Yeast Microbiota during Sauerkraut Fermentation and Its Characteristics


Abstract
:1. Introduction
2. Results
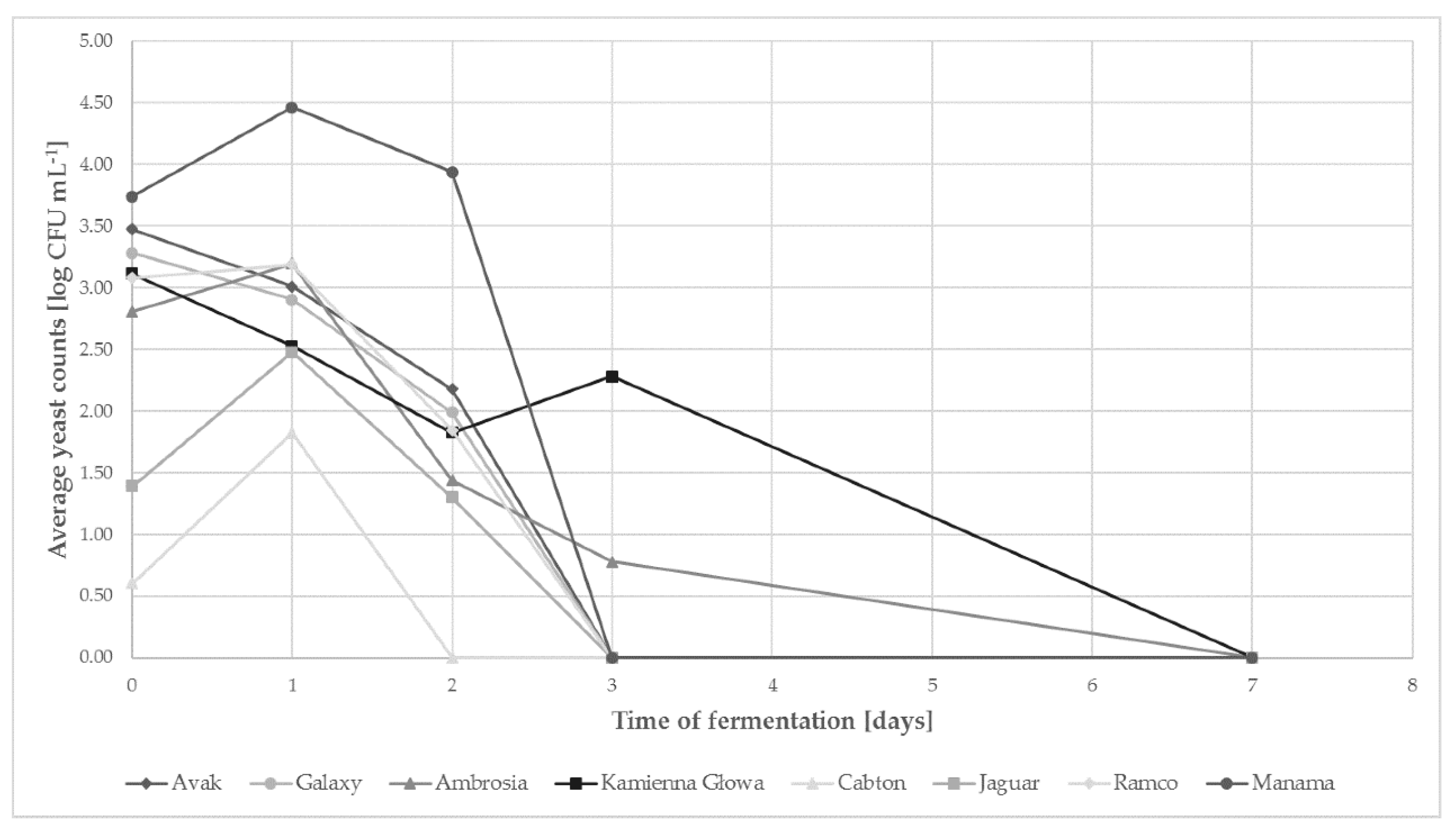
2.1. Yeast Population Kinetics
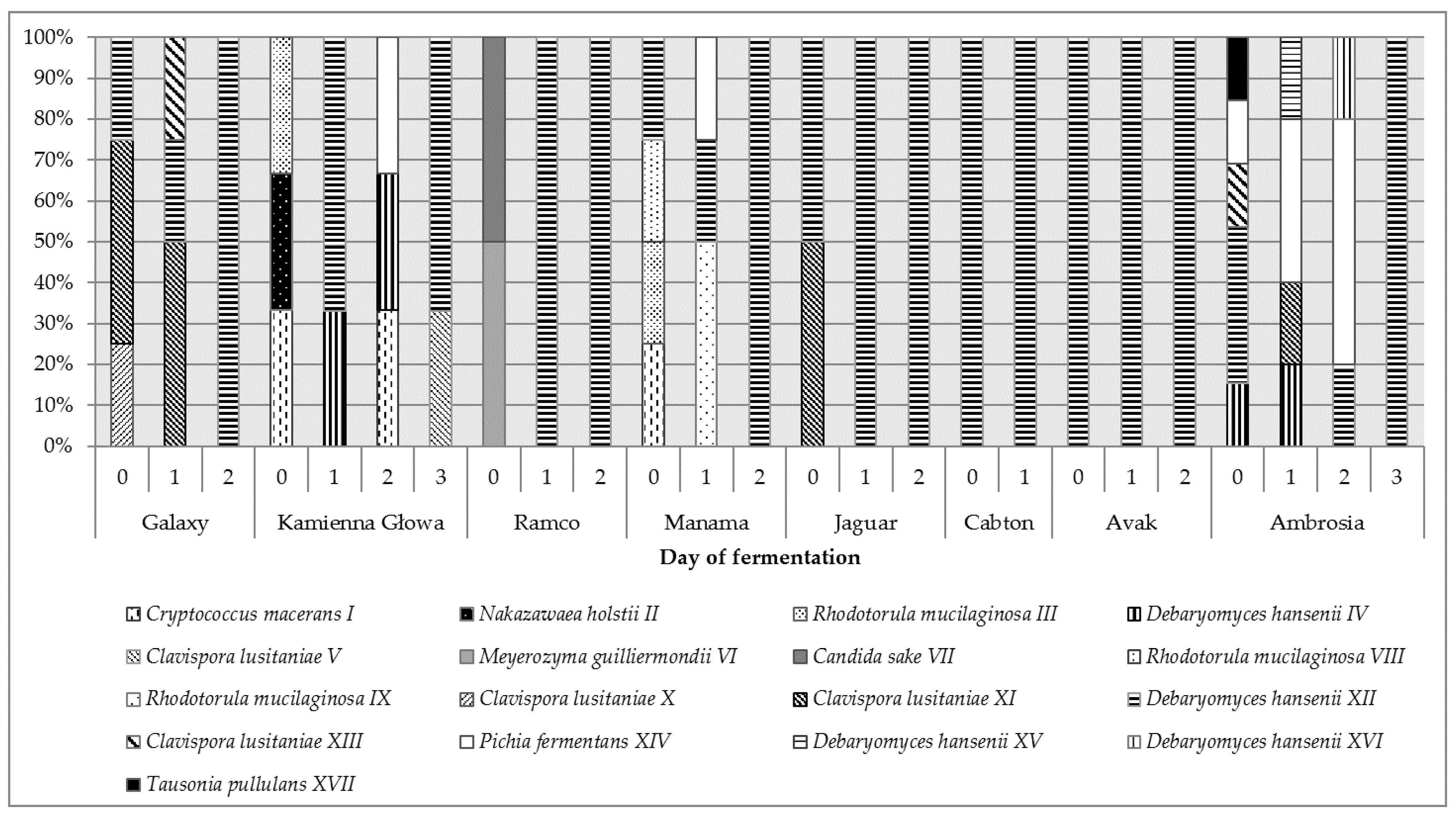
2.2. Biodiversity of Yeasts during Fermentation
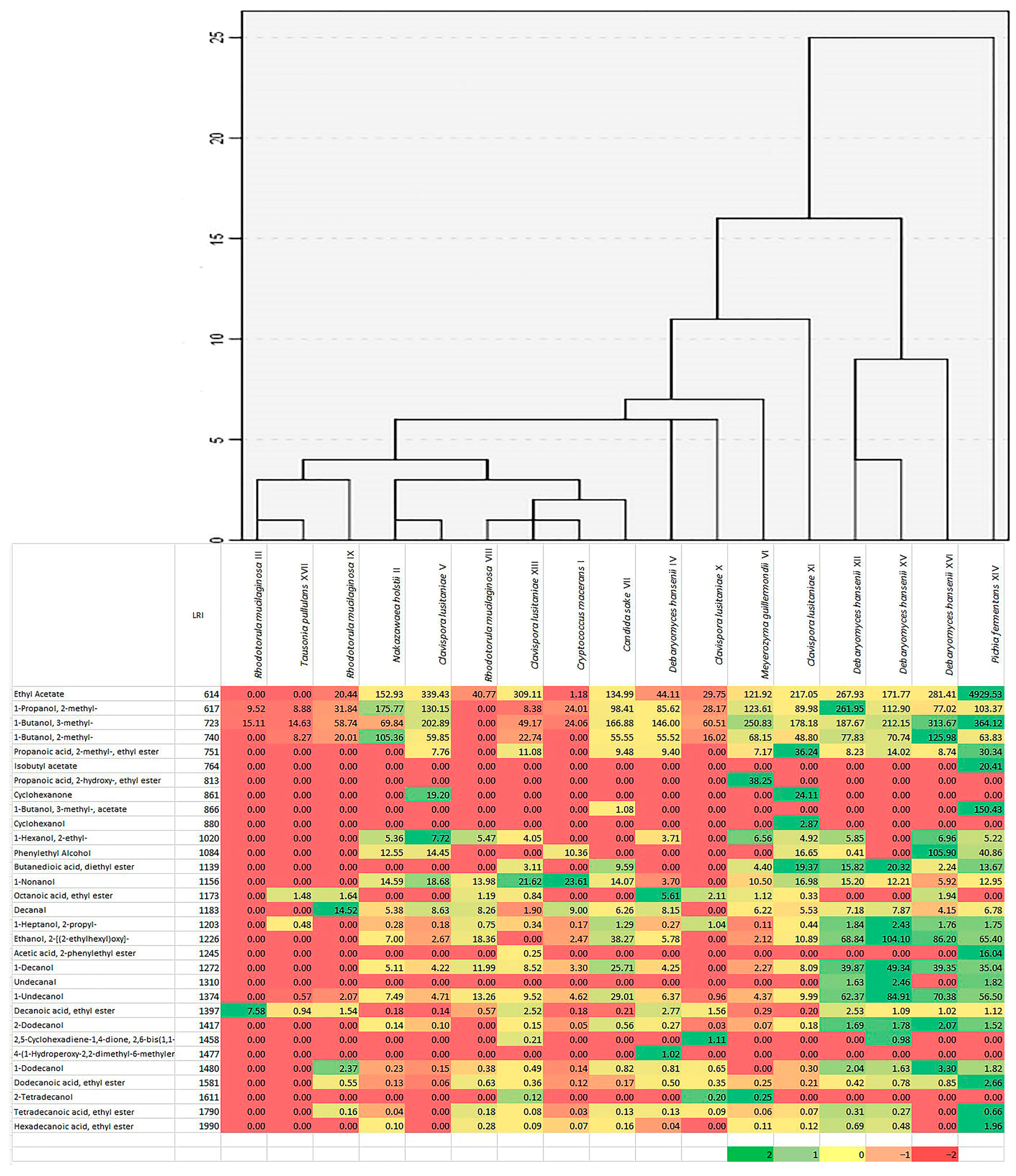
2.3. Production of Volatile Components by Isolates
2.4. Resistance of Isolates to Selected Stress Factors Present during Sauerkraut Fermentation and Killer Activity
3. Discussion
4. Materials and Methods
4.1. Cabbages and Fermentation of Sauerkraut
4.2. Yeast Enumeration and Isolation
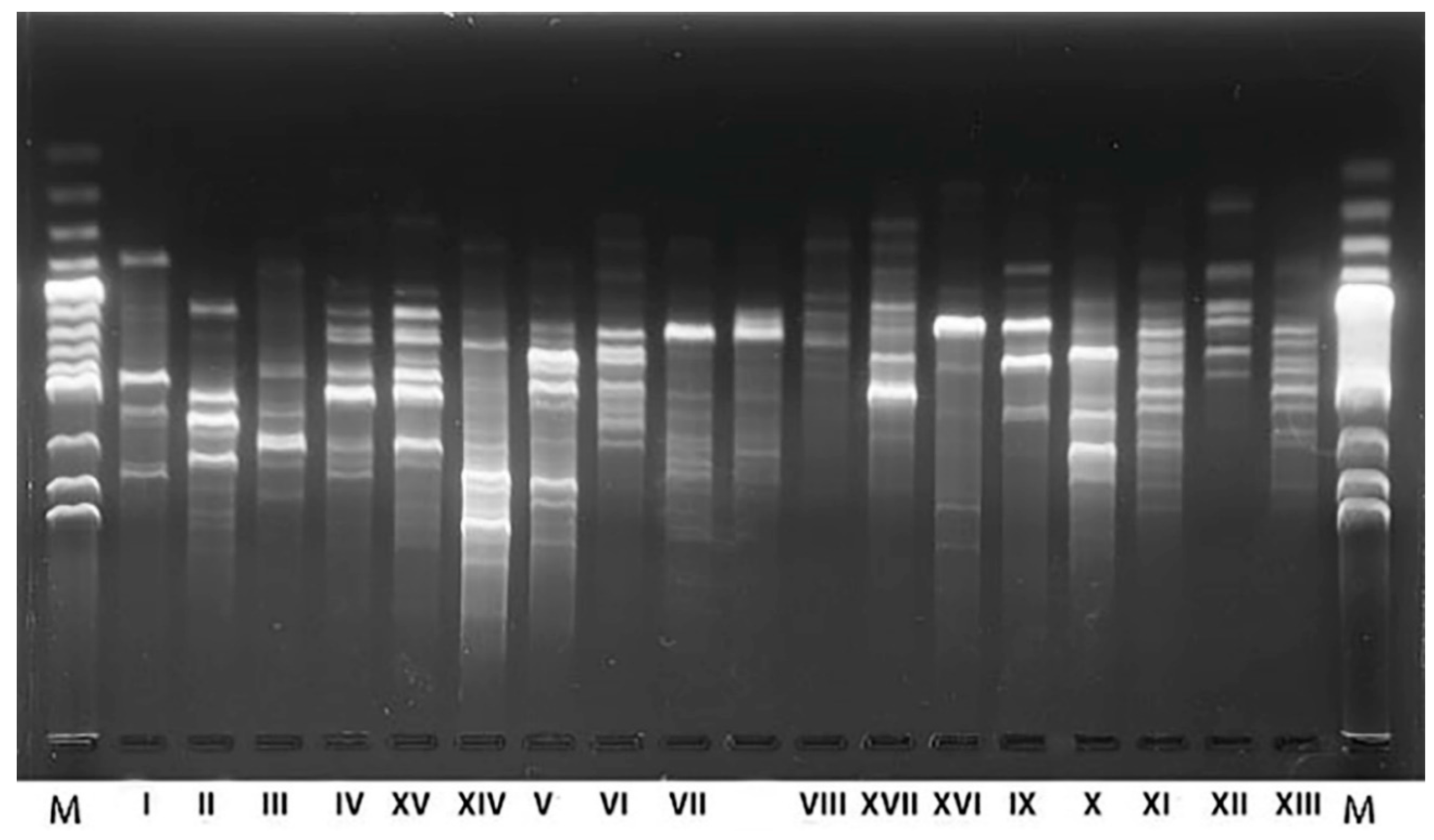
4.3. DNA Extraction and RAPD-PCR Analysis
4.4. PCR-RFLP Analysis and 5.8 S-ITS rRNA Gene Region Sequencing
4.5. Production of Volatile Components by Isolates (SPME-GC-TOFMS)
4.6. Resistance of Isolates to Selected Stress Factors Present during Sauerkraut Fermentation
4.7. Killer Activity
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Filipiak, T.; Maciejczak, M. Vegetable production in Poland and selected countries of the European Union. Econ. Sci. Rural Dev. 2011, 2, 30–39. [Google Scholar]
- Beganović, J.; Kos, B.; Pavunc, A.L.; Uroić, K.; Jokić, M.; Šušković, J. Traditionally produced sauerkraut as source of autochthonous functional starter cultures. Microbiol. Res. 2014, 169, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Kusznierewicz, B.; Śmiechowska, A.; Bartoszek, A.; Namieśnik, J. The effect of heating and fermenting on antioxidant properties of white cabbage. Food Chem. 2008, 108, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Peñas, E.; Frias, J.; Gomez, R.; Vidal-Valverde, C. High hydrostatic pressure can improve the microbial quality of sauerkraut during storage. Food Control 2010, 21, 524–528. [Google Scholar] [CrossRef]
- Plengvidhya, V.; Breidt, F., Jr.; Lu, Z.; Fleming, H.P. DNA fingerprinting of lactic acid bacteria in sauerkraut fermentations. Appl. Environ. Microbiol. 2007, 73, 7697–7702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.-Y. Fermentation. In Handbook of Food and Beverage Fermentation Technology; Hui, Y.H., Meunier-Goddik, L., Hansen, Å.S., Josephsen, J., Nip, W.K., Stanfield, P.S., Toldrá, F., Eds.; Marcel Dekker Inc.: New York, USA, 2004; pp. 595–610. [Google Scholar]
- Hang, Y.D. Sauerkraut. In Handbook of Food and Beverage Fermentation Technology; Hui, Y.H., Meunier-Goddik, L., Hansen, Å.S., Josephsen, J., Nip, W.K., Stanfield, P.S., Toldrá, F., Eds.; Marcel Dekker Inc.: New York, USA, 2004; pp. 669–676. [Google Scholar]
- Shih, C.T.; Hang, Y.D. Production of carotenoids by Rhodotorula rubra from sauerkraut brine. LTW-Food Sci Technol 1996, 29, 570–572. [Google Scholar]
- Du, R.; Ge, J.; Zhao, D.; Sun, J.; Ping, W.; Song, G. Bacterial diversity and community structure during fermentation of Chinese sauerkraut with Lactobacillus casei 11MZ-5-1 by Illumina Miseq sequencing. Lett. Appl. Microbiol. 2018, 66, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, A.; Rösch, N.; Cho, G.S.; Meinhardt, A.K.; Kabisch, J.; Habermann, D.; Böhnlein, C.; Brinks, E.; Greiner, R.; Franz, C.M.A.P. Influence of iodized table salt on fermentation characteristics and bacterial diversity during sauerkraut fermentation. Food Microbiol. 2018, 76, 473–480. [Google Scholar] [CrossRef]
- Touret, T.; Oliveira, M.; Semedo-Lemsaddek, T. Putative probiotic lactic acid bacteria isolated from sauerkraut fermentations. PLoS ONE 2018, 13, e0203501. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Zang, S.; Zhao, Z.; Li, X. Dynamic changes of bacterial communities and nitrite character during northeastern Chinese sauerkraut fermentation. Food Sci. Biotechnol. 2017, 27, 79–85. [Google Scholar] [CrossRef]
- Beljaars, P.R.; Van Dijk, R.; Jonker, K.M.; Schout, L.J. Liquid chromatographic determination of histamine in fish, sauerkraut, and wine: Interlaboratory study. J. AOAC Int. 1998, 81, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallmann, E.; Kazimierczak, R.; Marszałek, K.; Drela, N.; Kiernozek, E.; Toomik, P.; Matt, D.; Luik, A.; Rembiałkowska, E. The Nutritive Value of Organic and Conventional White Cabbage (Brassica Oleracea L. Var. Capitata) and Anti-Apoptotic Activity in Gastric Adenocarcinoma Cells of Sauerkraut Juice Produced Therof. J. Agric. Food Chem. 2017, 65, 8171–8183. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Villaluenga, C.; Peñas, E.; Frias, J.; Ciska, E.; Honke, J.; Piskula, M.K.; Kozlowska, H.; Vidal-Valverde, C. Influence of fermentation conditions on glucosinolates, ascorbigen, and ascorbic acid content in white cabbage (Brassica oleracea var. capitata cv. Taler) cultivated in different seasons. J. Food Sci. 2009, 74, C62–C67. [Google Scholar] [PubMed]
- Palani, K.; Harbaum-Piayda, B.; Meske, D.; Keppler, J.K.; Bockelmann, W.; Heller, K.J.; Schwarz, K. Influence of fermentation on glucosinolates and glucobrassicin degradation products in sauerkraut. Food Chem. 2016, 190, 755–762. [Google Scholar] [CrossRef]
- Harbaum-Piayda, B.; Palani, K.; Schwarz, K. Influence of postharvest UV-B treatment and fermentation on secondary plant compounds in white cabbage leaves. Food Chem. 2016, 197 Pt A, 47–56. [Google Scholar] [CrossRef]
- Hang, Y.D.; Woodams, E.E. Lipase production by Geotrichum candidum from sauerkraut brine. World J. Microbiol. Biotechnol. 1990, 6, 418–421. [Google Scholar] [CrossRef]
- Ku, M.A.; Hang, Y.D. Effect of inulin on yeast inulinase production in sauerkraut brine. World J. Microbiol. Biotechnol. 1994, 10, 354–355. [Google Scholar] [CrossRef]
- Satora, P.; Skotniczny, M.; Strnad, S.; Piechowicz, W. Chemical composition of sauerkraut produced of different cabbage varieties. LWT-Food Sci. Technol. 2021, 136, 110325. [Google Scholar] [CrossRef]
- Kiai, H.; Hafidi, A. Chemical composition changes in four green olive cultivars during spontaneous fermentation. LWT-Food Sci. Technol. 2014, 57, 663–670. [Google Scholar] [CrossRef]
- Cioch-Skoneczny, M.; Satora, P.; Skotniczny, M.; Skoneczny, S. Quantitative and qualitative composition of yeast microbiota in spontaneously fermented grape musts obtained from cool climate grape varieties ‘Rondo’ and ‘Regent’. FEMS Yeast Res. 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Breuer, U.; Harms, H. Debaryomyces hansenii—An extremophilic yeast with biotechnological potential. Yeast 2006, 23, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Masoud, W.; Jakobsen, M. Surface ripened cheeses: The effects of Debaryomyces hansenii, NaCl and pH on the intensity of pigmentation produced by Brevibacterium linens and Corynebacterium flavescens. Int. Dairy J. 2003, 13, 231–237. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Yeasts. In Fungi and Food Spoilage; Pitt, J.I., Hocking, A.D., Eds.; Springer: Boston, MA, USA, 2009; pp. 357–382. [Google Scholar]
- Ramos, J.; Melero, Y.; Ramos-Moreno, L.; MichÁn, C.; Cabezas, L. Debaryomyces hansenii strains from valle de los pedroches iberian dry meat products: Isolation, identification, characterization, and selection for starter cultures. J. Microbiol. Biotechnol. 2017, 27, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deak, T.; Beuchat, L.R. Handbook of Food Spoilage Yeasts; CRC Press: New York, NY, USA, 1996. [Google Scholar]
- Calahorra, M.; Sánchez, N.S.; Peña, A. Activation of fermentation by salts in Debaryomyces hansenii. FEMS Yeast Res. 2009, 9, 1293–1301. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Brito, D.; Magaña-Alvarez, A.; Lappe-Oliveras, P.; Cortes-Velazquez, A.; Torres-Calzada, C.; Herrera-Suarez, T.; Larqué-Saavedra, A.; Tapia-Tussell, R. Genetic diversity of Clavispora lusitaniae isolated from Agave fourcroydes Lem, as revealed by DNA fingerprinting. J. Microbiol. 2015, 53, 14–20. [Google Scholar] [CrossRef]
- El-Sharoud, W.M.; Belloch, C.; Peris, D.; Querol, A. Molecular identification of yeasts associated with traditional Egyptian dairy products. J. Food Sci. 2009, 74, M341–M346. [Google Scholar] [CrossRef]
- Kaminarides, S.E.; Anifantakis, E.M. Evolution of the microflora of Kopanisti cheese during ripening. Study of the yeast flora. Le Lait INRA Ed. 1989, 69, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Boekhout, T. Yeasts in Food: Beneficial and Detrimental Aspects; Behr’s Verlag: Hamburg, Germany, 2003. [Google Scholar]
- Romo-Sánchez, S.; Alves-Baffi, M.; Arévalo-Villena, M.; Ubeda-Iranzo, J.; Briones-Pérez, A. Yeast biodiversity from oleic ecosystems: Study of their biotechnological properties. Food Microbiol. 2010, 27, 487–492. [Google Scholar] [CrossRef]
- Satora, P.; Celej, D.; Skotniczny, M.; Trojan, N. Identyfikacja drożdży obecnych w kiszonej kapuście komercyjnej i otrzymywanej w gospodarstwach rolnych. ŻYWNOŚĆ Nauka Technol. Jakość 2017, 4, 27–36. [Google Scholar]
- Cano-García, L.; Flores, M.; Belloch, C. Molecular characterization and aromatic potential of Debaryomyces hansenii strains isolated from naturally fermented sausages. Food Res. Int. 2013, 52, 42–49. [Google Scholar] [CrossRef]
- Amorim, J.C.; Piccoli, R.H.; Duarte, W.F. Probiotic potential of yeasts isolated from pineapple and their use in the elaboration of potentially functional fermented beverages. Food Res. Int. 2018, 107, 518–527. [Google Scholar] [CrossRef]
- Hassan, R.; Sand, M.I.; El-Kadi, S.M. Effect of some organic acids on fungal growth and their toxins production. J. Agric. Chem. Biotechn. Mansoura Univ. 2012, 3, 391–397. [Google Scholar] [CrossRef]
- Satora, P.; Tuszynski, T. Biodiversity of yeasts during plum Wegierka Zwykla spontaneous fermentation. Food Technol. Biotechnol. 2005, 43, 277–282. [Google Scholar]
- Al-Qaysi, S.A.S.; Al-Haideri, H.; Thabit, Z.A.; Al-Kubaisy, W.H.A.A.-R.; Ibrahim, J.A.A.-R. Production, Characterization, and Antimicrobial Activity of Mycocin Produced by Debaryomyces hansenii DSMZ70238. Int. J. Microbiol. 2017, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdaniewicz, M.; Satora, P.; Pater, A.; Bogacz, S. Low Lactic Acid-Producing Strain of Lachancea thermotolerans as a New Starter for Beer Production. Biomolecules 2020, 10, 256. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 5.8S-ITS rRNA Gene [bp] | Restriction Fragments [bp] | Species Identification by 5.8S‑ITS rRNA Gene Sequencing | Restriction Pattern | Accession No. | ||
|---|---|---|---|---|---|---|---|
| CfoI | HinfI | HaeIII | |||||
| I | 650 | 260 + 200 + 95 + 95 | 270 + 200 + 175 | 440 + 80 + 70 + 60 | Cryptococcus macerans | 1 | MK312605 |
| II | 650 | 590 | 310 + 310 | 570 + 80 | Nakazawaea holstii | 2 | MK312606 |
| III | 640 | 320 + 240 + 80 | 340 + 225 + 75 | 425 + 215 | Rhodotorula mucilaginosa | 3 | MK312607 |
| IV | 650 | 300 + 300 + 50 | 325 + 325 | 420 + 150 + 90 | Debaryomyces hansenii | 4 | MK312608 |
| V | 370 | 210 + 180 | 180 + 160 | 370 | Clavispora lusitaniae | 7 | MK312609 |
| VI | 625 | 300 + 265 + 60 | 320 + 300 | 400 + 115 + 90 | Meyerozyma guilliermondii | 8 | MK312610 |
| VII | 450 | 250 + 200 | 230 + 220 | 450 | Candida sake | 9 | MK312611 |
| VIII | 640 | 320 + 240 + 80 | 340 + 225 + 75 | 425 + 215 | Rhodotorula mucilaginosa | 11 | MK312612 |
| IX | 640 | 320 + 240 + 80 | 340 + 225 + 75 | 425 + 215 | Rhodotorula mucilaginosa | 14 | MK312613 |
| X | 370 | 210 + 180 | 180 + 160 | 370 | Clavispora lusitaniae | 15 | MK312614 |
| XI | 370 | 210 + 180 | 180 + 160 | 370 | Clavispora lusitaniae | 16 | MK312615 |
| XII | 650 | 300 + 300 + 50 | 325 + 325 | 420 + 150 + 90 | Debaryomyces hansenii | 17 | MK312616 |
| XIII | 370 | 210 + 180 | 180 + 160 | 370 | Clavispora lusitaniae | 18 | MK312617 |
| XIV | 450 | 170 + 100 + 100 + 80 | 250 + 200 | 340 + 80 + 30 | Pichia fermentans | 21 | MK312618 |
| XV | 650 | 300 + 300 + 50 | 325 + 325 | 420 + 150 + 90 | Debaryomyces hansenii | 22 | MK312619 |
| XVI | 650 | 300 + 300 + 50 | 325 + 325 | 420 + 150 + 90 | Debaryomyces hansenii | 23 | MK312620 |
| XVII | 500 | 300 + 100 | 300 + 150 | 500 | Tausonia pullulans | 25 | MK312621 |
| Isolate | Control (pH 5.6) | NaCl | Lactic Acid | pH | Killer Activity [mm] | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5% | 6% | 8% | 10% | 6 g L−1 | 8 g L−1 | 10 g L−1 | 3.6 | 3.4 | 3.2 | |||
| Cryptococcus macerans | ||||||||||||
| I | + + + + | + + + + | + + + | + + + | + | + + | + | - | + + | - | - | 16 |
| Nakazaweae holstii | ||||||||||||
| II | + + + + | + + | - | - | - | - | - | - | + + | - | - | - |
| Rhodotorula mucilaginosa | ||||||||||||
| III | + + + + | + + + + | + + + | + + + | - | + + | + + | - | + | - | - | - |
| VIII | + + + + | + + + | + + + | + + | + | + + | + | - | + + + | + | - | - |
| IX | + + + + | + + + + | + + + | + + + | + + | + | + | - | + | - | - | - |
| Debaryomyces hansenii | ||||||||||||
| IV | + + + + | + + + | + + + | + + + | + + | + + + | + + + | + + | + + | - | - | 16 |
| XII | + + + + | + + + + | + + + + | + + + | + + | + + | - | - | + | - | - | 24 |
| XV | + + + + | + + + + | + + + + | + + + + | + + + | + | + | - | + | + | - | 14 |
| XVI | + + + + | + | - | - | - | + | - | - | + | - | - | 11 |
| Clavispora lusitaniae | ||||||||||||
| V | + + + + | + + + + | + + + | + + + | + + | + + + | + + | - | + + + | + | - | 22 |
| X | + + + + | + | - | - | - | + + | + | - | + | - | - | 16 |
| XI | + + + + | + + + + | + + + | + + | + | + + | + | - | + + | + | - | 6 |
| XIII | + + + + | + + + + | + + + | + + | + + | + + | + | - | + + | - | - | - |
| Meyerozyma guilliermondii | ||||||||||||
| VI | + + + + | + + + + | + + + | + + + | + + + | + + | + + | - | + + + | + + | - | - |
| Candida sake | ||||||||||||
| VII | + + + + | + + + + | + + + | + + | + | + + | + + | - | + + + | + | - | 22 |
| Pichia fermentans | ||||||||||||
| XIV | + + + + | + | - | - | - | + | + | - | + | + | - | 18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satora, P.; Skotniczny, M.; Strnad, S.; Ženišová, K. Yeast Microbiota during Sauerkraut Fermentation and Its Characteristics. Int. J. Mol. Sci. 2020, 21, 9699. https://doi.org/10.3390/ijms21249699
Satora P, Skotniczny M, Strnad S, Ženišová K. Yeast Microbiota during Sauerkraut Fermentation and Its Characteristics. International Journal of Molecular Sciences. 2020; 21(24):9699. https://doi.org/10.3390/ijms21249699
Chicago/Turabian StyleSatora, Paweł, Magdalena Skotniczny, Szymon Strnad, and Katarína Ženišová. 2020. "Yeast Microbiota during Sauerkraut Fermentation and Its Characteristics" International Journal of Molecular Sciences 21, no. 24: 9699. https://doi.org/10.3390/ijms21249699
APA StyleSatora, P., Skotniczny, M., Strnad, S., & Ženišová, K. (2020). Yeast Microbiota during Sauerkraut Fermentation and Its Characteristics. International Journal of Molecular Sciences, 21(24), 9699. https://doi.org/10.3390/ijms21249699