Hop Bitter Acids Increase Hippocampal Dopaminergic Activity in a Mouse Model of Social Defeat Stress



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
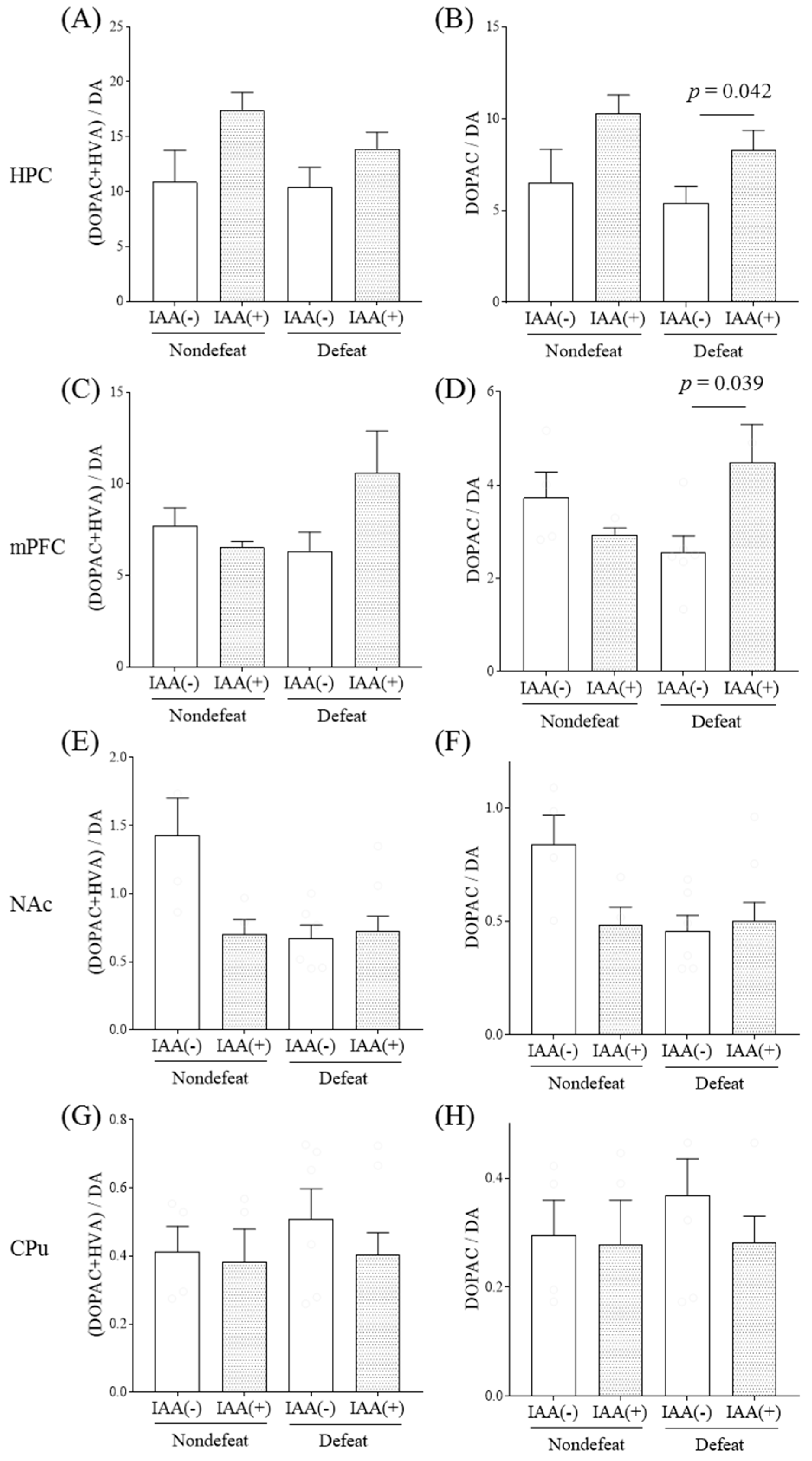
2.1. Effects of Hop Bitter Acids on Dopaminergic Activities in a Mouse Model of Social Defeat Stress
2.2. Effects of Hop Bitter Acids on Emotional Disturbance in a Mouse Model of Social Defeat Stress
3. Discussion
4. Materials and Methods
4.1. Preparation of IAA
4.2. Preparation of MHBA
4.3. Animals
4.4. Repeated SDS (R-SDS)
4.5. Social Interaction Test (SIT)
4.6. Monoamine Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CPu | Candate-putamen |
| DA | Dopamine |
| DOPAC | 3,4-dihydroxyphenylacetic acid |
| HVA | Homovanillic acid |
| IAA | Iso-α-acids |
| MHBA | Mature hop bitter acids |
| NAc | Nucleus accumbens |
| PFC | Prefrontal cortex |
| R-SDS | Repeated social defeat stress |
References
- Gu, Z.; Chu, L.; Han, Y. Therapeutic effect of resveratrol on mice with depression. Exp. Ther. Med. 2019, 17, 3061–3064. [Google Scholar] [CrossRef] [PubMed]
- Solanki, N.; Salvi, A.; Patki, G.; Salim, S. Modulating Oxidative Stress Relieves Stress-Induced Behavioral and Cognitive Impairments in Rats. Int. J. Neuropsychopharmacol. 2017, 20, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Ano, Y.; Hoshi, A.; Ayabe, T.; Ohya, R.; Uchida, S.; Yamada, K.; Kondo, K.; Kitaoka, S.; Furuyashiki, T. Iso-α-acids, the bitter components of beer, improve hippocampus-dependent memory through vagus nerve activation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 4987–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ano, Y.; Dohata, A.; Taniguchi, Y.; Hoshi, A.; Uchida, K.; Takashima, A.; Nakayama, H. Iso-alpha-acids, bitter components of beer, prevent inflammation and cognitive decline induced in a mouse model of alzheimer’s disease. J. Biol. Chem. 2017, 292, 3720–3728. [Google Scholar] [CrossRef] [Green Version]
- Ano, Y.; Ohya, R.; Kondo, K.; Nakayama, H. Iso-alpha-acids, hop-derived bitter components of beer, attenuate age-related inflammation and cognitive decline. Front. Aging Neurosci. 2019, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Ayabe, T.; Ohya, R.; Taniguchi, Y.; Shindo, K.; Kondo, K.; Ano, Y. Matured hop-derived bitter components in beer improve hippocampus-dependent memory through activation of the vagus nerve. Sci. Rep. 2018, 8, 15372. [Google Scholar] [CrossRef]
- Fukuda, T.; Obara, K.; Saito, J.; Umeda, S.; Ano, Y. Effects of hop bitter acids, bitter components in beer, on cognition in healthy adults: A randomized controlled trial. J. Agric. Food Chem. 2020, 68, 206–212. [Google Scholar] [CrossRef]
- Fukuda, T.; Ohnuma, T.; Obara, K.; Kondo, S.; Arai, H.; Ano, Y. Supplementation with matured hop bitter acids improves cognitive performance and mood state in healthy older adults with subjective cognitive decline. J. Alzheimer’s Dis. 2020, 76, 387–398. [Google Scholar] [CrossRef]
- Meyer, J.H.; Kruger, S.; Wilson, A.A.; Christensen, B.K.; Goulding, V.S.; Schaffer, A.; Minifie, C.; Houle, S.; Hussey, D.; Kennedy, S.H. Lower dopamine transporter binding potential in striatum during depression. Neuroreport 2001, 12, 4121–4125. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Hanke, M.L.; Corona, A.W.; Powell, N.D.; Stiner, L.M.; Bailey, M.T.; Nelson, R.J.; Godbout, J.P.; Sheridan, J.F. beta-Adrenergic receptor antagonism prevents anxiety-like behavior and microglial reactivity induced by repeated social defeat. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 6277–6288. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Furuyashiki, T.; Kitaoka, S.; Senzai, Y.; Imoto, Y.; Segi-Nishida, E.; Deguchi, Y.; Breyer, R.M.; Breyer, M.D.; Narumiya, S. Prostaglandin E2-mediated attenuation of mesocortical dopaminergic pathway is critical for susceptibility to repeated social defeat stress in mice. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 4319–4329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, M.; Ota, K.T.; Li, X.Y.; Sakaue, F.; Li, N.; Dutheil, S.; Banasr, M.; Duric, V.; Yamanashi, T.; Kaneko, K.; et al. Psychological Stress Activates the Inflammasome via Release of Adenosine Triphosphate and Stimulation of the Purinergic Type 2X7 Receptor. Biol. Psychiatry 2016, 80, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Kitaoka, S.; Tanaka, K.; Segi-Nishida, E.; Imoto, Y.; Ogawa, A.; Nakano, F.; Tomohiro, A.; Nakayama, K.; Taniguchi, M.; et al. The Innate Immune Receptors TLR2/4 Mediate Repeated Social Defeat Stress-Induced Social Avoidance through Prefrontal Microglial Activation. Neuron 2018, 99, 464–479.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menard, C.; Pfau, M.L.; Hodes, G.E.; Kana, V.; Wang, V.X.; Bouchard, S.; Takahashi, A.; Flanigan, M.E.; Aleyasin, H.; LeClair, K.B.; et al. Social stress induces neurovascular pathology promoting depression. Nat. Neurosci. 2017, 20, 1752–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKim, D.B.; Weber, M.D.; Niraula, A.; Sawicki, C.M.; Liu, X.; Jarrett, B.L.; Ramirez-Chan, K.; Wang, Y.; Roeth, R.M.; Sucaldito, A.D.; et al. Microglial recruitment of IL-1beta-producing monocytes to brain endothelium causes stress-induced anxiety. Mol. Psychiatry 2018, 23, 1421–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohleb, E.S.; Terwilliger, R.; Duman, C.H.; Duman, R.S. Stress-induced neuronal colony stimulating factor 1 provokes microglia-mediated neuronal remodeling and depressive-like behavior. Biol. Psychiatry 2018, 83, 38–49. [Google Scholar] [CrossRef]
- Shinohara, R.; Taniguchi, M.; Ehrlich, A.T.; Yokogawa, K.; Deguchi, Y.; Cherasse, Y.; Lazarus, M.; Urade, Y.; Ogawa, A.; Kitaoka, S.; et al. Dopamine D1 receptor subtype mediates acute stress-induced dendritic growth in excitatory neurons of the medial prefrontal cortex and contributes to suppression of stress susceptibility in mice. Mol. Psychiatry 2018, 23, 1717–1730. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, D.; Walsh, J.J.; Friedman, A.K.; Juarez, B.; Ku, S.M.; Koo, J.W.; Ferguson, D.; Tsai, H.C.; Pomeranz, L.; Christoffel, D.J.; et al. Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nature 2013, 493, 532–536. [Google Scholar] [CrossRef]
- Hare, B.D.; Shinohara, R.; Liu, R.J.; Pothula, S.; DiLeone, R.J.; Duman, R.S. Optogenetic stimulation of medial prefrontal cortex Drd1 neurons produces rapid and long-lasting antidepressant effects. Nat. Commun. 2019, 10, 223. [Google Scholar] [CrossRef]
- Yamazaki, T.; Takahashi, C.; Taniguchi, Y.; Narukawa, M.; Misaka, T.; Ano, Y. Bitter taste receptor activation by hop-derived bitter components induces gastrointestinal hormone production in enteroendocrine cells. Biochem. Biophys. Res. Commun. 2020, 533, 704–709. [Google Scholar] [CrossRef]
- Iñiguez, S.D.; Aubry, A.; Riggs, L.M.; Alipio, J.B.; Zanca, R.M.; Flores-Ramirez, F.J.; Hernandez, M.A.; Nieto, S.J.; Musheyev, D.; Serrano, P.A. Social defeat stress induces depression-like behavior and alters spine morphology in the hippocampus of adolescent male C57BL/6 mice. Neurobiol. Stress 2016, 5, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, S.; Miyake, Y.; Yoshimi, A.; Mouri, A.; Hida, H.; Yamada, K.; Ozaki, N.; Nabeshima, T.; Noda, Y. Dysfunction of Serotonergic and Dopaminergic Neuronal Systems in the Antidepressant-Resistant Impairment of Social Behaviors Induced by Social Defeat Stress Exposure as Juveniles. Int. J. Neuropsychopharmacol. 2018, 21, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Shuto, T.; Kuroiwa, M.; Sotogaku, N.; Kawahara, Y.; Oh, Y.S.; Jang, J.H.; Shin, C.H.; Ohnishi, Y.N.; Hanada, Y.; Miyakawa, T.; et al. Obligatory roles of dopamine D1 receptors in the dentate gyrus in antidepressant actions of a selective serotonin reuptake inhibitor, fluoxetine. Mol. Psychiatry 2020, 25, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Anacker, C.; Luna, V.M.; Stevens, G.S.; Millette, A.; Shores, R.; Jimenez, J.C.; Chen, B.; Hen, R. Hippocampal neurogenesis confers stress resilience by inhibiting the ventral dentate gyrus. Nature 2018, 559, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, Y.; Matsukura, Y.; Taniguchi, H.; Koizumi, H.; Katayama, M. Development of preparative and analytical methods of the hop bitter acid oxide fraction and chemical properties of its components. Biosci. Biotechnol. Biochem. 2015, 79, 1684–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ano, Y.; Ohya, R.; Yamazaki, T.; Takahashi, C.; Taniguchi, Y.; Kondo, K.; Takashima, A.; Uchida, K.; Nakayama, H. Hop bitter acids containing a β-carbonyl moiety prevent inflammation-induced cognitive decline via the vagus nerve and noradrenergic system. Sci. Rep. 2020, 10, 20028. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, R.; Rasool, A.; Ahmed, M.; Kaleem, U.; Duru, L.N.; Ma, H.; Deng, Y. Synergistic neuroprotective effect of endogenously-produced hydroxytyrosol and synaptic vesicle proteins on pheochromocytoma cell line against salsolinol. Molecule 2020, 25, 1715. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, Y.; Matsukura, Y.; Ozaki, H.; Nishimura, K.; Shindo, K. Identification and quantification of the oxidation products derived from alpha-acids and beta-acids during storage of hops (Humulus lupulus L.). J. Agric. Food Chem. 2013, 61, 3121–3130. [Google Scholar] [CrossRef]
- Higashida, S.; Nagai, H.; Nakayama, K.; Shinohara, R.; Taniguchi, M.; Nagai, M.; Hikida, T.; Yawata, S.; Ago, Y.; Kitaoka, S.; et al. Repeated social defeat stress impairs attentional set shifting irrespective of social avoidance and increases female preference associated with heightened anxiety. Sci. Rep. 2018, 8, 10454. [Google Scholar] [CrossRef]
- Ano, Y.; Ayabe, T.; Kutsukake, T.; Ohya, R.; Takaichi, Y.; Uchida, S.; Yamada, K.; Uchida, K.; Takashima, A.; Nakayama, H. Novel lactopeptides in fermented dairy products improve memory function and cognitive decline. Neurobiol. Aging 2018, 72, 23–31. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ano, Y.; Kitaoka, S.; Ohya, R.; Kondo, K.; Furuyashiki, T. Hop Bitter Acids Increase Hippocampal Dopaminergic Activity in a Mouse Model of Social Defeat Stress. Int. J. Mol. Sci. 2020, 21, 9612. https://doi.org/10.3390/ijms21249612
Ano Y, Kitaoka S, Ohya R, Kondo K, Furuyashiki T. Hop Bitter Acids Increase Hippocampal Dopaminergic Activity in a Mouse Model of Social Defeat Stress. International Journal of Molecular Sciences. 2020; 21(24):9612. https://doi.org/10.3390/ijms21249612
Chicago/Turabian StyleAno, Yasuhisa, Shiho Kitaoka, Rena Ohya, Keiji Kondo, and Tomoyuki Furuyashiki. 2020. "Hop Bitter Acids Increase Hippocampal Dopaminergic Activity in a Mouse Model of Social Defeat Stress" International Journal of Molecular Sciences 21, no. 24: 9612. https://doi.org/10.3390/ijms21249612
APA StyleAno, Y., Kitaoka, S., Ohya, R., Kondo, K., & Furuyashiki, T. (2020). Hop Bitter Acids Increase Hippocampal Dopaminergic Activity in a Mouse Model of Social Defeat Stress. International Journal of Molecular Sciences, 21(24), 9612. https://doi.org/10.3390/ijms21249612