Peptide–Peptide Co-Assembly: A Design Strategy for Functional Detection of C-peptide, A Biomarker of Diabetic Neuropathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
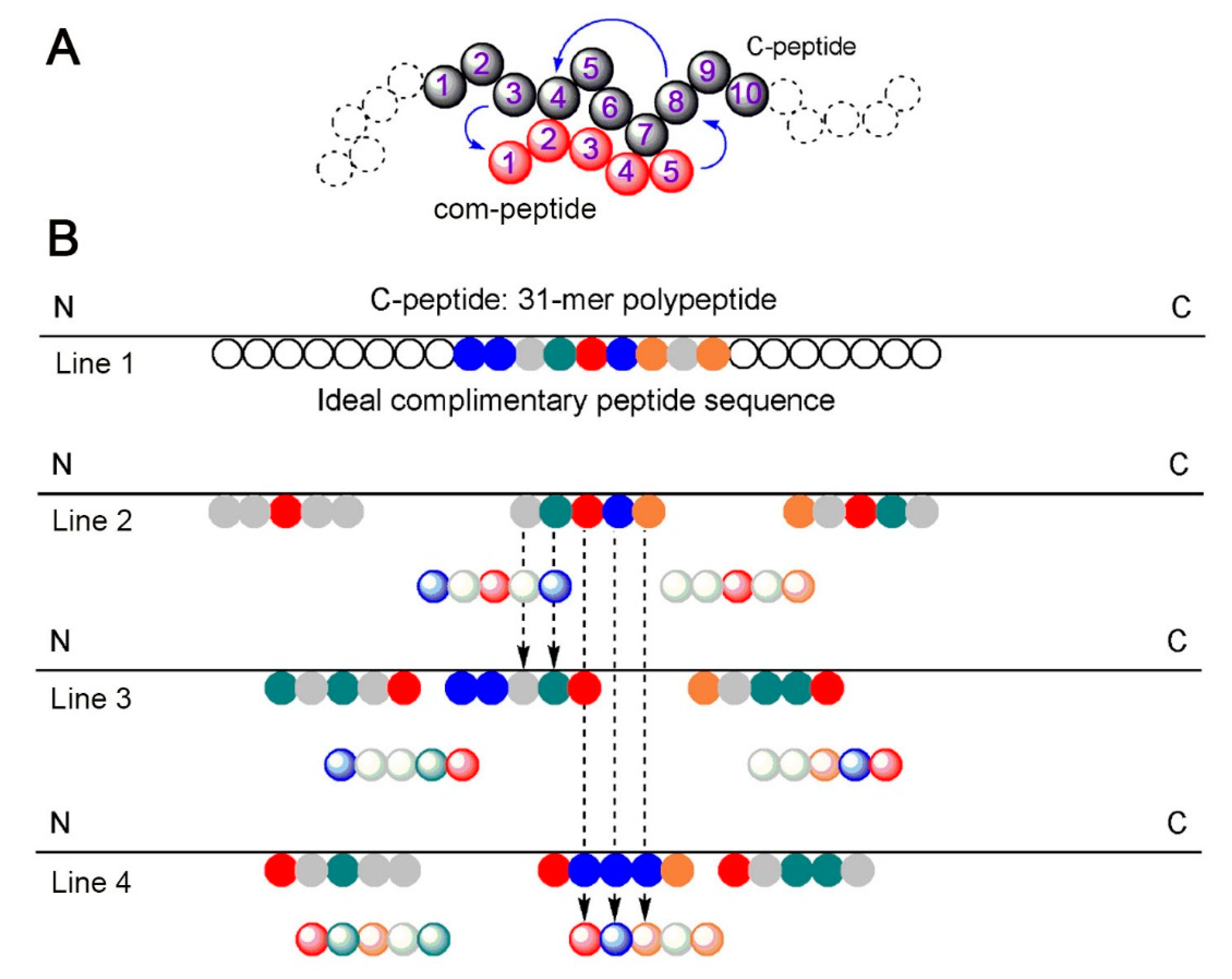
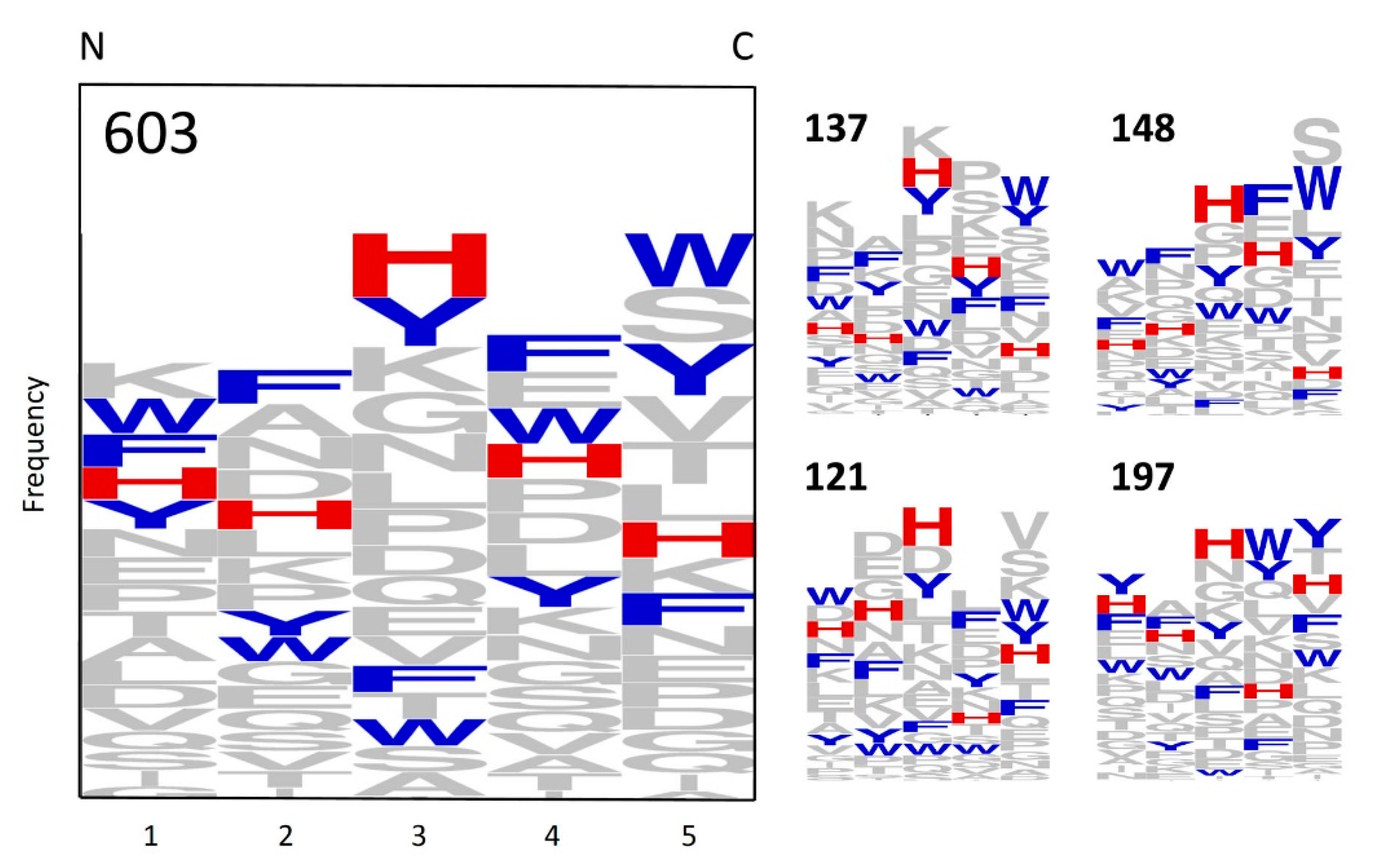
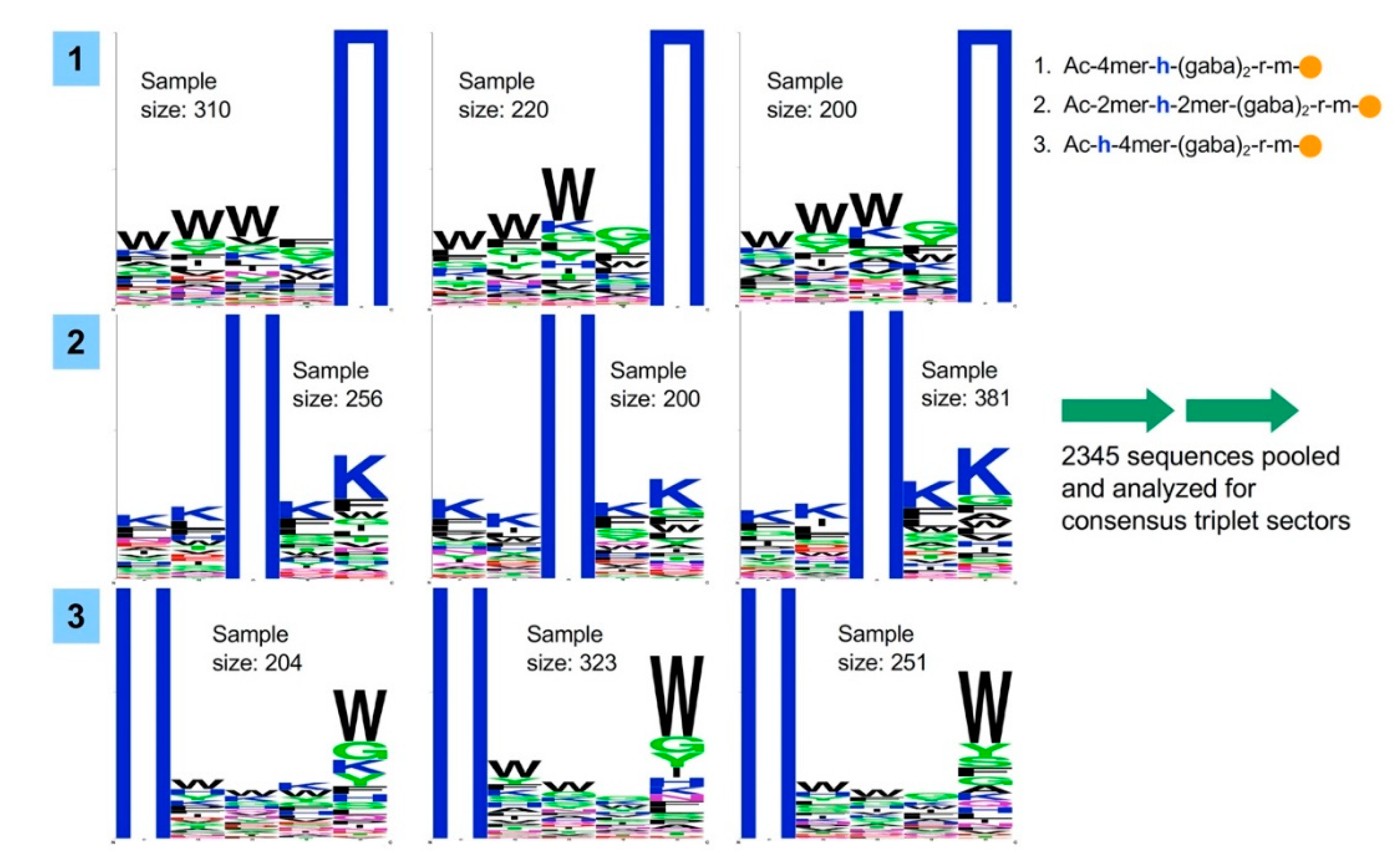
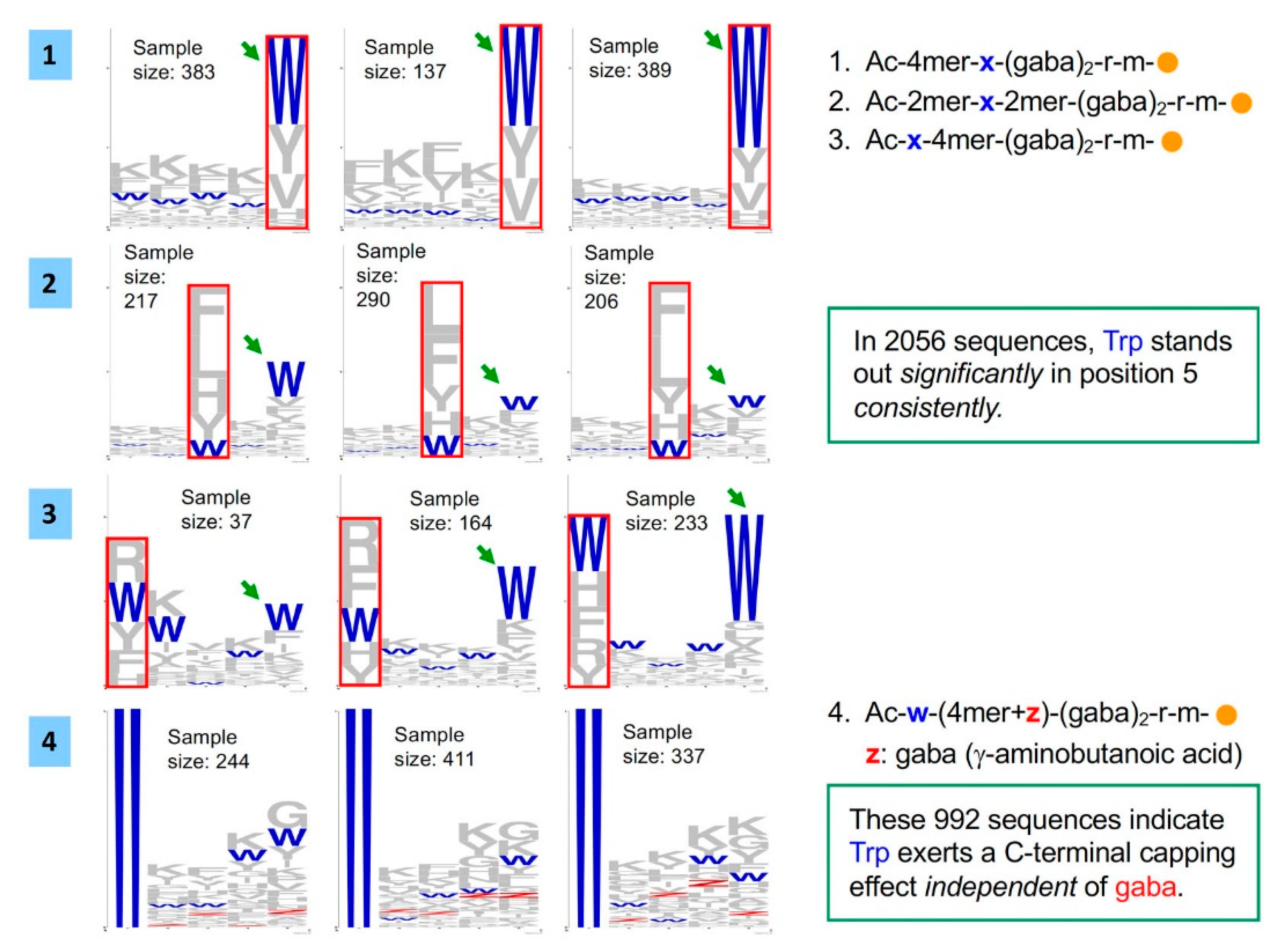
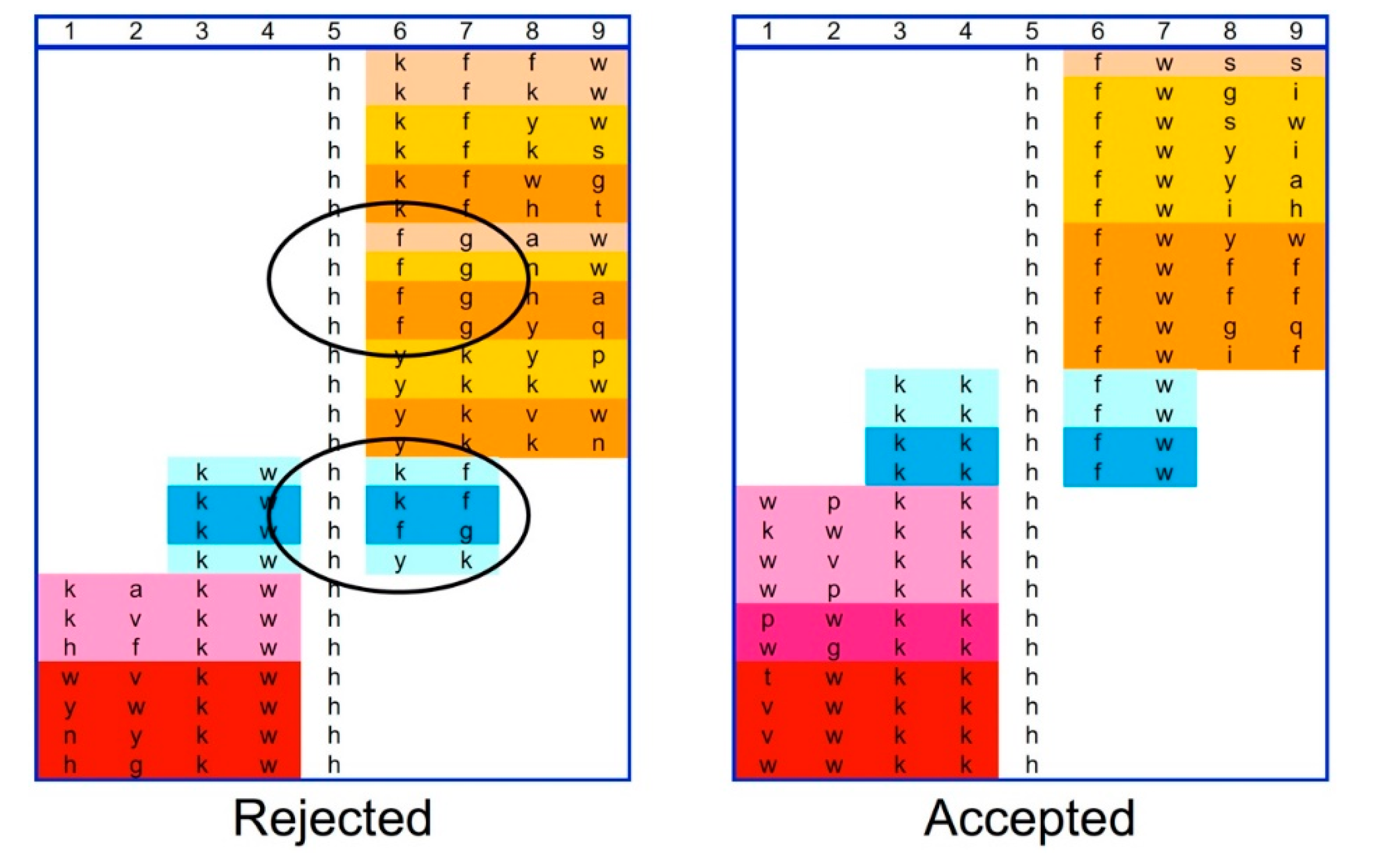
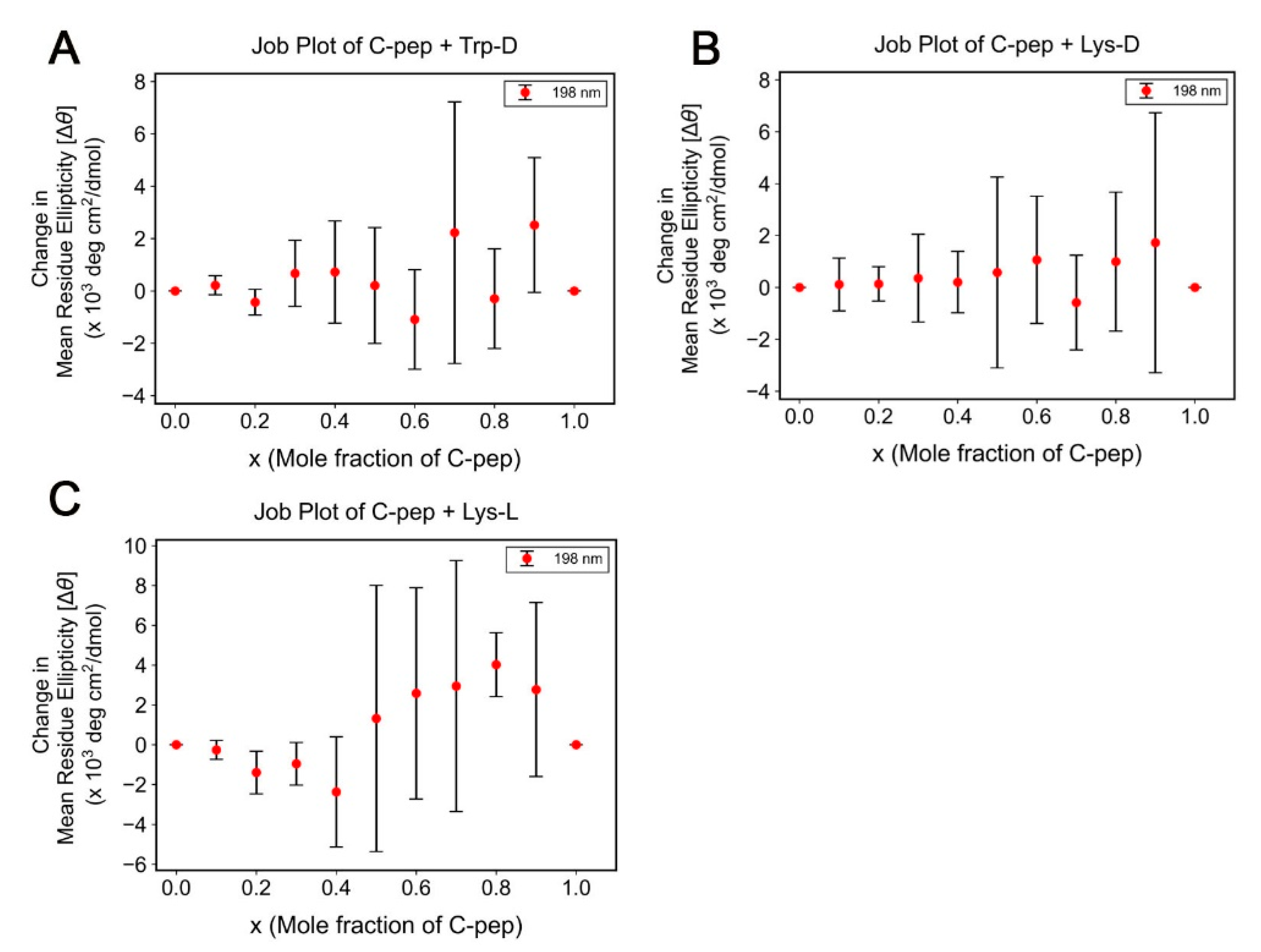
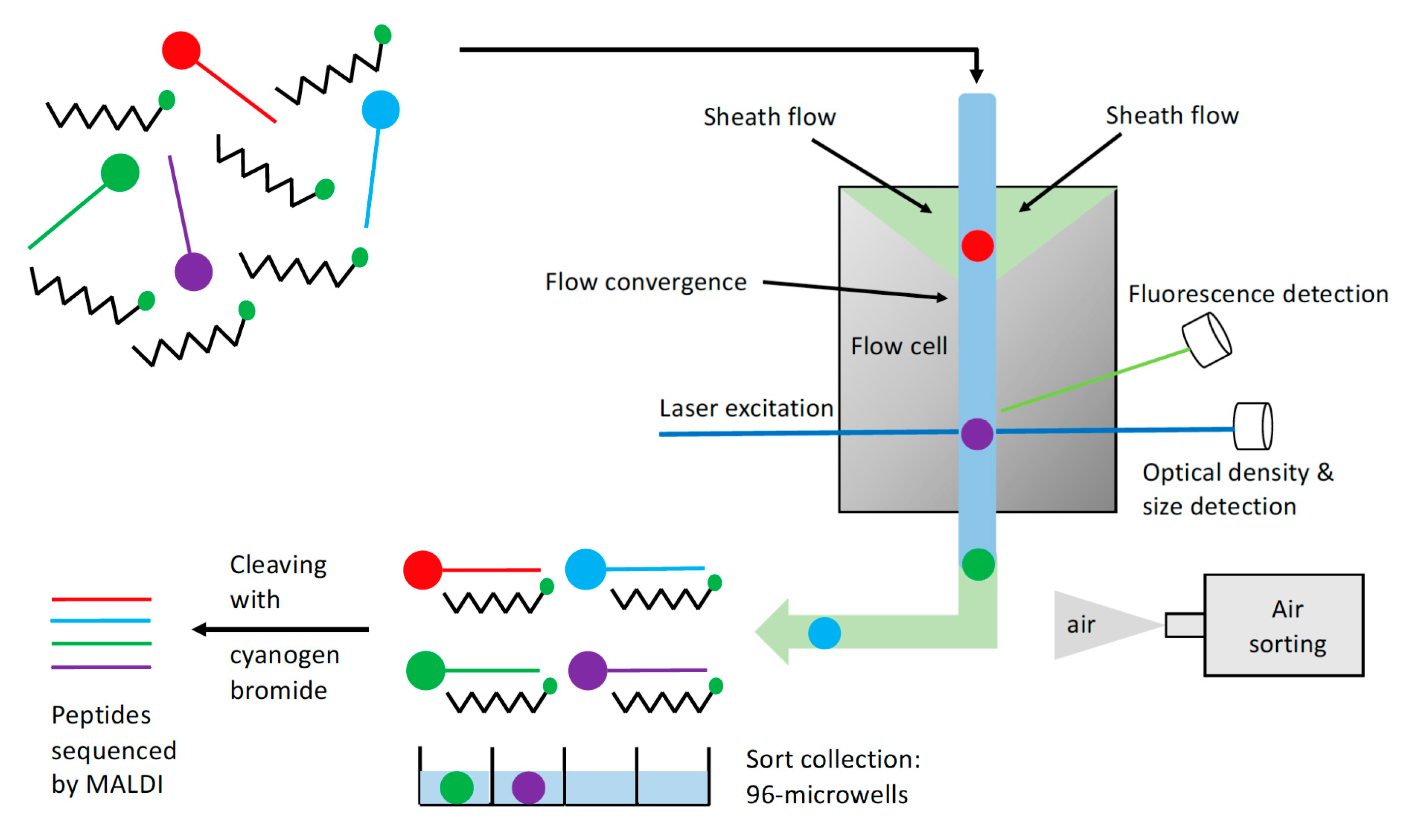
2.1. Bead Screening Strategy for Complementary Peptide to C-Peptide
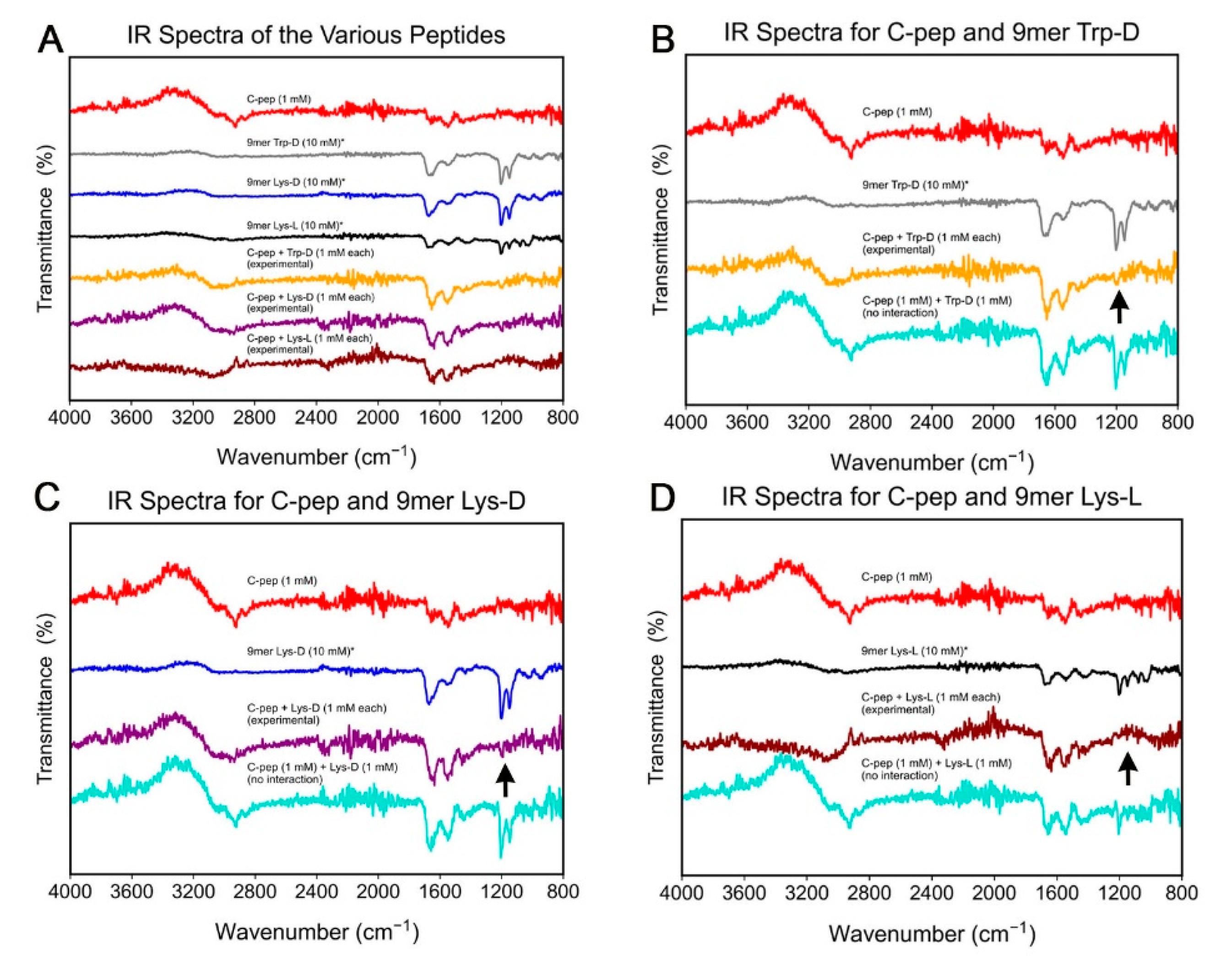
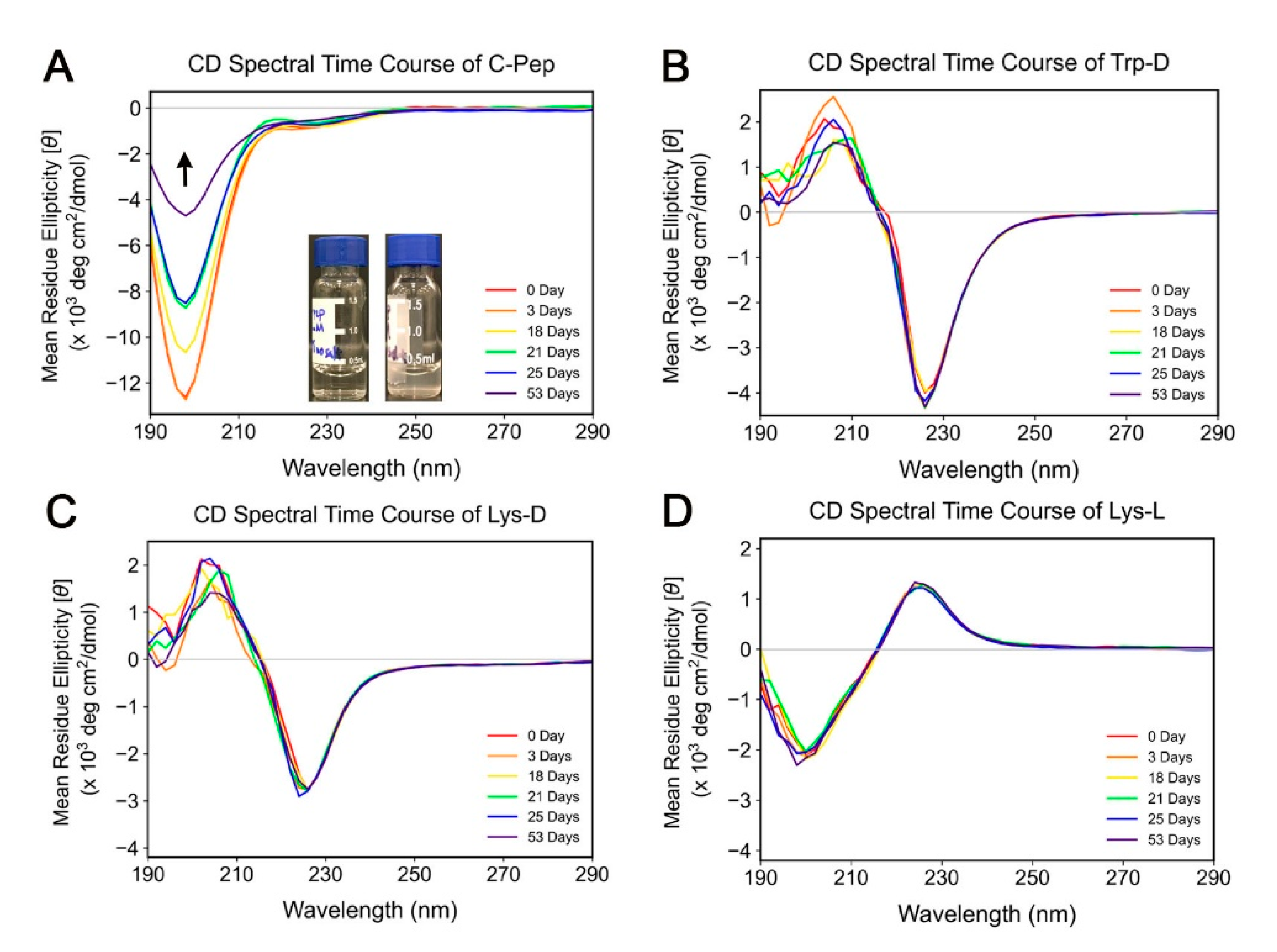
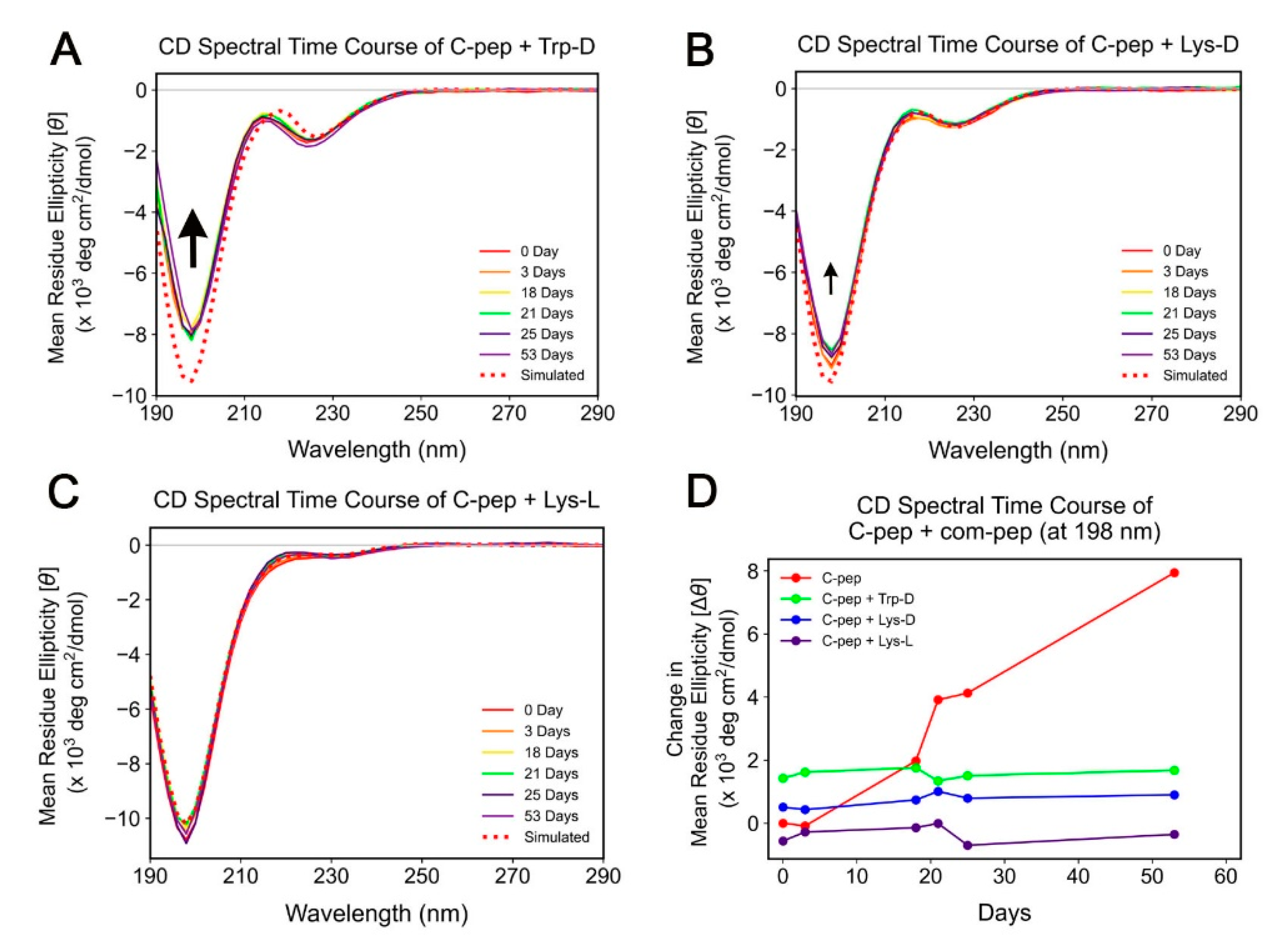
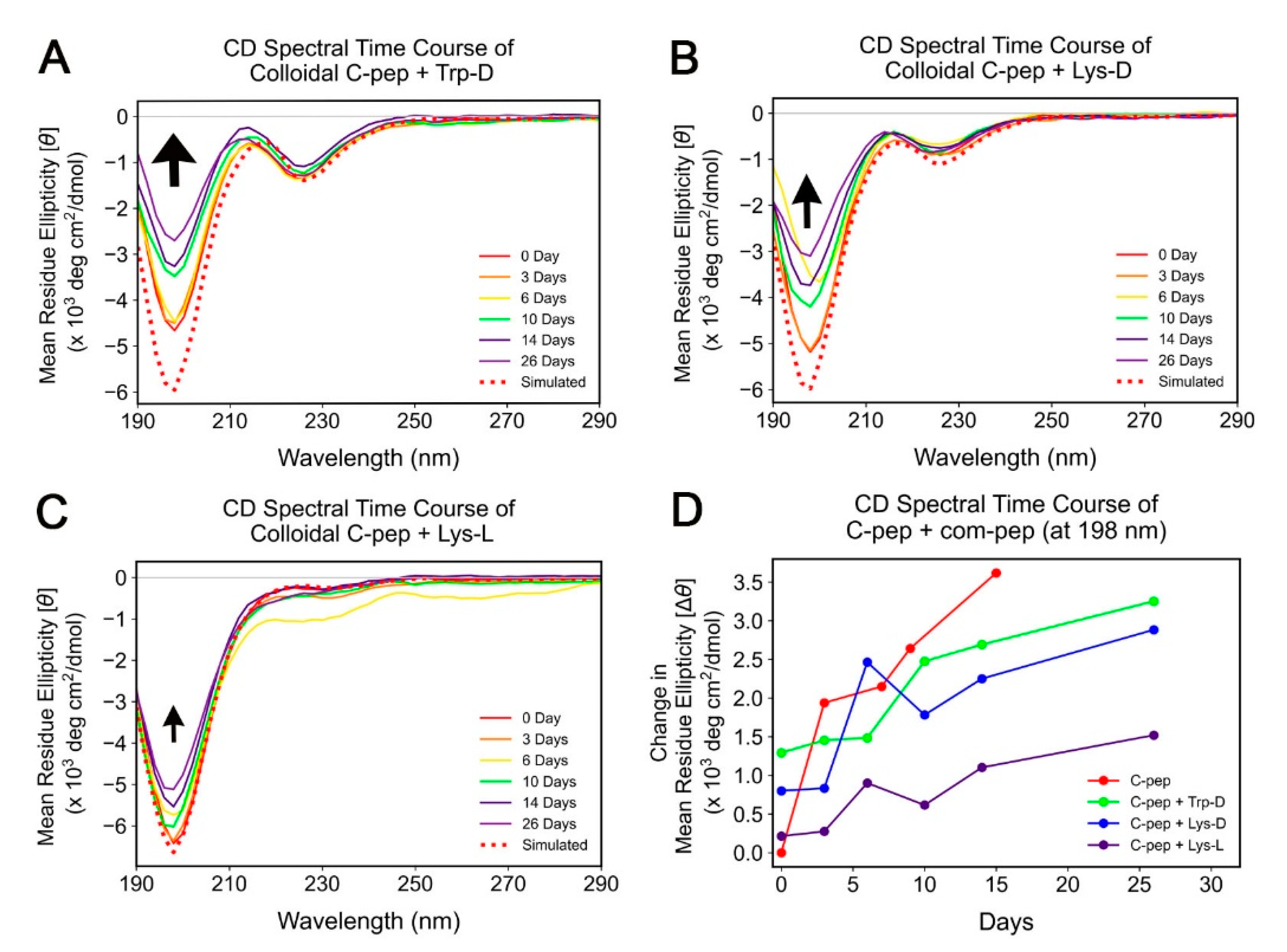
2.2. Spectroscopic Study of C-Peptide–Com-Peptide Coassembly
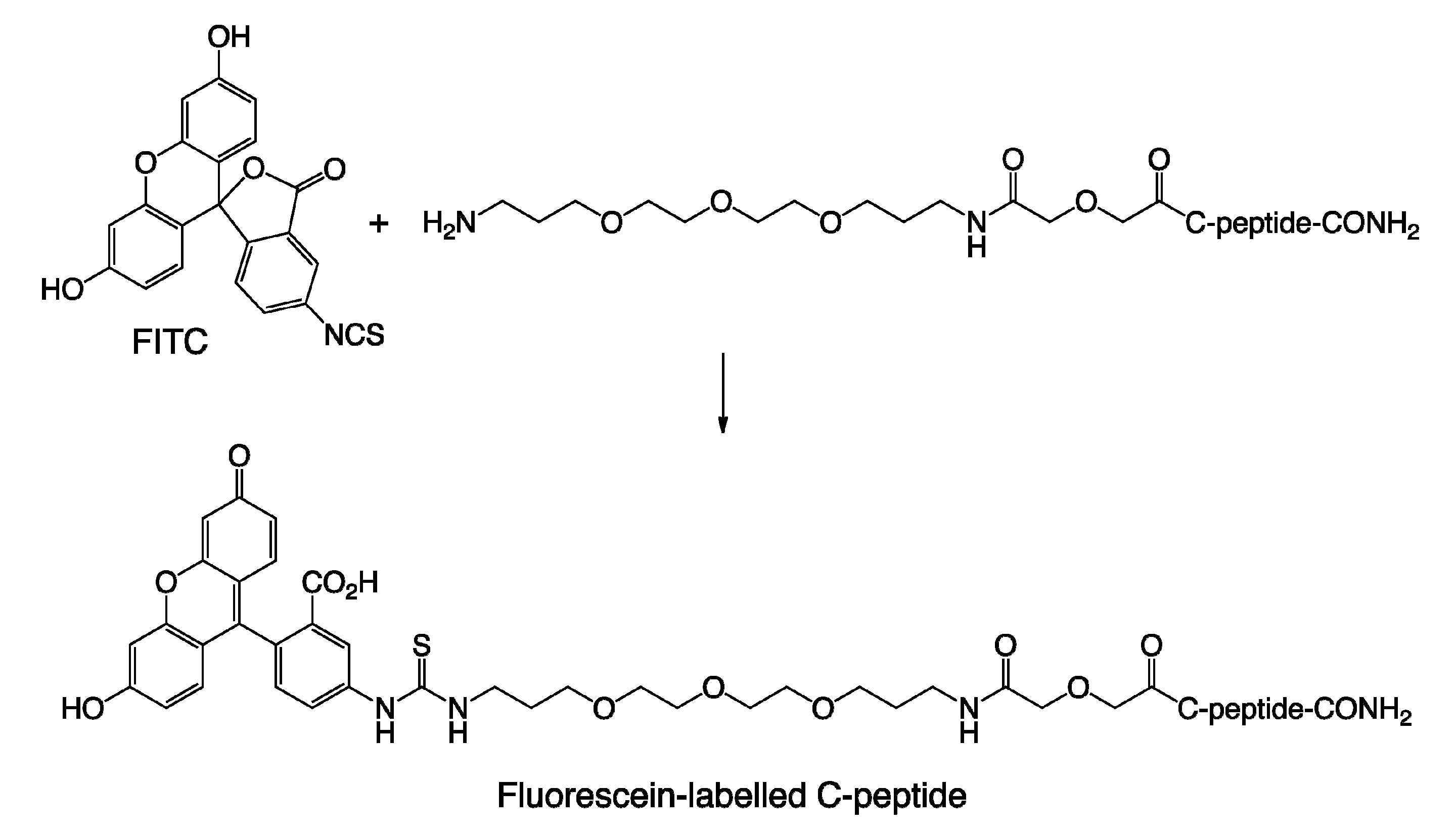
3. Materials and Method
4. Conclusion and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wahren, J.; Kallas, A.; Sima, A.A.F. The Clinical Potential of C-Peptide Replacement in Type 1 Diabetes. Diabetes 2012, 61, 761–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, M.A.; Ekberg, K.; Wahren, J.; Cameron, N.E. Effects of proinsulin C-peptide in experimental diabetic neuropathy: Vascular actions and modulation by nitric oxide synthase inhibition. Diabetes 2003, 52, 1812–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekberg, K.; Brismar, T.; Johansson, B.L.; Jonsson, B.; Lindstrom, P.; Wahren, J. Amelioration of sensory nerve dysfunction by C-peptide in patients with type 1 diabetes. Diabetes 2003, 52, 536–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagida, K.; Okochi, M.; Tagami, S.; Nakayama, T.; Kodama, T.S.; Nishitomi, K.; Jiang, J.; Mori, K.; Tatsumi, S.-i.; Arai, T.; et al. The 28-amino acid form of an APLP1-derived A beta-like peptide is a surrogate marker for A beta 42 production in the central nervous system. EMBO Mol. Med. 2009, 1, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Kandil, E.; Burack, J.; Sawas, A.; Bibawy, H.; Schwartzman, A.; Zenilman, M.E.; Bluth, M.H. B-type natriuretic peptide. Arch. Surg. 2008, 143, 242–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIlroy, D.R.; Wallace, S.; Roubos, N. Brain Natriuretic Peptide (BNP) as a Biomarker of Myocardial Ischemia-Reperfusion Injury in Cardiac Transplantation. J. Cardiothorac. Vasc. Anesth. 2010, 24, 939–945. [Google Scholar] [CrossRef]
- Jones, A.G.; Hattersley, A.T. The clinical utility of C-peptide measurement in the care of patients with diabetes. Diabet. Med. 2013, 30, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Leighton, E.; Sainsbury, C.A.R.; Jones, G.C. A Practical Review of C-Peptide Testing in Diabetes. Diabetes Ther. 2017, 8, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.-J.; Tsai, F.-J.; Ho, C.-M.; Liu, Y.-C.; Chen, C.-J. Peptide Biomarker Discovery for Identification of Methicillin-Resistant and Vancomycin-Intermediate Staphylococcus aureus Strains by MALDI-TOF. Anal. Chem. 2012, 84, 5685–5692. [Google Scholar] [CrossRef]
- Carrera, M.; Canas, B.; Lopez-Ferrer, D.; Pineiro, C.; Vazquez, J.; Gallardo, J.M. Fast Monitoring of Species-Specific Peptide Biomarkers Using High-Intensity-Focused-Ultrasound-Assisted Tryptic Digestion and Selected MS/MS Ion Monitoring. Anal. Chem. 2011, 83, 5688–5695. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, J.; Lawlor, K.; Toledo-Crow, R.; Tempst, P. Automated serum peptide profiling. Nat. Protoc. 2006, 1, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Berti, F. Short peptides as biosensor transducers. Anal. Bioanal. Chem. 2012, 402, 3055–3070. [Google Scholar] [CrossRef] [PubMed]
- Makam, P.; Gazit, E. Minimalistic peptide supramolecular co-assembly: Expanding the conformational space for nanotechnology. Chem. Soc. Rev. 2018, 47, 3406–3420. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Wong, K.M.; Seroski, D.T.; Wang, Y.M.; Liu, R.J.; Paravastu, A.K.; Hudalla, G.A.; Hall, C.K. Anatomy of a selectively coassembled beta-sheet peptide nanofiber. Proc. Natl. Acad. Sci. USA 2020, 117, 4710–4717. [Google Scholar] [CrossRef]
- Chan, K.H.; Tay, J.J.J. Advancement of Peptide Nanobiotechnology via Emerging Microfluidic Technology. Micromachines 2019, 10, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bera, S.; Mondal, S.; Tang, Y.M.; Jacoby, G.; Arad, E.; Guterman, T.; Jelinek, R.; Beck, R.; Wei, G.H.; Gazit, E. Deciphering the Rules for Amino Acid Co-Assembly Based on Interlayer Distances. Acs. Nano. 2019, 13, 1703–1712. [Google Scholar] [CrossRef] [PubMed]
- Agnew, H.D.; Coppock, M.B.; Idso, M.N.; Lai, B.T.; Liang, J.X.; McCarthy-Torrens, A.M.; Warren, C.M.; Heath, J.R. Protein-Catalyzed Capture Agents. Chem. Rev. 2019, 119, 9950–9970. [Google Scholar] [CrossRef] [PubMed]
- Jee, J.E.; Lim, J.; Ong, Y.S.; Oon, J.; Gao, L.Q.; Choi, H.S.; Lee, S.S. An efficient strategy to enhance binding affinity and specificity of a known isozyme inhibitor. Org. Biomol. Chem. 2016, 14, 6833–6839. [Google Scholar] [CrossRef] [Green Version]
- Agnew, H.D.; Rohde, R.D.; Millward, S.W.; Nag, A.; Yeo, W.-S.; Hein, J.E.; Pitram, S.M.; Tariq, A.A.; Burns, V.M.; Krom, R.J.; et al. Iterative In Situ Click Chemistry Creates Antibody-like Protein-Capture Agents. Angew. Chem. Int. Ed. 2009, 48, 4944–4948. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Shin, D.-S.; Kim, E.-M.; Park, H.-Y.; Lee, C.-S.; Kim, J.-H.; Lee, B.-S.; Lee, Y.-S.; Kim, B.-G. High-throughput identification of substrate specificity for protein kinase by using an improved one-bead-one-compound library approach. Angew. Chem. Int. Ed. 2007, 46, 5408–5411. [Google Scholar] [CrossRef]
- Gonzalez-Vera, J.A.; Lukovic, E.; Imperiali, B. A rapid method for generation of selective Sox-based chemosensors of Ser/Thr kinases using combinatorial peptide libraries. Bioorg. Med. Chem. Lett. 2009, 19, 1258–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millward, S.W.; Henning, R.K.; Kwong, G.A.; Pitram, S.; Agnew, H.D.; Deyle, K.M.; Nag, A.; Hein, J.; Lee, S.S.; Lim, J.; et al. Iterative in Situ Click Chemistry Assembles a Branched Capture Agent and Allosteric Inhibitor for Akt1. J. Am. Chem. Soc. 2011, 133, 18280–18288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Shirotsuki, S.; Iwata, T.; Kageyama, C.; Tahara, T.; Nozaki, S.; Siwu, E.R.O.; Tamura, S.; Douke, S.; Murakam, N.; et al. Template-Assisted and Self-Activating Clicked Peptide as a Synthetic Mimic of the SH2 Domain. ACS Chem. Biol. 2012, 7, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Chen, M.S.; Chen, Y.X.; Zhao, Y.F.; Li, Y.M. Short Peptide Segment and Insulin Co-Assembly Forms Cytotoxic Oligomers. Int. J. Pept. Res. Ther. 2013, 19, 185–189. [Google Scholar] [CrossRef]
- Lee, S.S.; Lim, J.; Cha, J.; Tan, S.; Heath, J.R. Rapid Microwave-Assisted CNBr Cleavage of Bead-Bound Peptides. J. Comb. Chem. 2008, 10, 807–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.S.; Lim, J.; Tan, S.; Cha, J.; Yeo, S.Y.; Agnew, H.D.; Heath, J.R. Accurate MALDI-TOF/TOF Sequencing of One-Bead-One-Compound Peptide Libraries with Application to the Identification of Multiligand Protein Affinity Agents Using in Situ Click Chemistry Screening. Anal. Chem. 2010, 82, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Ko, A.S.C.; Smyth, D.G.; Markusse, J.; Sundby, F. Amino Acid Sequence of C-Peptide of Human Proinsulin. Eur. J. Biochem. 1971, 20, 190. [Google Scholar] [CrossRef]
- Chan, K.H.; Xue, B.; Robinson, R.C.; Hauser, C.A.E. Systematic Moiety Variations of Ultrashort Peptides Produce Profound Effects on Self-Assembly, Nanostructure Formation, Hydrogelation, and Phase Transition. Sci. Rep. 2017, 7, 12897. [Google Scholar] [CrossRef] [Green Version]
- Vendruscolo, M.; Paci, E.; Dobson, C.M.; Karplus, M. Three key residues form a critical contact network in a protein folding transition state. Nature 2001, 409, 641–645. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Frederix, P.; Ide, J.; Altay, Y.; Schaeffer, G.; Surin, M.; Beljonne, D.; Bondarenko, A.S.; Jansen, T.L.C.; Otto, S.; Marrink, S.J. Structural and Spectroscopic Properties of Assemblies of Self-Replicating Peptide Macrocycles. Acs. Nano. 2017, 11, 7858–7868. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Jiang, Y.X.; Xiong, W.; Li, H.; Zhang, P.Y.; Yin, F.; Zhang, Q.L.; Geng, H.; Jiang, F.; Li, Z.; et al. Tuning peptide self-assembly by an in-tether chiral center. Sci. Adv. 2018, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurouski, D.; Van Duyne, R.P.; Lednev, I.K. Exploring the structure and formation mechanism of amyloid fibrils by Raman spectroscopy: A review. Analyst 2015, 140, 4967–4980. [Google Scholar] [CrossRef] [PubMed]
- Balan, V.; Mihai, C.T.; Cojocaru, F.D.; Uritu, C.M.; Dodi, G.; Botezat, D.; Gardikiotis, I. Vibrational Spectroscopy Fingerprinting in Medicine: From Molecular to Clinical Practice. Materials 2019, 12, 2884. [Google Scholar] [CrossRef] [Green Version]
- Santiveri, C.M.; Angeles Jimenez, M. Tryptophan Residues: Scarce in Proteins but Strong Stabilizers of beta-Hairpin Peptides. Biopolymers 2010, 94, 779–790. [Google Scholar] [CrossRef]
- Nasstrom, T.; Aden, J.; Shibata, F.; Andersson, P.O.; Karlsson, B.C.G. A Capped Peptide of the Aggregation Prone NAC 71–82 Amino Acid Stretch of alpha-Synuclein Folds into Soluble beta-Sheet Oligomers at Low and Elevated Peptide Concentrations. Int. J. Mol. Sci. 2020, 21, 1629. [Google Scholar] [CrossRef] [Green Version]
- Rucker, A.L.; Creamer, T.P. Polyproline II helical structure in protein unfolded states: Lysine peptides revisited. Protein Sci. 2002, 11, 980–985. [Google Scholar]
- Shi, Z.S.; Olson, C.A.; Rose, G.D.; Baldwin, R.L.; Kallenbach, N.R. Polyproline II structure in a sequence of seven alanine residues. Proc. Natl. Acad. Sci. USA 2002, 99, 9190–9195. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, N.J. Using circular dichroism collected as a function of temperature to determine the thermodynamics of protein unfolding and binding interactions. Nat. Protoc. 2006, 1, 2527–2535. [Google Scholar] [CrossRef]
- Wang, M.; Zhou, P.; Wang, J.Q.; Zhao, Y.R.; Ma, H.C.; Lu, J.R.; Xu, H. Left or Right: How Does Amino Acid Chirality Affect the Handedness of Nanostructures Self-Assembled from Short Amphiphilic Peptides? J. Am. Chem. Soc. 2017, 139, 4185–4194. [Google Scholar] [CrossRef] [Green Version]
- Lind, J.; Lindahl, E.; Peralvarez-Marin, A.; Holmlund, A.; Jornvall, H.; Maler, L. Structural features of proinsulin C-peptide oligomeric and amyloid states. FEBS J. 2010, 277, 3759–3768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, S.Y.; Yang, H.; Chen, P. Formation of colloidal suspension of hydrophobic compounds with an amphiphilic self-assembling peptide. Colloids Surf. B-Biointerfaces 2007, 55, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Truex, N.L.; Nowick, J.S. Coassembly of Peptides Derived from beta-Sheet Regions of beta-Amyloid. J. Am. Chem. Soc. 2016, 138, 13891–13900. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.L.; Harris, J.L.; Khanna, K.K.; Hong, J.H. A Comprehensive Review on Current Advances in Peptide Drug Development and Design. Int. J. Mol. Sci. 2019, 20, 2383. [Google Scholar] [CrossRef] [Green Version]
- Jerabek-Willemsen, M.; Andre, T.; Wanner, R.; Roth, H.M.; Duhr, S.; Baaske, P.; Breitsprecher, D. MicroScale Thermophoresis: Interaction analysis and beyond. J. Mol. Struct. 2014, 1077, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Schauenstein, K.; Schauenstein, E.; Wick, G. Fluorescence Properties of Free and Protein Bound Fluorescein Dyes. J. Histochem. Cytochem. 1978, 26, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.; Lim, J.; Zheng, Y.; Tan, S.; Ang, Y.L.; Oon, J.; Ang, M.W.; Ling, J.; Bode, M.; Lee, S.S. Process Automation toward Ultra-High-Throughput Screening of Combinatorial One-Bead-One-Compound (OBOC) Peptide Libraries. J. Lab. Automat. 2012, 17, 186–200. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Chan, K.H.; Reithofer, M.R.; Hauser, C.A.E. Influence of metal salts on the hydrogelation properties of ultrashort aliphatic peptides. Rsc. Adv. 2013, 3, 9985–9993. [Google Scholar] [CrossRef]
- Chan, K.H.; Lee, W.H.; Ni, M.; Loo, Y.; Hauser, C.A.E. C-Terminal Residue of Ultrashort Peptides Impacts on Molecular Self-Assembly, Hydrogelation, and Interaction with Small-Molecule Drugs. Sci. Rep. 2018, 8, 17127. [Google Scholar] [CrossRef]
- Smadbeck, J.; Chan, K.H.; Khoury, G.A.; Xue, B.; Robinson, R.C.; Hauser, C.A.E.; Floudas, C.A. De Novo Design and Experimental Characterization of Ultrashort Self-Associating Peptides. Plos Comput. Biol. 2014, 10, e1003718. [Google Scholar] [CrossRef] [Green Version]
- Tuttle, T. Computational Approaches to Understanding the Self-assembly of Peptide-based Nanostructures. Isr. J. Chem. 2015, 55, 724–734. [Google Scholar] [CrossRef] [Green Version]
- Caputo, C.B.; Salama, A.I. The Amyloid Proteins Of Alzheimers-Disease As Potential Targets For Drug-Therapy. Neurobiol. Aging 1989, 10, 451–461. [Google Scholar] [CrossRef]
- Ladiwala, A.R.A.; Bhattacharya, M.; Perchiacca, J.M.; Cao, P.; Raleigh, D.P.; Abedini, A.; Schmidt, A.M.; Varkey, J.; Langen, R.; Tessier, P.M. Rational design of potent domain antibody inhibitors of amyloid fibril assembly. Proc. Natl. Acad. Sci. USA 2012, 109, 19965–19970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iscen, A.; Brue, C.R.; Roberts, K.F.; Kim, J.; Schatz, G.C.; Meade, T.J. Inhibition of Amyloid-beta Aggregation by Cobalt(III) Schiff Base Complexes: A Computational and Experimental Approach. J. Am. Chem. Soc. 2019, 141, 16685–16695. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.X.; Cao, Q.; Wang, C.C.; Zheng, J.; Luo, F.; Xie, J.F.; Li, Y.C.; Ma, X.J.; He, L.; Eisenberg, D.; et al. Structure-Based Peptide Inhibitor Design of Amyloid-beta Aggregation. Front. Mol. Neurosci. 2019, 12, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidler, P.M.; Boyer, D.R.; Rodriguez, J.A.; Sawaya, M.R.; Cascio, D.; Murray, K.; Gonen, T.; Eisenberg, D.S. Structure-based inhibitors of tau aggregation. Nat. Chem. 2018, 10, 170–176. [Google Scholar] [CrossRef]
- Hard, T.; Lendel, C. Inhibition of Amyloid Formation. J. Mol. Biol. 2012, 421, 441–465. [Google Scholar] [CrossRef]
- Sharma, V.; Ghosh, K.S. Inhibition of Amyloid Fibrillation by Small Molecules and Nanomaterials: Strategic Development of Pharmaceuticals Against Amyloidosis. Protein Pept. Lett. 2019, 26, 315–323. [Google Scholar] [CrossRef]
- Acx, H.; Chavez-Gutierrez, L.; Serneels, L.; Lismont, S.; Benurwar, M.; Elad, N.; De Strooper, B. Signature Amyloid beta Profiles Are Produced by Different gamma-Secretase Complexes. J. Biol. Chem. 2014, 289, 4346–4355. [Google Scholar] [CrossRef] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, K.H.; Lim, J.; Jee, J.E.; Aw, J.H.; Lee, S.S. Peptide–Peptide Co-Assembly: A Design Strategy for Functional Detection of C-peptide, A Biomarker of Diabetic Neuropathy. Int. J. Mol. Sci. 2020, 21, 9671. https://doi.org/10.3390/ijms21249671
Chan KH, Lim J, Jee JE, Aw JH, Lee SS. Peptide–Peptide Co-Assembly: A Design Strategy for Functional Detection of C-peptide, A Biomarker of Diabetic Neuropathy. International Journal of Molecular Sciences. 2020; 21(24):9671. https://doi.org/10.3390/ijms21249671
Chicago/Turabian StyleChan, Kiat Hwa, Jaehong Lim, Joo Eun Jee, Jia Hui Aw, and Su Seong Lee. 2020. "Peptide–Peptide Co-Assembly: A Design Strategy for Functional Detection of C-peptide, A Biomarker of Diabetic Neuropathy" International Journal of Molecular Sciences 21, no. 24: 9671. https://doi.org/10.3390/ijms21249671
APA StyleChan, K. H., Lim, J., Jee, J. E., Aw, J. H., & Lee, S. S. (2020). Peptide–Peptide Co-Assembly: A Design Strategy for Functional Detection of C-peptide, A Biomarker of Diabetic Neuropathy. International Journal of Molecular Sciences, 21(24), 9671. https://doi.org/10.3390/ijms21249671