1. Introduction
Breast cancer (Online Mendelian Inheritance in Man, OMIM no. 114480) is the most common cancer in women in the world [
1]. One of the frequently studied reasons for the lack of successful treatment outcomes in a considerable portion of the patients is multidrug resistance [
2]. Multidrug resistance can be caused by ATP-binding cassette (ABC) transporters, e.g., by higher efflux of drugs out of tumor cells by P-glycoprotein (Multidrug resistance, MDR coded by
ABCB1 gene) [
2,
3].
ABC transporters represent a large superfamily of membrane transporter proteins classified into seven families and translocate numerous compounds across intra and extracellular membranes. Their substrates include metabolic products, sterols, lipids, and xenobiotics [
3]. However, of the total number of 48 active human ABC transporters, up to only 16 are able to transport anticancer drugs [
3]. Most of the drug resistance is ascribed to largely studied multidrug resistance-related transporters
ABCB1 (MDR, OMIM no. 171050),
ABCC1 (MRP, OMIM no. 158343), and
ABCG2 (BCRP, OMIM no. 603756) [
4]. Proteins of the family ABCA are mostly lipid sterol transporters and can be associated with several diseases, e.g., Tangier or Alzheimer disease [
5]. Their roles in cancer progression and the metastatic potential linked to lipid trafficking recently became the focus of numerous studies [
6]. ABCBs participate in antigen processing and immune deficiency. Apart from ABCB1, a promiscuous and ubiquitous efflux pump [
2], ABCB members also represent transporters of heme and bile acids [
7]. Family C is mostly dedicated to multidrug resistance (MRP1-6) [
8,
9], but ABCC6, ABCC7, and ABCC8/9 are linked to serious diseases (pseudoxanthoma elasticum, cystic fibrosis, and diabetes mellitus, respectively) [
9]. ABCDs are responsible for transport of fatty acids from peroxisomes to the cytoplasm [
10] and ABCGs transport various products of metabolism, xenobiotics including anti-cancer drugs, bile acids, and steroids [
11]. The rest of the transporters are not involved in transport, but rather act as translational inhibitors or protein synthesis regulators (ABCFs and ABCEs) [
12].
Our recent pharmacogenomics study revealed a prognostic and predictive potential in a number of alterations in breast cancer [
13]. The studied genes were implicated in the metabolism and transport of drugs administered to breast cancer patients in the clinics, clearly documenting the importance of this tool for the personalized medicine. The study provided a proof-of-the principle for the design and bioinformatics methodology, namely, the assembly and testing of an in-house pipeline for variant prioritization. Given the total number of 509 genes screened by the next generation sequencing (NGS), only a portion of variants could be validated in a subsequent phase. In order to select the most relevant functional alterations from the statistically significant set of variants, we down-sampled the results using information from in silico predictions and according to previously confirmed pharmacogenomic and clinical evidence. Thus, some potentially useful biomarkers of prognosis or prediction of therapy outcome could have been missed.
In the present study, we aimed to use less strict criteria for investigating the importance of germline genetic variability in coding, untranslated regions (UTR), and adjacent regions of all human members of the ABC superfamily for prognosis and response to cytotoxic therapy of breast cancer patients. All variants in ABCs identified in the testing phase were correlated with disease-free survival (DFS) and response of the patients to preoperative cytotoxic therapy, and thoroughly reviewed, including permutation analysis, evaluation of haplotypes, and gene dosage. We have not addressed functional relevance to enable identification of purely correlative biomarkers. Prioritized variants were further validated in a large cohort of breast cancer patients from a single population. Thus, the present study brings a more detailed view of the relevance of genetic variability of ABC transporters for breast cancer prognosis and therapy outcome predictions.
3. Discussion
The role of germline genetic variability among ABC transporters in prognosis of breast cancer patients as well as in their response to chemotherapy is underexplored. In our previous publication, we dealt with pharmacologically relevant germline genetic polymorphisms in 509 breast cancer-related genes [
13]. In the present study, we used the same approach to reveal all associations of genetic variants in human ABC transporters with chemotherapy response and survival of the patients.
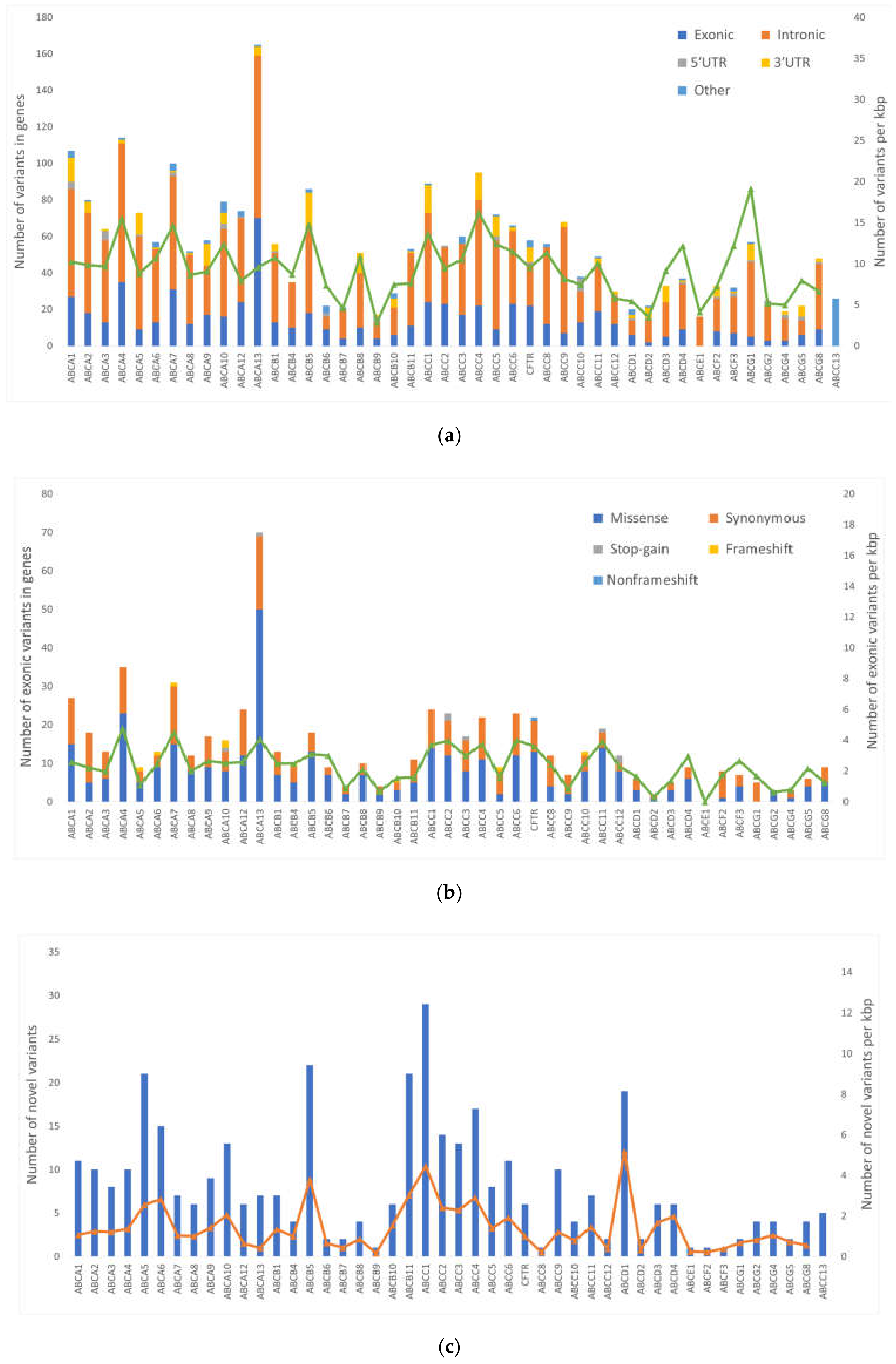
A total of 2611 variants were found in a testing set. The majority of variants were found in intronic regions. Lower numbers of variants were found in coding regions and UTRs. Interestingly, no variants were found in
ABCF1,
TAP1 (alias
ABCB2), and
TAP2 (
ABCB3).
TAP1 and
TAP2 are antigen presenting transporters and alterations in these genes associate with autoimmune diseases, susceptibility to infections, or malignancies [
15]. Similarly,
ABCF1 plays a role in the regulation of inflammatory processes [
16] and alterations in
ABCF1 are linked with autoimmune diseases as well [
17]. Therefore, it seems that genetic variants in these genes negatively impacts immunity and inflammatory processes which explain limited variability, in line with our findings. On the other hand, the most variable genes were
ABCA13 (165 alterations),
ABCA4 (114), and
ABCA1 (109). The members of ABCA family are typically large genes (transcript length 7‒17 kbp) and thus likely to accumulate variants. When normalized for the length of transcript,
ABCG1, ABCC4, and
ABCA4 have the highest count of variants per kbp, ranging from 16 to 20. Interestingly,
ABCA4, ABCA7, and
ABCA13 had the highest variant counts in exonic regions (4.1‒4.8 variants per kbp). We found several LOF variants in ABC transporters—eight stop-gains and nine frameshift indels. These events have high impact on function of the protein. Moreover, all 17 LOF variants were present in genes of the first quartile of the most intolerant genes to LOF events [
18]. These facts advocate for further investigation of LOF variants in ABC transporters. Unfortunately, LOF variants are rare. For the sake of maintaining enough statistical power for comparison with clinical data, only common variants (MAF > 0.05) could be used in the present study.
In total, we selected 903 variants and subjected them to a thorough statistical analyses. Of these variants, 43 associated with response or DFS and were capable of validation in a cohort of 802 breast cancer patients. Five associations with DFS and nine with response to NACT were replicated in the validation set. If multiple simultaneous statistical tests are calculated, a type I error (a risk of false positive results) occurs. To prevent this error, correction for multiple testing must be used. There are several methods to do so. Here, we applied the wildly used FDR, a test by Benjamini–Hochberg. After this correction, none of the associations of variants with clinical features remained significant and, thus, cannot be considered validated. Nevertheless, we found some interesting associations which we will discuss further.
ABCA13 is responsible for lipid transport and variants in this gene can cause schizophrenia [
5]. Carriage of the rare allele of SNP rs17548783, located in downstream intronic region of
ABCA13, was associated with shorter DFS of patients in our study. Based on our findings, a rare allele of this variant, significantly associated with lower
ABCA13 intratumoral transcript levels in a validation set (
Table 6). Lower transcript levels of
ABCA13 were associated with worse response to NACT in a previous study [
14], further underpinning the role of this SNP as a putative poor prognosis biomarker. This consequence is the most interesting link observed at present. Nevertheless, the response to NACT does not significantly associate with DFS in our datasets a fact that clearly calls for further research.
Another variant in
ABCA13, the missense rs74859514 (Ala2223Pro), associated with DFS in patients treated with chemotherapy, but without gene dosage relationship. Neither of these two SNPs has records in the present literature, although associations of
ABCA13 with patients’ outcome have been described in several previous studies. A decreased expression of
ABCA13 was associated with shorter DFS in 51 glioblastoma patients [
19] and 51 colorectal cancer patients [
20]. The opposite was found for ovarian cancer (
n = 77) and higher levels of
ABCA13 predicted worse overall survival in ovarian cancer patients [
21]. Amplification of 7p12 (which includes
ABCA13 and
HUS1, EGFR, and
IKZF1) predicted worse response to NACT in muscle-invasive bladder cancer [
22]. Such contradictory results from different cancers are puzzling. Despite we must bear in mind that none of the associations found in our study passed the FDR test, some may still have clinical potential. Additional studies will be needed to confirm these results.
A synonymous variant rs71428357 in
ABCA12 associated with response to NACT. Patients responding well to chemotherapy were more often carriers of the rare allele. Synonymous variants can affect RNA splicing, folding, and stability [
23] and are associated with several diseases, such as Alzheimer disease, pulmonary sarcoidosis, galactosemia, or cancer [
24]. The role of this particular
ABCA12 variant in cancer or other diseases is still unknown. However, higher
ABCA12 transcript levels in non-tumor tissues associated with worse response to NACT in breast cancer patients in our previous study [
14]. The opposite, i.e., higher levels associating with residual disease, was found by Park et al. [
25]. Interestingly, we previously identified this gene among candidate ABCs with predictive or prognostic potential for patients with breast, colorectal, and pancreatic carcinomas [
26].
Among other members of the ABCA family, associations with response to NACT were observed for
ABCA4 (variant rs2275032) and
ABCA9 (rs11871944). A deletion in
ABCA7 (rs9282562) associated with shorter DFS of the patients. These variants are not described in the present literature, however, higher transcript levels of
ABCA9 significantly associated with worse survival in high-grade serous ovarian cancer tumors [
6]. Silencing of
ABCA7 reduces epithelial to mesenchymal transition in ovarian cancer cell lines and knockdown of
ABCA7 inhibited migration, cell proliferation, and invasion [
27]. In addition, lower
ABCA7 levels associated with shorter DFS of colorectal cancer patients [
20].
ABCB5 confers 5-fluorouracil resistance and promotes cell invasiveness in colorectal cancer [
28]. Variant rs3210441 in
ABCB5 associated with response to NACT in our study, but no eQTL was found and additional supportive data about the role of this SNP or protein in breast cancer is lacking.
Protein coded by
ABCC11 is responsible for transport of bile acids, conjugated steroids, or cyclic nucleotides. Diseases linked with this gene include malfunction of apocrine gland secretion and lateral sinus thrombosis (
https://www.genecards.org).
ABCC11 is a transporter of 5-fluorouracil [
3]. In our study, a missense
ABCC11 variant rs17822931 (Gly180Arg) associated with response to NACT. Carriers of the wild-type allele had significantly poorer outcomes than patients with an alternative allele. This variant is known for its determination of human earwax type [
29]. It is associated with breast cancer risk in the Japanese population [
30]. This variant is also linked with axillary osmidrosis, colostrum secretion in the mammary gland, and mastopathies [
31]. Wild type allele C also confers to chemotherapy resistance to 5-fluorouracil by exporting active metabolite 5-fluoro-2′-deoxyuridine 5′-monophosphate (FdUMP) [
32].
ABCC11 expression (together with
ABCB1) is responsible for resistant phenotype of breast cancer cell lines resistant to eribulin and inhibition of
ABCC11 can partially restore the cross-resistance to 5-fluorouracil [
33]. Higher
ABCC11 gene expression was also associated with poor response to NACT in breast cancer patients [
25]. Interestingly, this SNP is associated with expression of
ABCC11 only in the brain, but with
LONP2, coding mitochondrial matrix protein, in breast tissue (
Table 7). Relations between mastopathy, breast cancer risk, and, after chemotherapy, even drug resistance suggest strong connection of this variant to breast cancer. Association with response to chemotherapy of breast cancer patients has been suggested previously [
31], our result corroborates this assertion.
Among other members of the ABCC family,
ABCC5 (rs4148579) and
ABCC8 (rs739689) associated with response to NACT and
ABCC4 (rs899494) with DFS of the patients.
ABCC4 was among amplified genes in resistant cancer cell lines [
34]. The
ABCC4 gene was also identified to play a role in cellular migration of breast cancer cell line models MCF-7 and MDA-MB-231 [
35]. In our previous study [
14], we have seen associations of high
ABCC8 transcript levels with low grade and negative/positive status of estrogen receptor. Additionally, the expression level non-significantly (
p = 0.096) associated with worse responses of breast cancer patients to NACT [
14]. Nevertheless, in the present study we did not find association of rs739689 (intronic A > G transition) with
ABCC8 transcript levels. eQTL associations at the GTEx portal are ambiguous. The wild-type AA genotype has the highest expression of
ABCC8 in cerebellum, but no significant association can be found in breast tissue. This SNP is also highly significantly associated with expression of
NCR3LG1,
KCNJ11, and
SNORD14 genes with fragmentary and elusive data on association with breast cancer. From the data discussed above, it can be summarized that the present knowledge is incomplete and, thus, no clear picture can be presented.
Unlike other ABCD transporters,
ABCD4 is not found in peroxisomes, but in lysosomes. It takes part in transport of cobalamin (vitamin B12) and mutations in this transporter cause inherited defects of intracellular cobalamin metabolism [
10]. Low transcript levels of this gene were also associated with shorter DFS of colorectal cancer patients [
20] and
ABCD4 was among amplified genes in resistant cancer cell lines [
34]. In our study, wild-type variants rs2301347 and rs2301346 (both intronic) associated with the good response to NACT. Wild-type genotypes of these two variants show lower transcript levels of long non-coding (lnc) RNA lnc-SYNDIG1L-2 overlapping
ABCD4 in mammary tissue (
Table 7) suggesting potential clinical relevance. However, the lack of association with
ABCD4 transcript levels that we found in our dataset precludes any strict conclusions.
ABCG8 is a transporter of sterols from hepatocytes and enterocytes [
36]. The rare allele of its SNP rs34198326 was associated with longer DFS of chemotherapy treated patients in our study. Expression of
ABCG8 was downregulated in tumors of breast cancer patients compared to non-neoplastic control tissues [
14], but the role of germline polymorphism is unclear.
The role of ABC transporters in cancer has been known for a long time. Multidrug resistance has been studied since 1970, when it was first mentioned [
37]. The well-studied
ABCB1 gene (MDR1) was discovered in 1974 by V. Ling, and nearly twenty years later, the discovery of
ABCC1 and
ABCG2 concerning drug resistance was reported [
2]. Although associations of
ABCB1 gene expression with breast cancer prognosis were reported repeatedly, evidence for the role of its genetic variability in response to treatment is elusive. A recent review demonstrated that three frequently studied polymorphisms in
ABCB1 (rs1045642, rs1128503, and rs2032582) cannot be considered reliable predictors of response to chemotherapy in breast cancer patients [
38]. Similarly, an association of
ABCC1 expression with the survival of breast cancer patients was described [
39]. However, only a few studies on genetic polymorphisms can be found.
ABCC1 variants rs4148350, rs45511401, and rs246221 associated with the risk of febrile neutropenia in patients treated with 5-fluorouracil, epirubicin, and cyclophosphamide [
40] and it was very recently discovered that
ABCC1 variant burden is a strong predictor of DFS in breast cancer patients rather than the genotype attributed to individual variants [
41].
ABCG2 transports several drugs used for breast cancer treatment. In a recent study on the TCGA cohort,
ABCG2 transcript levels associated with a decreased progression-free survival, although, gene variants (either somatic or germline) influenced
ABCG2 expression only moderately [
42]. From the above-reviewed information, it can be summarized that despite numerous studies on drug transporters utilization for predicting therapy outcome, strong support is still missing. Other transporters, with rather physiological roles, are much less explored in oncology, and studies were largely dedicated to gene expression in contrast with less studied genetic variability.
In conclusion, genetic variability in ABC transporters might play a role in breast cancer prognosis and help with prediction of therapy outcome of the patients. Although no alterations observed by this study can be considered statistically validated, particularly associations of downstream variant affecting expression, rs17548783 in ABCA13 with DFS and variant rs17822931 (Gly180Arg) in ABCC11 with response to NACT attract attention because of their support in the literature. These are interesting candidates for future research. Furthermore, elucidations are needed to explore additional genetic component, e.g., non-coding sequences, copy numbers and structural variations, somatic mutations, etc. of the ABC transporter superfamily.

 ,
, 



{kind=link}
{kind=link}