Metabolism of Melatonin Synthesis-Related Indoles in the Turkey Pineal Organ and Its Modification by Monochromatic Light



Abstract
:1. Introduction
2. Results
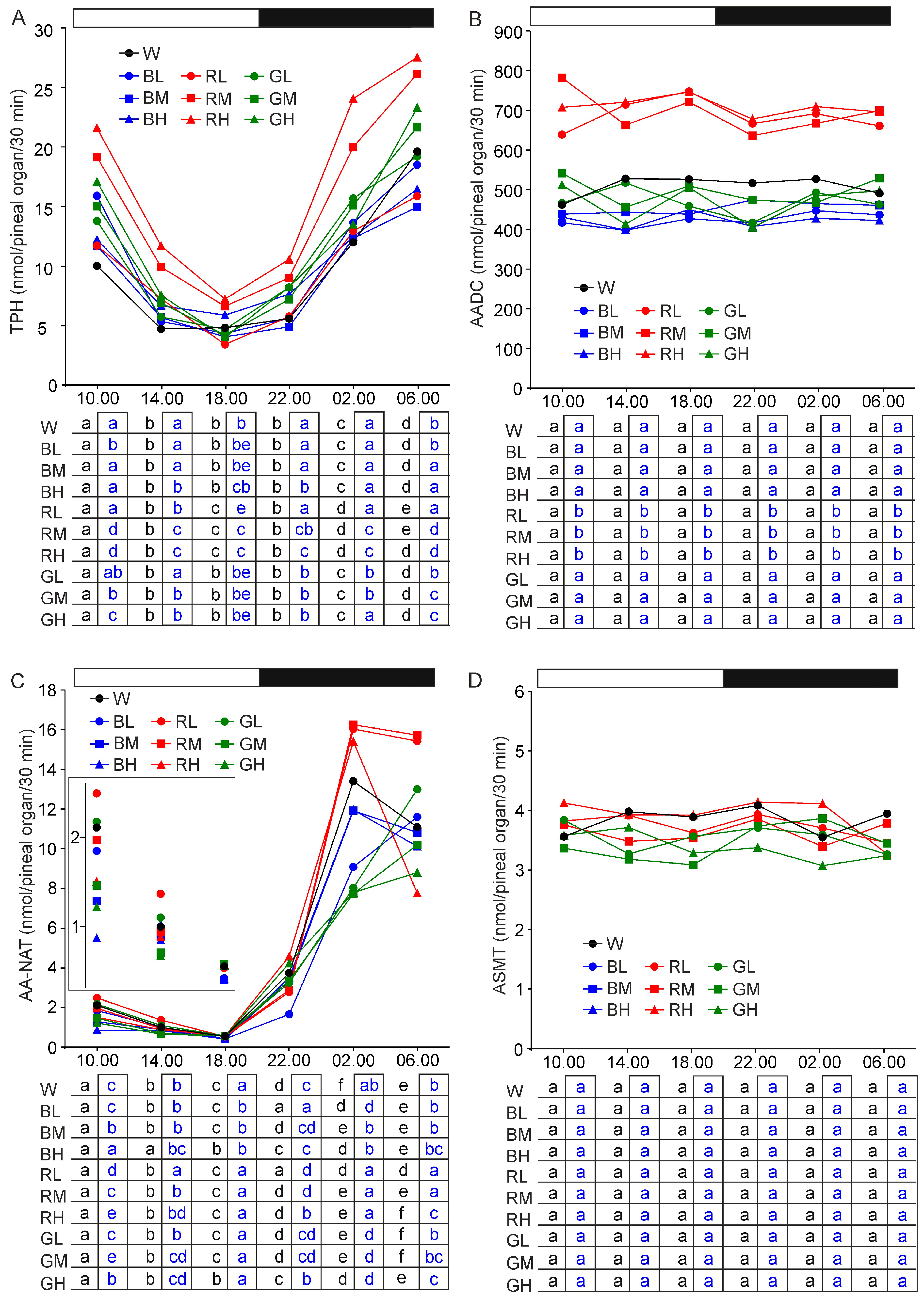
2.1. Activities of Melatonin Synthesis Pathway Enzymes
2.1.1. Tryptophan Hydroxylase
2.1.2. Aromatic Amino Acid Decarboxylase
2.1.3. Arylalkylamine N-Acetyltransferase
2.1.4. N-Acetylserotonin O-Methyltransferase
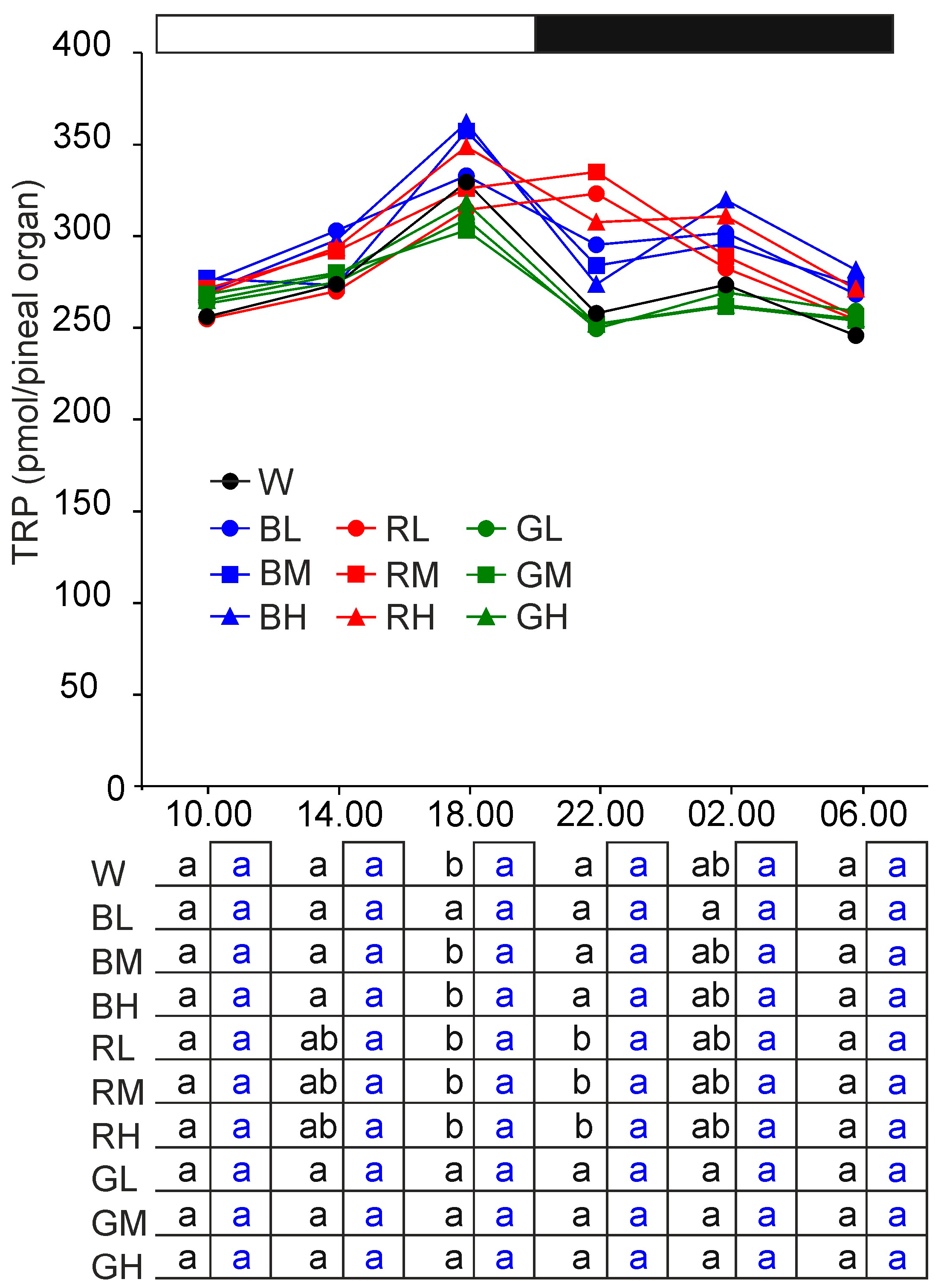
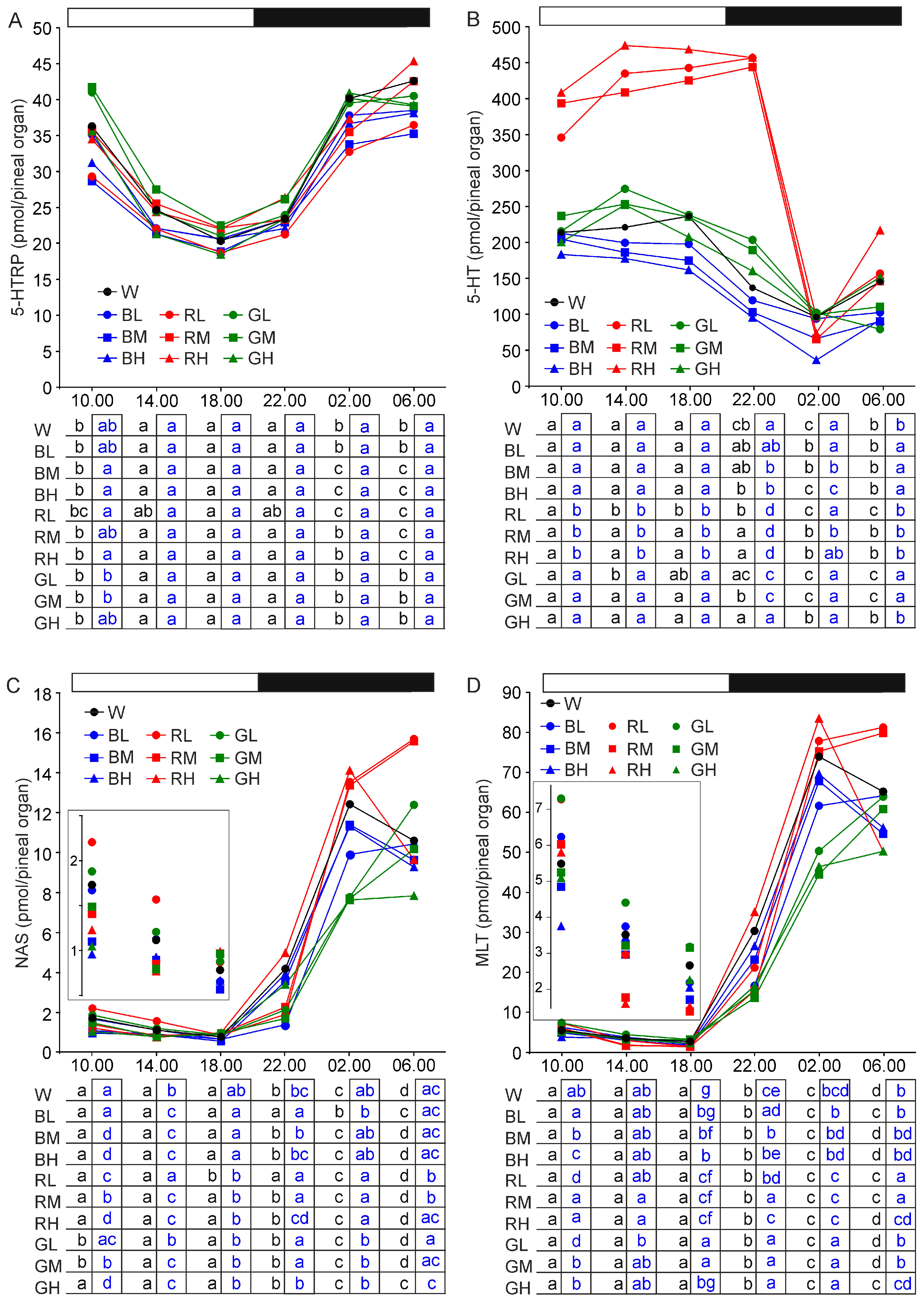
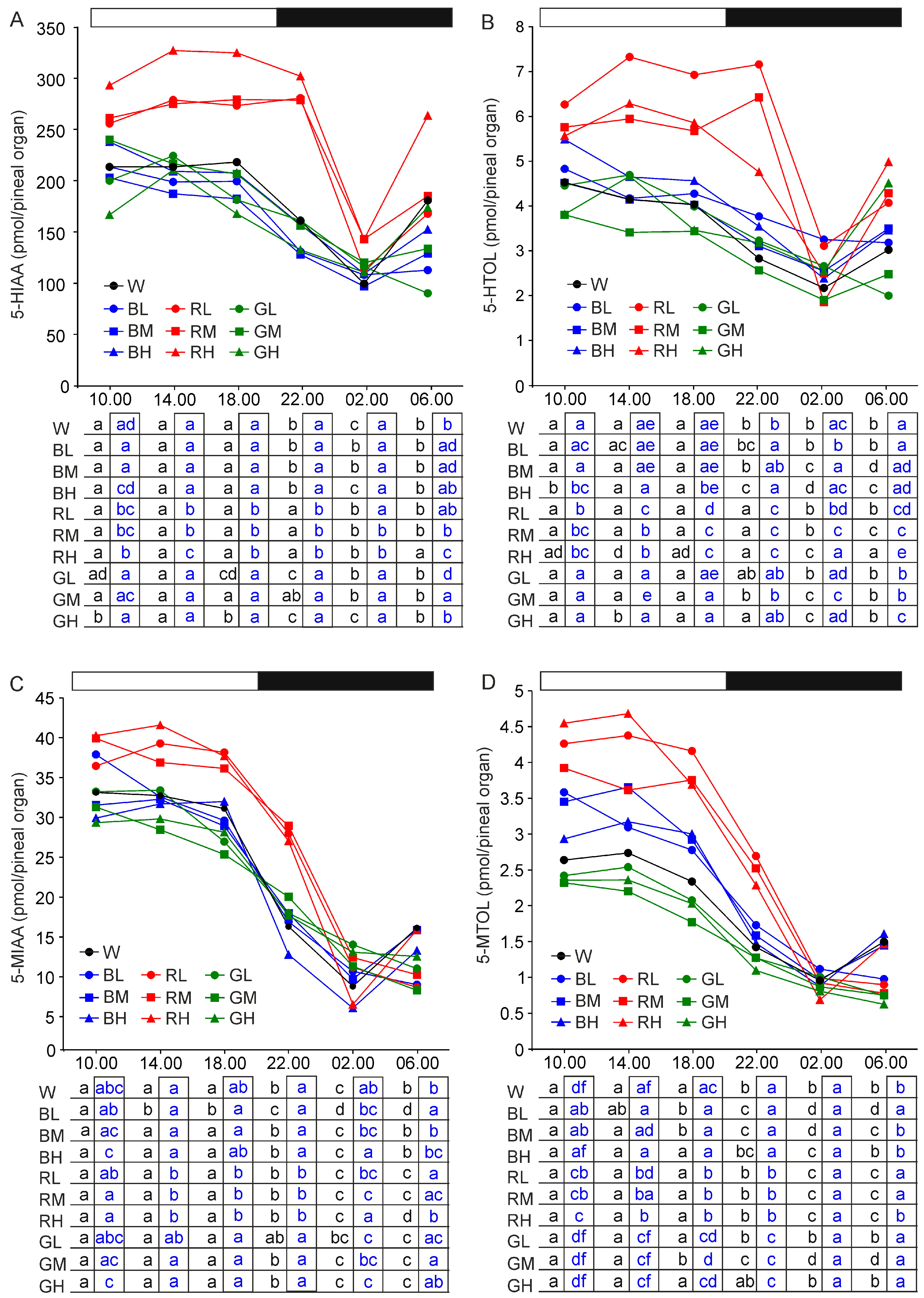
2.2. Content of Melatonin Synthesis-Related Indoles
2.2.1. Tryptophan
2.2.2. 5-Hydroxytryptophan
2.2.3. Serotonin
2.2.4. N-Acetylserotonin
2.2.5. Melatonin
2.2.6. 5-Hydroxyindoleacetic Acid and 5-Hydroxytryptophol
2.2.7. 5-Methoxyindoleacetic Acid and 5-Methoxytryptophol
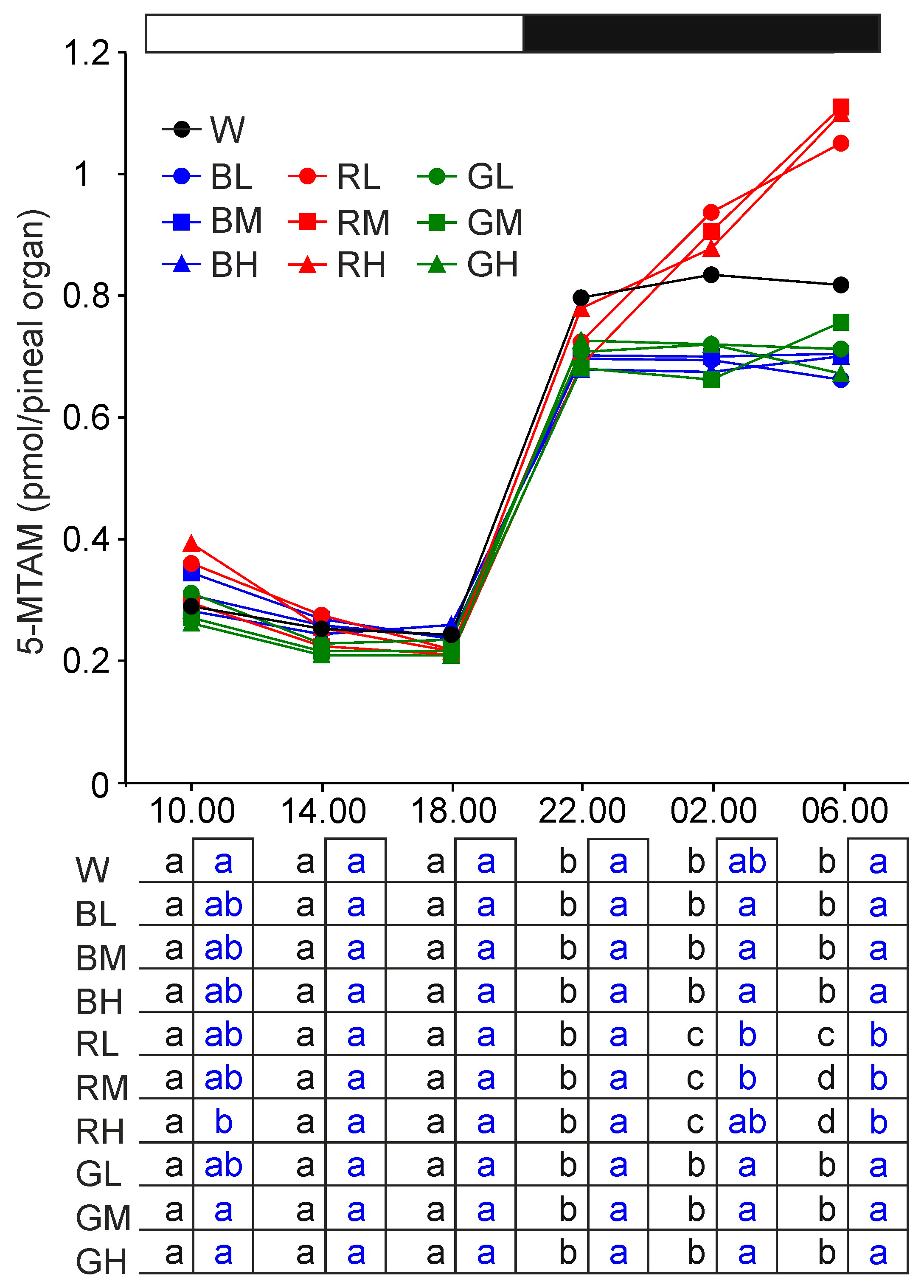
2.2.8. 5-Methoxytryptamine and 5-Methoxytryptophan
3. Discussion
3.1. Daily Rhythmicity in the Activities of Enzymes Governing Indole Metabolism in the Turkey Pineal Organ
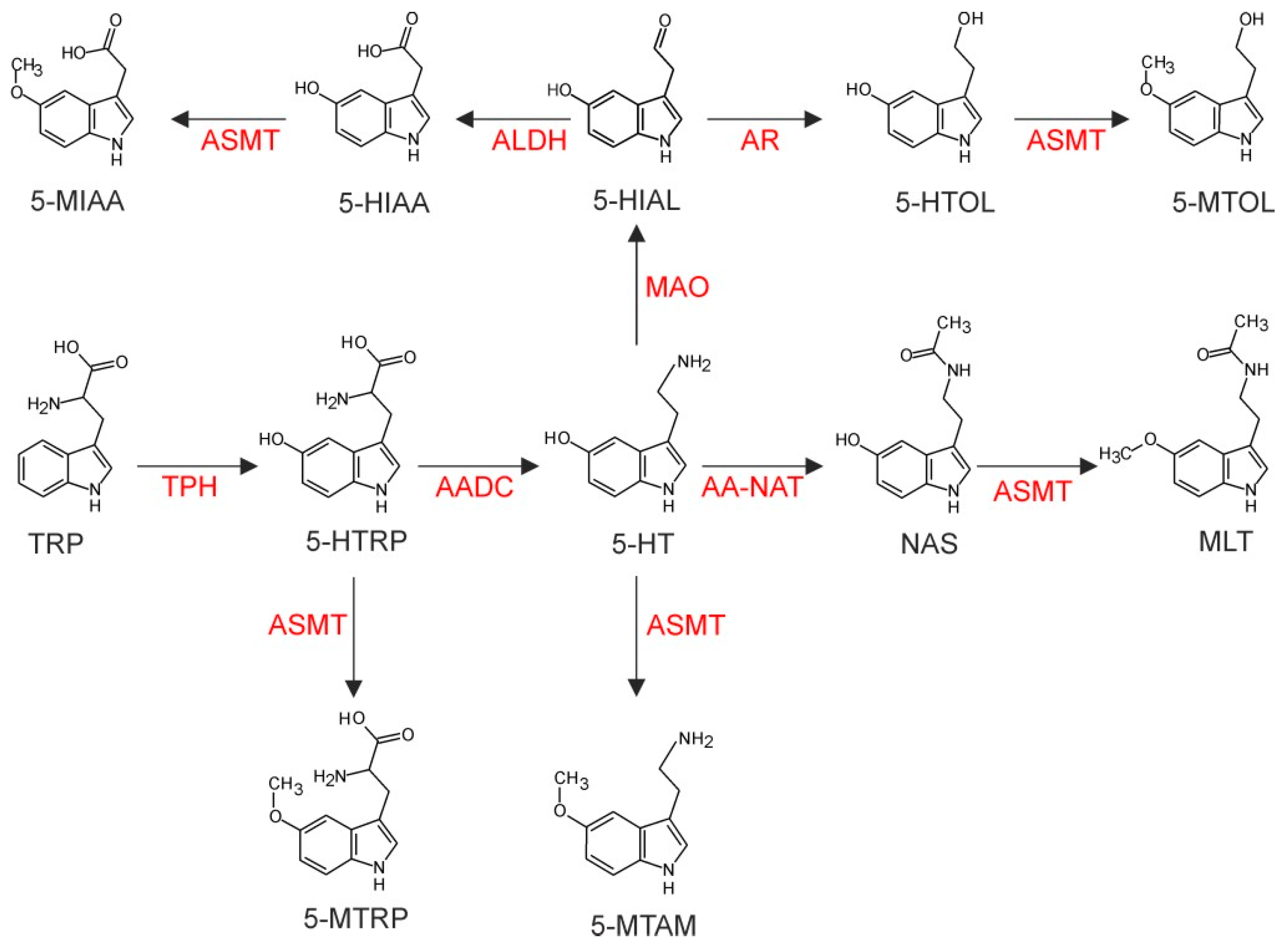
3.2. Metabolic Profile of Pineal Indoles
3.3. Effect of Monochromatic Light on Pineal Indole Metabolism
4. Materials and Methods
4.1. Birds and Experimental Design
4.2. Assay of Enzyme Activity and Indole Content
4.2.1. Chemicals
4.2.2. Sample Preparation for the Assays of Enzyme Activity and Indole Content
4.2.3. Content of Melatonin-Synthesis Related Indoles
4.2.4. Measurement of the Melatonin-Synthesis Pathway Enzymes Activities
Tryptophan Hydroxylase Activity
Aromatic L-Amino Acid Decarboxylase Activity
Arylalkylamine N-Acetyltransferase Activity
N-Acetylserotonin O-Methyltransferase Activity
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-HIAA | 5-hydroxyindoleacetic acid |
| 5-HT | serotonin |
| 5-HTOL | 5-hydroxytryptophol |
| 5-HTRP | 5-hydroxytryptophan |
| 5-MIAA | 5-methoxyindoleacetic acid |
| 5-MTAM | 5-methoxytryptamine |
| 5-MTOL | 5-methoxytryptophol |
| 5-MTRP | 5-methoxytryptophan |
| AADC | aromatic L-amino acid decarboxylase |
| AA-NAT | arylalkylamine N-acetyltransferase |
| ASMT | N-acetylserotonin O-methyltransferase |
| HPLC | high pressure liquid chromatography |
| MLT | melatonin |
| NAS | N-acetylserotonin |
| TPH | tryptophan hydroxylase |
| TRP | tryptophan |
| BL | blue light 16 lx |
| BM | blue light 32 lx |
| BH | blue light 64 lx |
| GL | green light 16 lx |
| GM | green light 32 lx |
| GH | green light 64 lx |
| RL | red light 16 lx |
| RM | red light 32 lx |
| RH | red light 64 lx |
| W | white light 16 lx |
Appendix A

References
- Gwinner, E.; Zeman, M.; Klaassen, M. Synchronization by low-amplitude light-dark cycles of 24-hour pineal and plasma melatonin rhythms of hatchling European starlings (Sturnus vulgaris). J. Pineal Res. 1997, 23, 176–181. [Google Scholar] [CrossRef]
- Okano, T.; Takanaka, Y.; Nakamura, A.; Hirunagi, K.; Adachi, A.; Ebihara, S.; Fukada, Y. Immunocytochemical identification of pinopsin in the pineal glands of chicken and pigeon. Brain Res. Mol. Brain Res. 1997, 50, 190–196. [Google Scholar] [CrossRef]
- Petrusewicz-Kosińska, M.; Przybylska-Gornowicz, B.; Prusik, M.; Ziółkowska, N.; Lewczuk, B. Pinopsin and photoreception in the pineal organ of the domestic turkey during post-hatching development. Micron 2019, 126, 102749. [Google Scholar] [CrossRef]
- Panda, S.; Sato, T.K.; Castrucci, A.M.; Rollag, M.D.; DeGrip, W.J.; Hogenesch, J.B.; Provencio, I.; Kay, S.A. Melanopsin (Opn4) requirement for normal light-induced circadian phase shifting. Science 2002, 298, 2213–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, T.; Yoshizawa, T.; Fukada, Y. Pinopsin is a chicken pineal photoreceptive molecule. Nature 1994, 372, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Fukada, Y. Phototransduction cascade and circadian oscillator in chicken pineal gland. J. Pineal Res. 1997, 22, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holthues, H.; Engel, L.; Spessert, R.; Vollrath, L. Circadian gene expression patterns of melanopsin and pinopsin in the chick pineal gland. Biochem. Biophys. Res. Commun. 2005, 326, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Torii, M.; Kojima, D.; Okano, T. Two isoforms of chicken melanopsins show blue light sensitivity. FEBS Lett. 2007, 581, 5327–5331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooley, J.J.; Lu, J.; Chou, T.C.; Scammell, T.E.; Saper, C.B. Melanopsin in cells of origin of the retinohypothalamic tract. Nat. Neurosci. 2001, 4, 1165. [Google Scholar] [CrossRef]
- Hannibal, J. Neurotransmitters of the retino-hypothalamic tract. Cell Tissue Res. 2002, 309, 73–88. [Google Scholar] [CrossRef]
- Jia, L.; Wang, Z.; Chen, Y. Distribution and cytoarchitecture of sympathetic neurons innervating the pineal gland in chick: A CTB-HRP study. Anat. Histol. Embryol. 2009, 38, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Cassone, V.M.; Forsyth, A.M.; Woodlee, G.L. Hypothalamic regulation of circadian noradrenergic input to the chick pineal gland. J. Comp. Physiol. A Sens. Neural Bahav. Physiol. 1990, 167, 187–192. [Google Scholar] [CrossRef]
- Huber-Eicher, B.; Suter, A.; Spring-Stahli, P. Effects of colored light-emitting diode illumination on behavior and performance of laying hens. Poultry Sci. 2013, 92, 869–873. [Google Scholar] [CrossRef] [PubMed]
- El Halawani, M.F. Light intensity requirement for breeder hen turkeys. Gobbles 2009, 66, 6–7. [Google Scholar]
- Hassan, M.R.; Sultana, S.; Choe, H.S.; Ryu, K.S. Effect of monochromatic and combined light color on performance, blood parameters, ovarian, morphology, and reproductive hormones in laying hens. Italian J. Anim. Sci. 2013, 12, e56. [Google Scholar] [CrossRef] [Green Version]
- Rozenboim, I.; Biran, I.; Uni, Z.; Robinzon, B.; Halevy, O. The effect of monochromatic light on broiler growth and development. Poult. Sci. 1999, 78, 135–138. [Google Scholar] [CrossRef]
- Rozenboim, I.; Robinzon, B.; Rosenstrauch, A. Effect of light source and regimen on growing broilers. Br. Poult. Sci. 1999, 40, 452–457. [Google Scholar] [CrossRef]
- Halevy, O.; Biran, I.; Rozenboim, I. Various light source treatments affect body and skeletal muscle growth by affecting skeletal muscle satellite cell proliferation in broilers. Comp. Biochem. Physiol. A-Mol. Integr. Physiol. 1998, 120, 317–323. [Google Scholar] [CrossRef]
- Olanrewaju, H.A.; Purswell, J.L.; Maslin, W.R.; Collier, S.D.; Branton, S.L. Effects of color temperatures (kelvin) of LED bulbs on growth performance, carcass characteristics, and ocular development indices of broilers grown to heavy weights. Poult. Sci. 2015, 94, 338–344. [Google Scholar] [CrossRef]
- Rodenboog, H. Sodium, green, blue, cool or warm-white light? World Poult. 2001, 17, 22–23. [Google Scholar]
- Xie, D.; Wang, Z.X.; Dong, Y.L.; Cao, J.; Wang, J.F.; Chen, J.L.; Chen, Y.X. Effects of monochromatic light on immune response of broilers. Poult. Sci. 2008, 87, 1535–1539. [Google Scholar] [CrossRef] [PubMed]
- Rozenboim, I.; Biran, I.; Chaiseha, Y.; Yahav, S. The effect of a green and blue monochromatic light combination on broiler growth and development. Poult. Sci. 2004, 83, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, Y.; Pan, J.; Ying, Y.; Zhou, H. A new method to manipulate broiler chicken growth and metabolism: Response to mixed LED light system. Sci. Rep. 2016, 6, 25972. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Yang, Y.; Yang, B.; Yu, Y. Artificial Polychromatic Light Affects Growth and Physiology in Chicks. PLoS ONE 2014, 9, 113595. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Role of monochromatic light on daily variation of clock gene expression in the pineal gland of chick. J. Photochem. Photobiol. B 2016, 164, 57–64. [Google Scholar] [CrossRef]
- Ma, S.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Effect of Monochromatic Light on Circadian Rhythm of Clock Genes in Chick Pinealocytes. Photochem. Photobiol. 2018, 94, 1263–1272. [Google Scholar] [CrossRef]
- Ma, S.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. BMAL1 but not CLOCK is associated with monochromatic green light-induced circadian rhythm of melatonin in chick pinealocytes. Endocr Connect 2019, 8, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Prusik, M.; Lewczuk, B. Roles of Direct Photoreception and the Internal Circadian Oscillator in the Regulation of Melatonin Secretion in the Pineal Organ of the Domestic Turkey: A Novel In Vitro Clock and Calendar Model. Int. J. Mol. Sci. 2019, 20, 4022. [Google Scholar] [CrossRef] [Green Version]
- Prusik, M.; Lewczuk, B. Diurnal Rhythm of Plasma Melatonin Concentration in the Domestic Turkey and Its Regulation by Light and Endogenous Oscillators. Animals 2020, 10, 678. [Google Scholar] [CrossRef]
- Ohshima, K.; Matsuo, S. Functional morphology of the pineal gland in young chickens. Anat. Anz. 1984, 156, 407–418. [Google Scholar]
- Przybylska-Gornowicz, B.; Lewczuk, B.; Prusik, M.; Nowicki, M. Post-hatching development of the turkey pineal organ: Histological and immunohistochemical studies. Neuro Endocrinol. Lett. 2005, 26, 383–392. [Google Scholar] [PubMed]
- Przybylska-Gornowicz, B.; Lewczuk, B.; Prusik, M.; Bulc, M. Pineal concretions in Turkey (Meleagris gallopavo) as a results of collagen mediated calcification. Histol. Histopathol. 2009, 24, 407–415. [Google Scholar] [PubMed]
- Petrusewicz-Kosińska, M.; Przybylska-Gornowicz, B.; Ziółkowska, N.; Martyniuk, K.; Lewczuk, B. Developmental morphology of the turkey pineal organ. Immunocytochemical and ultrastructural studies. Micron 2019, 122, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Adamska, I.; Lewczuk, B.; Markowska, M.; Majewski, P.M. Daily profiles of melatonin synthesis-related indoles in the pineal glands of young chickens (Gallus gallus domesticus L.). J. Photochem. Photobiol. B. 2016, 164, 335–343. [Google Scholar] [CrossRef]
- Lewczuk, B.; Ziółkowska, N.; Prusik, M.; Przybylska-Gornowicz, B. Diurnal profiles of melatonin synthesis-related indoles, catecholamines and their metabolites in the duck pineal organ. Int. J. Mol. Sci. 2014, 15, 12604–12630. [Google Scholar] [CrossRef] [Green Version]
- Ziółkowska, N.; Lewczuk, B.; Prusik, M. Diurnal and circadian variations in indole contents in the goose pineal gland. Chronobiol. Int. 2018, 35, 1560–1575. [Google Scholar] [CrossRef]
- Hanuszewska, M.; Prusik, M.; Lewczuk, B. Embryonic Ontogeny of 5-Hydroxyindoles and 5-Methoxyindoles Synthesis Pathways in the Goose Pineal Organ. Int. J. Mol. Sci. 2019, 20, 3948. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Borjigin, J. N-acetyltransferase is not the rate limiting enzyme of melatonin synthesis at night. J. Pineal Res. 2005, 39, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Chattoraj, A.; Liu, T.; Zhang, L.S.; Huang, Z.; Borjigin, J. Melatonin formation in mammals: In vivo perspectives. Rev. Endocr. Metab. Disord. 2009, 10, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Yadav, G.; Malik, S.; Rani, S.; Kumar, V. Role of light wavelengths in synchronization of circadian physiology in songbirds. Physiol. Behav. 2015, 140, 164–171. [Google Scholar] [CrossRef]
- Rumanova, V.S.; Okuliarova, M.; Zeman, M. Differential Effects of Constant Light and Dim Light at Night on the Circadian Control of Metabolism and Behavior. Int. J. Mol. Sci. 2020, 21, 5478. [Google Scholar] [CrossRef] [PubMed]
- Csernus, V.; Becher, P.; Mess, B. Wavelength dependency of light-induced changes in rhythmic melatonin secretion from chicken pineal gland in vitro. Neuro Endocrinol. Lett. 1999, 20, 299–304. [Google Scholar] [PubMed]
- Sato, K.; Yamashita, T.; Kojima, K.; Sakai, K.; Matsutani, Y.; Yanagawa, M.; Yamano, Y.; Wada, A.; Iwabe, N.; Ohuchi, H.; et al. Pinopsin evolved as the ancestral dim-light visual opsin in vertebrates. Commun. Biol. 2018, 1, 156. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Wang, Z.; Dong, Y.; Cao, J.; Chen, Y. Effect of monochromatic light on the circadian clock of cultured chick retinal tissue. Exp. Eye Res. 2020, 194, 108008. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Q.; Wang, T.; Pan, J. Wavelength-specific artificial light disrupts molecular clock in avian species: A power-calibrated statistical approach. Environ. Pollut. 2020, 265, 114206. [Google Scholar] [CrossRef]
- Jin, E.; Jia, L.; Li, J.; Yang, G.; Wang, Z.; Cao, J.; Chen, Y. Effect of monochromatic light on melatonin secretion and arylalkylamine N-acetyltransferase mRNA expression in the retina and pineal gland of broilers. Anat. Rec. 2011, 294, 1233–1241. [Google Scholar] [CrossRef]
- Drozdova, A.; Okuliarova, M.; Zeman, M. The effect of different wavelengths of light during incubation on the development of rhythmic pineal melatonin biosynthesis in chick embryos. Animal 2019, 13, 1635–1640. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Cao, J.; Wang, Z.; Dong, Y.; Chen, Y. Effect of monochromatic light on the temporal expression of N-acetyltransferase in chick pineal gland. Chronobiol. Int. 2020, 37, 1140–1150. [Google Scholar] [CrossRef]
- Grubisic, M.; Haim, A.; Bhusal, P.; Dominoni, D.M.; Gabriel, K.M.A.; Jechow, A.; Kupprat, F.; Lerner, A.; Marchant, P.; Riley, W.; et al. Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates. Sustainability 2019, 11, 6400. [Google Scholar] [CrossRef] [Green Version]
- Barber, C.L.; Prescott, N.B.; Jarvis, J.R.; Le Seuer, C.; Perry, G.C.; Wathes, D.M. Comparative study of the photopic spectral sensitivity of domestic ducks (Anas platyrhynchos domesticus), turkeys (Meleagris gallopavo gallopavo) and humans. Br. Poult. Sci. 2006, 47, 365–374. [Google Scholar] [CrossRef]
- Lewis, P.D.; Morris, T.R. Poultry Lighting: The Theory and Practice; Northcot: Andover, UK, 2006; p. 176. [Google Scholar]
- Mohammed, H.H. Assessment of The role of Light in Welfare of Layers. SVU-Int. J. Vet. Sci. 2019, 2, 36–50. [Google Scholar] [CrossRef]
- Barbosa, R.; Scialfa, J.H.; Terra, I.M.; Cipolla-Neto, J.; Simonneaux, V.; Afeche, S.C. Tryptophan hydroxylase is modulated by L-type calcium channels in the rat pineal gland. Life Sci. 2008, 82, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.K.; Nagatsu, T.; Nagatsu, I.; Iizuka, R.; Narabayashi, H. Aromatic L-Animo decarboxylase activity in brains from normal human subjects and from patients with extrapyramidal diseases. Biomed. Res. 1981, 2, 560–566. [Google Scholar] [CrossRef] [Green Version]
- Bradford, K.T.; Zawilska, J.; Iuvone, P.M. Arylalkylamine (serotonin) N-acetyltransferase assay using high-performance liquid chromatography with fluorescence or electrochemical detection of N-acetyltryptamine. Anal. Biochem. 1990, 184, 228–234. [Google Scholar]
- Itoh, M.T.; Hattori, A.; Sumi, Y. Hydroxyindole-O-methyltransferase activity assay using high-performance liquid chromatography with fluorometric detection: Determination of melatonin enzymatically formed from N-acetylserotonin and S-adenosyl-l-methionine. J. Chromatogr. 1997, 692, 217–221. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part of the Experiment | Animal Room | Light Source | Light Intensity |
|---|---|---|---|
| Red color | RL | GOQ 3 LED Red Dominant wavelength: 620–626 nm Luminous flux of module 24 lm Luminous intensity of LED: 1600–2200 mcd Half angle ± 60° | 16 lx |
| RM | 32 lx | ||
| RH | 64 lx | ||
| W | Tungsten lamps, 3200 K | 16 lx | |
| Blue color | BL | GOQ 3 LED Blue Dominant wavelength: 450–460 nm Luminous flux of module 21 lm Luminous intensity of LED: 600–1200 mcd Half angle ± 60° | 16 lx |
| BM | 32 lx | ||
| BH | 64 lx | ||
| W | Tungsten lamps, 3200 K | 16 lx | |
| Green color | GL | GOQ 3 LED Green Dominant wavelength: 520–530 nm Luminous flux of module 48 lm Luminous intensity of LED: 3000–4000 mcd Half angle ± 60° | 16 lx |
| GM | 32 lx | ||
| GH | 64 lx | ||
| W | Tungsten lamps, 3200 K | 16 lx |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martyniuk, K.; Hanuszewska, M.; Lewczuk, B. Metabolism of Melatonin Synthesis-Related Indoles in the Turkey Pineal Organ and Its Modification by Monochromatic Light. Int. J. Mol. Sci. 2020, 21, 9750. https://doi.org/10.3390/ijms21249750
Martyniuk K, Hanuszewska M, Lewczuk B. Metabolism of Melatonin Synthesis-Related Indoles in the Turkey Pineal Organ and Its Modification by Monochromatic Light. International Journal of Molecular Sciences. 2020; 21(24):9750. https://doi.org/10.3390/ijms21249750
Chicago/Turabian StyleMartyniuk, Kamila, Maria Hanuszewska, and Bogdan Lewczuk. 2020. "Metabolism of Melatonin Synthesis-Related Indoles in the Turkey Pineal Organ and Its Modification by Monochromatic Light" International Journal of Molecular Sciences 21, no. 24: 9750. https://doi.org/10.3390/ijms21249750
APA StyleMartyniuk, K., Hanuszewska, M., & Lewczuk, B. (2020). Metabolism of Melatonin Synthesis-Related Indoles in the Turkey Pineal Organ and Its Modification by Monochromatic Light. International Journal of Molecular Sciences, 21(24), 9750. https://doi.org/10.3390/ijms21249750