Zamora, J.; Raine, K.; et al. Mobile DNA in cancer. Extensive transduction of nonrepetitive DNA mediated by L1 retrotransposition in cancer genomes. Science 2014, 345, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Chénais, B. Transposable Elements in Cancer and Other Human Diseases. Curr. Cancer Drug Targets 2015, 15, 227–242. [Google Scholar] [CrossRef]
- Ardeljan, D.; Taylor, M.S.; Ting, D.T.; Burns, K.H. The Human Long Interspersed Element-1 Retrotransposon: An Emerging Biomarker of Neoplasia. Clin. Chem. 2017, 63, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, M. A role of LINE-1 in telomere regulation. Front. Biosci. 2018, 23, 1310–1319. [Google Scholar] [CrossRef] [Green Version]
- Rangwala, S.H.; Zhang, L., Jr.; Many, H.H.K. LINE1 elements contribute to the transcriptome of human somatic cells. Genome Biol. 2009, 10, R100. [Google Scholar] [CrossRef] [Green Version]
- Philippe, C.; Vargas-Landin, D.B.; Doucet, A.J.; Van Essen, D.; Vera-Otarola, J.; Kuciak, M.; Corbin, A.; Nigumann, P.; Cristofari, G. Activation of individual L1 retrotransposon instances is restricted to cell-type dependent permissive loci. eLife 2016, 5, e13926. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, E.; Cajuso, T.; Katainen, R.; Kaasinen, E.; Välimäki, N.; Palin, K.; Taipale, J.; Aaltonen, L.A.; Kilpivaara, O. Frequent L1 retrotranspositions originating fromTTC28in colorectal cancer. Oncotarget 2014, 5, 853–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Martin, B.; Alvarez, E.G.; Baez-Ortega, A.; Zamora, J.; Supek, F.; Demeulemeester, J.; Santamarina, M.; Ju, Y.S.; Temes, J.; Garcia-Souto, D.; et al. Pan-cancer analysis of whole genomes identifies driver rearrangements promoted by LINE-1 retrotransposition. Nat. Genet. 2020, 52, 306–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.S.; Lacava, J.; Mita, P.; Molloy, K.R.; Huang, C.R.L.; Li, N.; Adney, E.M.; Jiang, H.; Burns, K.H.; Chait, B.T.; et al. Affinity proteomics reveals human host factors implicated in discrete stages of LINE-1 retrotransposition. Cell 2013, 155, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Deininger, P.; Morales, M.E.; White, T.B.; Baddoo, M.; Hedges, D.J.; Servant, G.; Srivastav, S.; Smither, M.E.; Concha, M.; Deharo, D.L.; et al. A comprehensive approach to expression of L1 loci. Nucleic Acids Res. 2017, 45, e31. [Google Scholar] [CrossRef]
- Ekreimer, U.; Schulz, W.A.; Ekoch, A.; Niegisch, G.; Egoering, W. HERV-K and LINE-1 DNA Methylation and Reexpression in Urothelial Carcinoma. Front. Oncol. 2013, 3. [Google Scholar] [CrossRef] [Green Version]
- Nüsgen, N.; Goering, W.; Dauksa, A.; Biswas, A.; Jamil, M.A.; Dimitriou, I.; Sharma, A.; Singer, H.; Fimmers, R.; Fröhlich, H.; et al. Inter-locus as well as intra-locus heterogeneity in LINE-1 promoter methylation in common human cancers suggests selective demethylation pressure at specific CpGs. Clin. Epigenetics 2015, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, A.A.J.; Kreimer, U.; Schulz, W.A.; Krikoni, A.; Schumann, G.G.; Häussinger, D.; Münk, C.; Egoering, W. APOBEC3B Activity Is Prevalent in Urothelial Carcinoma Cells and Only Slightly Affected by LINE-1 Expression. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Whongsiri, P.; Pimratana, C.; Wijitsettakul, U.; Sanpavat, A.; Jindatip, D.; Hoffmann, M.J.; Goering, W.; Schulz, W.A.; Boonla, C. Oxidative stress and LINE-1 reactivation in bladder cancer are epigenetically linked through active chromatin formation. Free. Radic. Biol. Med. 2019, 134, 419–428. [Google Scholar] [CrossRef]
- Rodić, N.; Sharma, R.; Sharma, R.; Zampella, J.; Dai, L.; Taylor, M.S.; Hruban, R.H.; Iacobuzio-Donahue, C.A.; Maitra, A.; Torbenson, M.S.; et al. Long Interspersed Element-1 Protein Expression Is a Hallmark of Many Human Cancers. Am. J. Pathol. 2014, 184, 1280–1286. [Google Scholar] [CrossRef] [Green Version]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodier, J.L.; Zhang, L.; Vetter, M.R.; Kazazian, H.H. LINE-1 ORF1 Protein Localizes in Stress Granules with Other RNA-Binding Proteins, Including Components of RNA Interference RNA-Induced Silencing Complex. Mol. Cell. Biol. 2007, 27, 6469–6483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodier, J.L.; Cheung, L.E.; Kazazian, J.H.H. Mapping the LINE1 ORF1 protein interactome reveals associated inhibitors of human retrotransposition. Nucleic Acids Res. 2013, 41, 7401–7419. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Li, J.; Xu, F.; Mei, S.; Le Duff, Y.; Yin, L.; Pang, X.; Cen, S.; Jin, Q.; Liang, C.; et al. SAMHD1 Inhibits LINE-1 Retrotransposition by Promoting Stress Granule Formation. PLoS Genet. 2015, 11, e1005367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubstenberger, A.; Courel, M.; Bénard, M.; Souquere, S.; Ernoult-Lange, M.; Chouaib, R.; Yi, Z.; Morlot, J.-B.; Munier, A.; Fradet, M.; et al. P-Body Purification Reveals the Condensation of Repressed mRNA Regulons. Mol. Cell 2017, 68, 144–157.e5. [Google Scholar] [CrossRef] [Green Version]
- Khong, A.; Matheny, T.; Jain, S.; Mitchell, S.F.; Wheeler, J.R.; Parker, R. The Stress Granule Transcriptome Reveals Principles of mRNA Accumulation in Stress Granules. Mol Cell 2017, 68, 808–820.e5. [Google Scholar] [CrossRef]
- Goering, W.; Ribarska, T.; Schulz, W.A. Selective changes of retroelement expression in human prostate cancer. Carcinogenesis 2011, 32, 1484–1492. [Google Scholar] [CrossRef]
- Hoffmann, M.J.; Müller, M.; Engers, R.; Schulz, W.A. Epigenetic control of CTCFL/BORIS and OCT4 expression in urogenital malignancies. Biochem. Pharmacol. 2006, 72, 1577–1588. [Google Scholar] [CrossRef]
- Knowles, M.A.; Hurst, C.D. Molecular biology of bladder cancer: New insights into pathogenesis and clinical diversity. Nat. Rev. Cancer 2015, 15, 25–41. [Google Scholar] [CrossRef]
- Ardeljan, D.; Steranka, J.P.; Liu, C.; Li, Z.; Taylor, M.S.; Payer, L.M.; Gorbounov, M.; Sarnecki, J.S.; Deshpande, V.; Hruban, R.H.; et al. Cell fitness screens reveal a conflict between LINE-1 retrotransposition and DNA replication. Nat. Struct. Mol. Biol. 2020, 27, 168–178. [Google Scholar] [CrossRef]
- Mita, P.; Sun, X.; Fenyö, D.; Kahler, D.J.; Li, D.; Agmon, N.; Wudzinska, A.; Keegan, S.; Bader, J.S.; Yun, C.; et al. BRCA1 and S phase DNA repair pathways restrict LINE-1 retrotransposition in human cells. Nat. Struct. Mol. Biol. 2020, 27, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, D.; Williamson, M.; Doherty, J. A Sensitive RNase Protection Assay to Detect Transcripts from Potentially Functional Human Endogenous L1 Retrotransposons. Biochem. Biophys. Res. Commun. 1996, 222, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Belancio, V.P.; Roy-Engel, A.M.; Pochampally, R.R.; Deininger, P. Somatic expression of LINE-1 elements in human tissues. Nucleic Acids Res. 2010, 38, 3909–3922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briggs, E.M.; Ha, S.; Mita, P.; Brittingham, G.; Sciamanna, I.; Spadafora, C.; Logan, S.K. Long interspersed nuclear element-1 expression and retrotransposition in prostate cancer cells. Mob. DNA 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erichsen, L.; Seifert, H.-H.; Schulz, W.A.; Hoffmann, M.J.; Niegisch, G.; Arauzo-Bravo, M.J.; Bendhack, M.L.; Poyet, C.; Hermanns, T.; Beermann, A.; et al. Basic Hallmarks of Urothelial Cancer Unleashed in Primary Uroepithelium by Interference with the Epigenetic Master Regulator ODC1. Sci. Rep. 2020, 10, 3808–3810. [Google Scholar] [CrossRef]
- Su, S.-F.; Abreu, A.L.D.C.; Chihara, Y.; Tsai, Y.; Andreu-Vieyra, C.; Daneshmand, S.; Skinner, E.C.; Jones, P.A.; Siegmund, K.; Liang, G. A Panel of Three Markers Hyper- and Hypomethylated in Urine Sediments Accurately Predicts Bladder Cancer Recurrence. Clin. Cancer Res. 2014, 20, 1978–1989. [Google Scholar] [CrossRef] [Green Version]
- Vafadar-Isfahani, N.; Parr, C.; McMillan, L.E.; Sanner, J.; Yeo, Z.; Saddington, S.; Peacock, O.; Cruickshanks, H.A.; Meehan, R.R.; Lund, J.N.; et al. Decoupling of DNA methylation and activity of intergenic LINE-1 promoters in colorectal cancer. Epigenetics 2017, 12, 465–475. [Google Scholar] [CrossRef]
- Seifert, H.-H.; Meyer, A.; Cronauer, M.V.; Hatina, J.; Müller, M.; Rieder, H.; Hoffmann, M.J.; Ackermann, R.; Schulz, W.A. A new and reliable culture system for superficial low-grade urothelial carcinoma of the bladder. World J. Urol. 2007, 25, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Koutsogiannouli, E.; Skowron, M.A.; Pinkerneil, M.; Niegisch, G.; Brandt, A.; Stepanow, S.; Rieder, H.; Schulz, W.A. The New Immortalized Uroepithelial Cell Line HBLAK Contains Defined Genetic Aberrations Typical of Early Stage Urothelial Tumors. Bl. Cancer 2016, 2, 449–463. [Google Scholar] [CrossRef] [Green Version]
- Swiatkowski, S. Activities of MAP-Kinase Pathways in Normal Uroepithelial Cells and Urothelial Carcinoma Cell Lines. Exp. Cell Res. 2003, 282, 48–57. [Google Scholar] [CrossRef]
- Moran, V.A.; Niland, C.N.; Khalil, A.M. Co-Immunoprecipitation of Long Noncoding RNAs. Adv. Struct. Safe. Stud. 2012, 925, 219–228. [Google Scholar] [CrossRef]
- Hoffmann, M.J.; Dehn, J.; Droop, J.; Niegisch, G.; Niedworok, C.; Szarvas, T.; Schulz, W.A. Truncated Isoforms of lncRNA ANRIL Are Overexpressed in Bladder Cancer, But Do Not Contribute to Repression of INK4 Tumor Suppressors. Non-Coding RNA 2015, 1, 266–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penzkofer, T.; Dandekar, T.; Zemojtel, T. L1Base: From functional annotation to prediction of active LINE-1 elements. Nucleic Acids Res. 2004, 33, D498–D500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhead, B.L.; Karolchik, D.; Kuhn, R.M.; Hinrichs, A.S.; Zweig, A.S.; Fujita, P.A.; Diekhans, M.; Smith, K.E.; Rosenbloom, K.R.; Raney, B.J.; et al. The UCSC Genome Browser database: Update 2010. Nucleic Acids Res. 2010, 38, D613–D619. [Google Scholar] [CrossRef]
- Fleige, S.; Walf, V.; Huch, S.; Prgomet, C.; Sehm, J.; Pfaffl, M.W. Comparison of relative mRNA quantification models and the impact of RNA integrity in quantitative real-time RT-PCR. Biotechnol. Lett. 2006, 28, 1601–1613. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L1 ID | Rank Exp.1 | Expression Exp.1 | Rank Exp.2 | Expression Exp2. | Localization Chromosome | Localization Bases | Strand | Closest Gene |
|---|---|---|---|---|---|---|---|---|
| UID-108 | 1 | 69.06 | 4 | 23.90 | chr1p31.1 | 71,887,202–71,895,252 | + | NEGR1 |
| UID-66 | 2 | 43.33 | 1 | 30.76 | chr12q14.2 | 64,194,585–64,202,633 | + | C12ORF66 |
| UID-60 | 3 | 39.94 | 3 | 28.24 | chrXp21.1 | 36,464,177–36,472,219 | - | Desert |
| UID-33 | 4 | 36.08 | 5 | 22.58 | ch17p13.1 | 9,614,984–9,623,031 | + | CFAP52 |
| UID-103 | 5 | 24.15 | 6 | 21.15 | chr1q25.1 | 174,376,771–174,384,818 | - | RABGAP1L |
| UID-140 | 6 | 20.16 | 8 | 12.48 | chr14q12 | 26,628,254–26,636,301 | - | NOVA1-AS1 |
| UID-93 | 7 | 19.25 | 2 | 28.40 | chr3q23 | 141,756,134–141,764,155 | - | RNF7/GRK7 |
| UID-59 | 8 | 16.38 | 7 | 18.01 | chrXp11.3 | 47,782,657–47,790,701 | - | WASF4P |
| UID-135 | 9 | 13.70 | 11 | 5.94 | chr22q12.1 | 28,662,282–28,670,329 | + | TTC28 |
| UID-92 | 10 | 10.74 | 10 | 7.00 | chr3q25.1 | 159,102,396–159,094,350 | - | IQCJ-SCHIF1 |
| UID-19 | 11 | 9.74 | 9 | 11.18 | chr16p12.3 | 21,049,706–21,041,661 | - | DNAH3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whongsiri, P.; Goering, W.; Lautwein, T.; Hader, C.; Niegisch, G.; Köhrer, K.; Hoffmann, M.J.; Schulz, W.A. Many Different LINE-1 Retroelements Are Activated in Bladder Cancer. Int. J. Mol. Sci. 2020, 21, 9433. https://doi.org/10.3390/ijms21249433
Whongsiri P, Goering W, Lautwein T, Hader C, Niegisch G, Köhrer K, Hoffmann MJ, Schulz WA. Many Different LINE-1 Retroelements Are Activated in Bladder Cancer. International Journal of Molecular Sciences. 2020; 21(24):9433. https://doi.org/10.3390/ijms21249433
Chicago/Turabian StyleWhongsiri, Patcharawalai, Wolfgang Goering, Tobias Lautwein, Christiane Hader, Günter Niegisch, Karl Köhrer, Michèle J. Hoffmann, and Wolfgang A. Schulz. 2020. "Many Different LINE-1 Retroelements Are Activated in Bladder Cancer" International Journal of Molecular Sciences 21, no. 24: 9433. https://doi.org/10.3390/ijms21249433
APA StyleWhongsiri, P., Goering, W., Lautwein, T., Hader, C., Niegisch, G., Köhrer, K., Hoffmann, M. J., & Schulz, W. A. (2020). Many Different LINE-1 Retroelements Are Activated in Bladder Cancer. International Journal of Molecular Sciences, 21(24), 9433. https://doi.org/10.3390/ijms21249433