Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Role of LOX in Liver Cancer
2.1. Prognostic Value and Biological Role of LOX in HCC
2.2. LOX and Angiogenesis
3. Roles of LOXL1 and LOXL3 in Liver Cancer
4. Role of LOXL2 in Liver Cancer
4.1. Prognostic Value and Biological Role of LOXL2 in HCC
4.2. LOXL2 in HCC Microenvironment
4.3. LOXL2 in CCA
5. Roles of LOXL4 in Tumor Microenvironment and Progression
6. Regulatory Pathways at Various Sub-Cellular Levels and the Impact of Genetic/Epigenetic Perturbations
7. Therapeutic Potential of Targeting Approaches on LOX Family Members
8. Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barker, H.E.; Cox, T.R.; Erler, J.T. The rationale for targeting the LOX family in cancer. Nat. Rev. Cancer 2012, 12, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Song, Y.; Pan, S.; Chu, M.; Wang, Z.-W.; Zhu, X. Evolving roles of lysyl oxidase family in tumorigenesis and cancer therapy. Pharmacol. Ther. 2020, 215, 107633. [Google Scholar] [CrossRef] [PubMed]
- López, B.; González, A.; Hermida, N.; Valencia, F.; De Teresa, E.; Díez, J. Role of lysyl oxidase in myocardial fibrosis: From basic science to clinical aspects. Am. J. Physiol. Circ. Physiol. 2010, 299, H1–H9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.-H.; Hsia, S.-M.; Shieh, T.-M. Lysyl Oxidase and the Tumor Microenvironment. Int. J. Mol. Sci. 2016, 18, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Q.; Ge, G. Lysyl Oxidase, Extracellular Matrix Remodeling and Cancer Metastasis. Cancer Microenviron. 2012, 5, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borel, A.; Eichenberger, D.; Farjanel, J.; Kessler, E.; Gleyzal, C.; Hulmes, D.J.S.; Sommer, P.; Font, B. Lysyl Oxidase-like Protein from Bovine Aorta. Isolation and maturation to an active form by bone morphogenetic protein-1. J. Biol. Chem. 2001, 276, 48944–48949. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Yang, A.; Chen, W.; Wang, P.; Liu, T.; Cong, M.; Xu, A.; Yan, X.; Jia, J.; You, H. Inhibition of lysyl oxidase-like 1 (LOXL1) expression arrests liver fibrosis progression in cirrhosis by reducing elastin crosslinking. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2018, 1864, 1129–1137. [Google Scholar] [CrossRef]
- Ikenaga, N.; Peng, Z.-W.; Vaid, K.A.; Liu, S.B.; Yoshida, S.; Sverdlov, D.Y.; Mikels-Vigdal, A.; Smith, V.; Schuppan, D.; Popov, Y.V. Selective targeting of lysyl oxidase-like 2 (LOXL2) suppresses hepatic fibrosis progression and accelerates its reversal. Gut 2017, 66, 1697–1708. [Google Scholar] [CrossRef] [Green Version]
- Dongiovanni, P.; Meroni, M.; Baselli, G.A.; Bassani, G.A.; Rametta, R.; Pietrelli, A.; Maggioni, M.; Facciotti, F.; Trunzo, V.; Badiali, S.; et al. Insulin resistance promotes Lysyl Oxidase Like 2 induction and fibrosis accumulation in non-alcoholic fatty liver disease. Clin. Sci. 2017, 131, 1301–1315. [Google Scholar] [CrossRef] [Green Version]
- Aumiller, V.; Strobel, B.; Romeike, M.; Schuler, M.; Stierstorfer, B.E.; Kreuz, S. Comparative analysis of lysyl oxidase (like) family members in pulmonary fibrosis. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Miller, B.W.; Morton, J.P.; Pinese, M.; Saturno, G.; Jamieson, N.B.; McGhee, E.; Timpson, P.; Leach, J.; McGarry, L.; Shanks, E.; et al. Targeting the LOX / hypoxia axis reverses many of the features that make pancreatic cancer deadly: Inhibition of LOX abrogates metastasis and enhances drug efficacy. EMBO Mol. Med. 2015, 7, 1063–1076. [Google Scholar] [CrossRef] [PubMed]
- Wilgus, M.-L.; Borczuk, A.C.; Stoopler, M.; Ginsburg, M.; Gorenstein, L.; Sonett, J.R.; Powell, C.A. Lysyl oxidase: A lung adenocarcinoma biomarker of invasion and survival. Cancer 2010, 117, 2186–2191. [Google Scholar] [CrossRef] [PubMed]
- Erler, J.T.; Bennewith, K.L.; Nicolau, M.; Dornhöfer, N.; Kong, C.; Le, Q.-T.; Chi, J.-T.A.; Jeffrey, S.S.; Giaccia, A.J. Lysyl oxidase is essential for hypoxia-induced metastasis. Nat. Cell Biol. 2006, 440, 1222–1226. [Google Scholar] [CrossRef] [PubMed]
- Tenti, P.; Vannucci, L. Lysyl oxidases: Linking structures and immunity in the tumor microenvironment. Cancer Immunol. Immunother. 2019, 69, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.Y.; Wang, N.; Zhang, C.; Chan, Y.T.; Yuen, M.F.; Feng, Y. LOXL4 Fosters an Immunosuppressive Microenvironment During Hepatocarcinogenesis. Hepatology 2020. [Google Scholar] [CrossRef]
- Kasashima, H.; Yashiro, M.; Kinoshita, H.; Fukuoka, T.; Morisaki, T.; Masuda, G.; Sakurai, K.; Kubo, N.; Ohira, M.; Hirakawa, K. Lysyl oxidase is associated with the epithelial–mesenchymal transition of gastric cancer cells in hypoxia. Gastric Cancer 2016, 19, 431–442. [Google Scholar] [CrossRef]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix Crosslinking Forces Tumor Progression by Enhancing Integrin Signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef] [Green Version]
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Abate, D.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdel-Rahman, O.; Abdelalim, A.; Abdoli, A.; Abdollahpour, I.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-Years for 29 Cancer Groups, 1990 to 2017: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2019, 5, 1749–1768. [Google Scholar] [CrossRef] [Green Version]
- Nault, J.; Bioulac–Sage, P.; Zucman–Rossi, J. Hepatocellular Benign Tumors—From Molecular Classification to Personalized Clinical Care. Gastroenterology 2013, 144, 888–902. [Google Scholar] [CrossRef]
- World Health Organization. Projections of Mortality and Causes of Death, 2016 to 2060. Available online: http://www.who.int/healthinfo/global_burden_disease/projections/en/ (accessed on 4 November 2020).
- Tahmasebi-Birgani, M.; Carloni, V. Tumor Microenvironment, a Paradigm in Hepatocellular Carcinoma Progression and Therapy. Int. J. Mol. Sci. 2017, 18, 405. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Huang, S.; Wu, G.; Huang, C.; Li, X.; Chen, Z.; Zhao, L.; Zhao, Y. Lysyl Oxidase Is Predictive of Unfavorable Outcomes and Essential for Regulation of Vascular Endothelial Growth Factor in Hepatocellular Carcinoma. Dig. Dis. Sci. 2015, 60, 3019–3031. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Liu, J.; Wang, F.; Tian, Z.; Ma, B.; Li, Z.; Wang, B.; Zhao, W. Lysyl oxidase assists tumor-initiating cells to enhance angiogenesis in hepatocellular carcinoma. Int. J. Oncol. 2019, 54, 1398–1408. [Google Scholar] [CrossRef] [PubMed]
- Umezaki, N.; Nakagawa, S.; Yamashita, Y.; Kitano, Y.; Arima, K.; Miyata, T.; Hiyoshi, Y.; Okabe, H.; Nitta, H.; Hayashi, H.; et al. Lysyl oxidase induces epithelial-mesenchymal transition and predicts intrahepatic metastasis of hepatocellular carcinoma. Cancer Sci. 2019, 110, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Erler, J.T.; Giaccia, A.J. Lysyl Oxidase Mediates Hypoxic Control of Metastasis: Figure 1. Cancer Res. 2006, 66, 10238–10241. [Google Scholar] [CrossRef] [Green Version]
- Triantafyllou, E.-A.; Mylonis, I.; Simos, G.; Paraskeva, E. Hypoxia Induces Pro-Fibrotic and Fibrosis Marker Genes in Hepatocellular Carcinoma Cells Independently of Inflammatory Stimulation and the NF-κΒ Pathway. Hypoxia 2019, 7, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Wang, V.; Davis, D.A.; Yarchoan, R. Identification of functional hypoxia inducible factor response elements in the human lysyl oxidase gene promoter. Biochem. Biophys. Res. Commun. 2017, 490, 480–485. [Google Scholar] [CrossRef]
- Wang, V.; Davis, D.A.; Haque, M.; Huang, L.E.; Yarchoan, R. Differential Gene Up-Regulation by Hypoxia-Inducible Factor-1α and Hypoxia-Inducible Factor-2α in HEK293T Cells. Cancer Res. 2005, 65, 3299–3306. [Google Scholar] [CrossRef] [Green Version]
- Tse, A.P.-W.; Sze, K.M.-F.; Shea, Q.T.-K.; Chiu, E.Y.-T.; Tsang, F.H.-C.; Chiu, D.K.-C.; Zhang, M.S.; Lee, D.; Xu, I.M.-J.; Chan, C.Y.-K.; et al. Hepatitis transactivator protein X promotes extracellular matrix modification through HIF/LOX pathway in liver cancer. Oncogenesis 2018, 7, 44. [Google Scholar] [CrossRef]
- Huang, C.-S.; Ho, C.-T.; Tu, S.-H.; Pan, M.-H.; Chang, H.-W.; Wu, C.-H.; Ho, Y.-S.; Chang, C.-H.; Chuang, C.-H. Long-Term Ethanol Exposure-Induced Hepatocellular Carcinoma Cell Migration and Invasion through Lysyl Oxidase Activation Are Attenuated by Combined Treatment with Pterostilbene and Curcumin Analogues. J. Agric. Food Chem. 2013, 61, 4326–4335. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Wang, H.; Yan, W.; Zhang, Q.; Chang, X. Expression of the lysyl oxidase propeptide in hepatocellular carcinoma and its clinical relevance. Oncol. Rep. 2014, 31, 1669–1676. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Njah, K.; Hong, W. Agrin Mediates Angiogenesis in the Tumor Microenvironment. Trends Cancer 2020, 6, 81–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Ding, J.; Zhu, H.; Jing, Y.; Zhou, H.; Tian, H.; Tang, K.; Wang, G.; Wang, X. LOXL1 confers antiapoptosis and promotes gliomagenesis through stabilizing BAG2. Cell Death Differ. 2020, 27, 3021–3036. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Masuda, T.; Kuramitsu, S.; Tobo, T.; Sato, K.; Kidogami, S.; Nambara, S.; Ueda, M.; Tsuruda, Y.; Kuroda, Y.; et al. Potential association of LOXL1 with peritoneal dissemination in gastric cancer possibly via promotion of EMT. PLoS ONE 2020, 15, e0241140. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, J.; Wang, Y.; Wu, L.; Wu, C.; Mao, B.; Prasad, E.M.; Wang, Y.; Chin, Y.E. LOXL1 modulates the malignant progression of colorectal cancer by inhibiting the transcriptional activity of YAP. Cell Commun. Signal. 2020, 18, 1–16. [Google Scholar] [CrossRef]
- Le Calvé, B.; Griveau, A.; Vindrieux, D.; Maréchal, R.; Wiel, C.; Svrcek, M.; Gout, J.; Azzi-Martin, L.; Payen, L.; Cros, J.; et al. Lysyl oxidase family activity promotes resistance of pancreatic ductal adenocarcinoma to chemotherapy by limiting the intratumoral anticancer drug distribution. Oncotarget 2016, 7, 32100–32112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, A.G.; Eivers, S.B.; Dervan, E.W.; O’Brien, C.J.; Wallace, D.M. Lysyl Oxidase Like 1: Biological roles and regulation. Exp. Eye Res. 2020, 193, 107975. [Google Scholar] [CrossRef] [PubMed]
- Kasashima, H.; Yashiro, M.; Okuno, T.; Miki, Y.; Kitayama, K.; Masuda, G.; Kinoshita, H.; Morisaki, T.; Fukuoka, T.; Hasegawa, T.; et al. Significance of the Lysyl Oxidase Members Lysyl Oxidase Like 1, 3, and 4 in Gastric Cancer. Digesiton 2018, 98, 238–248. [Google Scholar] [CrossRef]
- Ren, J.; Wang, X.; Wei, G.; Meng, Y. Exposure to desflurane anesthesia confers colorectal cancer cells metastatic capacity through deregulation of miR-34a/LOXL3. Eur. J. Cancer Prev. 2020. [Google Scholar] [CrossRef]
- Laurentino, T.D.S.; Soares, R.D.S.; Marie, S.; Oba-Shinjo, S.M. LOXL3 Function beyond Amino Oxidase and Role in Pathologies, Including Cancer. Int. J. Mol. Sci. 2019, 20, 3587. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Zhou, J.; Gao, Y.; Pan, S.; Zhu, X. The prognostic value of the lysyl oxidase family in ovarian cancer. J. Clin. Lab. Anal. 2020, e23538. [Google Scholar] [CrossRef]
- Wong, C.C.-L.; Tse, A.P.-W.; Huang, Y.-P.; Zhu, Y.-T.; Chiu, D.K.-C.; Lai, R.K.-H.; Au, S.L.-K.; Kai, A.K.-L.; Lee, J.M.-F.; Wei, L.L.; et al. Lysyl oxidase-like 2 is critical to tumor microenvironment and metastatic niche formation in hepatocellular carcinoma. Hepatology 2014, 60, 1645–1658. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, X.; Zhu, D.; Liu, T.; Liang, X.; Liu, F.; Zhang, Y.; Dong, X.; Sun, B. HIF-1α promoted vasculogenic mimicry formation in hepatocellular carcinoma through LOXL2 up-regulation in hypoxic tumor microenvironment. J. Exp. Clin. Cancer Res. 2017, 36, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Chung, T.; Rhee, H.; Kim, Y.-J.; Jeon, Y.; Yoo, J.E.; Noh, S.; Han, D.H.; Park, Y.N. Increased Expression of the Matrix-Modifying Enzyme Lysyl Oxidase-Like 2 in Aggressive Hepatocellular Carcinoma with Poor Prognosis. Gut Liver 2019, 13, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.; Zhao, X.; Liu, T.; Zhang, Y.; Sun, R.; Dong, X.; Liu, F.; Zhao, N.; Zhang, D.; Wu, L.; et al. LOXL2 promotes vasculogenic mimicry and tumour aggressiveness in hepatocellular carcinoma. J. Cell. Mol. Med. 2019, 23, 1363–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninomiya, G.; Yamada, S.; Hayashi, M.; Takeda, S.; Suenaga, M.; Takami, H.; Kanda, M.; Iwata, N.; Niwa, Y.; Tanaka, C.; et al. Significance of Lysyl oxidase-like 2 gene expression on the epithelial-mesenchymal status of hepatocellular carcinoma. Oncol. Rep. 2018, 39, 2664–2672. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Y.; Zhu, Y.; Cong, Q.; Xiang, Y.; Fu, L. The effect of LOXL2 in hepatocellular carcinoma. Mol. Med. Rep. 2016, 14, 1923–1932. [Google Scholar] [CrossRef] [Green Version]
- Ezzoukhry, Z.; Henriet, E.; Piquet, L.; Boyé, K.; Bioulac-Sage, P.; Balabaud, C.; Couchy, G.; Zucman-Rossi, J.; Moreau, V.; Saltel, F. TGF-β1 promotes linear invadosome formation in hepatocellular carcinoma cells, through DDR1 up-regulation and collagen I cross-linking. Eur. J. Cell Biol. 2016, 95, 503–512. [Google Scholar] [CrossRef]
- Fan, Z.; Zheng, W.; Li, H.; Wu, W.; Liu, X.; Sun, Z.; Hu, H.; Du, L.; Jia, Q.; Liu, Q. LOXL2 upregulates hypoxia-inducible factor-1α signaling through Snail-FBP1 axis in hepatocellular carcinoma cells. Oncol. Rep. 2020, 43, 1641–1649. [Google Scholar] [CrossRef]
- Xing, X.; Wang, Y.; Zhang, X.; Gao, X.; Li, M.; Wu, S.; Zhao, Y.; Chen, J.; Gao, D.; Chen, R.; et al. Matrix stiffness-mediated effects on macrophages polarization and their LOXL2 expression. FEBS J. 2020. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, X. Characteristics and Significance of the Pre-metastatic Niche. Cancer Cell 2016, 30, 668–681. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zheng, Q.; Xing, X.; Dong, Y.; Wang, Y.; You, Y.; Chen, R.; Hu, C.; Chen, J.; Gao, D.; et al. Matrix stiffness-upregulated LOXL2 promotes fibronectin production, MMP9 and CXCL12 expression and BMDCs recruitment to assist pre-metastatic niche formation. J. Exp. Clin. Cancer Res. 2018, 37, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, D.; Li, X.; Liu, Z.; Li, T.; Jiang, P.; He, Q.; Tian, F.; Gao, Y.; Wang, D.; et al. 67 laminin receptor promotes the malignant potential of tumour cells up-regulating lysyl oxidase-like 2 expression in cholangiocarcinoma. Dig. Liver Dis. 2014, 46, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Bergeat, D.; Fautrel, A.; Turlin, B.; Merdrignac, A.; Rayar, M.; Boudjema, K.; Coulouarn, C.; Sulpice, L. Impact of stroma LOXL2 overexpression on the prognosis of intrahepatic cholangiocarcinoma. J. Surg. Res. 2016, 203, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Deng, X.; Tian, F.; Li, Z.; Jiang, P.; Zhao, X.; Chen, G.; Chen, Y.; Zheng, P.; Li, D.; et al. The interaction of LOXL2 with GATA6 induces VEGFA expression and angiogenesis in cholangiocarcinoma. Int. J. Oncol. 2019, 55, 657–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Li, D.; Cheng, L.; Wu, H.; Gao, Z.; Liu, Z.; Jiang, W.; Gao, Y.H.; Tian, F.; Zhao, L.; et al. Epithelial-Mesenchymal Transition Induced by Hepatitis C Virus Core Protein in Cholangiocarcinoma. Ann. Surg. Oncol. 2010, 17, 1937–1944. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, Y.; Zhang, X.; Feng, M.; Ma, J.; Li, J.; Yang, X.; Fang, F.; Xia, Q.; Zhang, Z.; et al. Exosome-mediated secretion of LOXL4 promotes hepatocellular carcinoma cell invasion and metastasis. Mol. Cancer 2019, 18, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, J.; Lu, J.; Zhu, W.; Yu, H.; Jing, X.; Wang, Y.; Wang, X.; Wang, X. Derepression of LOXL4 inhibits liver cancer growth by reactivating compromised p53. Cell Death Differ. 2019, 26, 2237–2252. [Google Scholar] [CrossRef]
- Tian, M.; Liu, W.; Jin, L.; Jiang, X.; Yang, L.; Ding, Z.; Shen, Y.; Peng, Y.; Gao, D.; Li, L.; et al. LOXL4 is downregulated in hepatocellular carcinoma with a favorable prognosis. Int. J. Clin. Exp. Pathol. 2015, 8, 3892–3900. [Google Scholar]
- Wong, C.C.-L.; Gilkes, D.M.; Zhang, H.; Chen, J.; Wei, H.; Chaturvedi, P.; Fraley, S.I.; Khoo, U.-S.; Ng, I.O.-L.; Wirtz, D.; et al. Hypoxia-inducible factor 1 is a master regulator of breast cancer metastatic niche formation. Proc. Natl. Acad. Sci. USA 2011, 108, 16369–16374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schietke, R.; Warnecke, C.; Wacker, I.; Schödel, J.; Mole, D.R.; Campean, V.; Amann, K.; Goppelt-Struebe, M.; Behrens, J.; Eckardt, K.-U.; et al. The Lysyl Oxidases LOX and LOXL2 Are Necessary and Sufficient to Repress E-cadherin in Hypoxia. J. Biol. Chem. 2010, 285, 6658–6669. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Gusarova, G.; Wang, Z.; Carr, J.R.; Li, J.; Kim, K.-H.; Qiu, J.; Park, Y.-D.; Williamson, P.R.; Hay, N.; et al. Deregulation of FoxM1b leads to tumour metastasis. EMBO Mol. Med. 2011, 3, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, I.; Kikkawa, N.; Matsushita, R.; Kato, M.; Kurozumi, A.; Nishikawa, R.; Goto, Y.; Koshizuka, K.; Hanazawa, T.; Enokida, H.; et al. Tumor-suppressive microRNAs (miR-26a/b, miR-29a/b/c and miR-218) concertedly suppressed metastasis-promoting LOXL2 in head and neck squamous cell carcinoma. J. Hum. Genet. 2016, 61, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Kurozumi, A.; Goto, Y.; Matsushita, R.; Okato, A.; Nishikawa, R.; Fukumoto, I.; Koshizuka, K.; Ichikawa, T.; Seki, N. Regulation of metastasis-promoting LOXL2 gene expression by antitumor microRNAs in prostate cancer. J. Hum. Genet. 2016, 62, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurozumi, A.; Kato, M.; Goto, Y.; Matsushita, R.; Nishikawa, R.; Okato, A.; Fukumoto, I.; Ichikawa, T.; Seki, N. Regulation of the collagen cross-linking enzymes LOXL2 and PLOD2 by tumor-suppressive microRNA-26a/b in renal cell carcinoma. Int. J. Oncol. 2016, 48, 1837–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saatci, O.; Kaymak, A.; Raza, U.; Ersan, P.G.; Akbulut, O.; Banister, C.E.; Sikirzhytski, V.; Tokat, U.M.; Aykut, G.; Ansari, S.A.; et al. Targeting lysyl oxidase (LOX) overcomes chemotherapy resistance in triple negative breast cancer. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Boufraqech, M.; Nilubol, N.; Zhang, L.; Gara, S.K.; Sadowski, S.M.; Mehta, A.; He, M.; Davis, S.; Dreiling, J.; Copland, J.A.; et al. miR30a Inhibits LOX Expression and Anaplastic Thyroid Cancer Progression. Cancer Res. 2015, 75, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, K.; Seki, N.; Mataki, H.; Matsushita, R.; Kamikawaji, K.; Kumamoto, T.; Takagi, K.; Goto, Y.; Nishikawa, R.; Kato, M.; et al. Tumor-suppressive microRNA-29 family inhibits cancer cell migration and invasion directly targeting LOXL2 in lung squamous cell carcinoma. Int. J. Oncol. 2016, 48, 450–460. [Google Scholar] [CrossRef]
- Kamikawaji, K.; Seki, N.; Watanabe, M.; Mataki, H.; Kumamoto, T.; Takagi, K.; Mizuno, K.; Inoue, H. Regulation of LOXL2 and SERPINH1 by antitumor microRNA-29a in lung cancer with idiopathic pulmonary fibrosis. J. Hum. Genet. 2016, 61, 985–993. [Google Scholar] [CrossRef]
- Duan, Z.; Li, L.; Li, Y. Involvement of miR-30b in kynurenine-mediated lysyl oxidase expression. J. Physiol. Biochem. 2019, 75, 135–142. [Google Scholar] [CrossRef]
- Ren, J.; Wu, X.; He, W.; Shao, J.; Cheng, B.; Huang, T. Lysyl Oxidase 473 G>A Polymorphism and Breast Cancer Susceptibility in Chinese Han Population. DNA Cell Biol. 2011, 30, 111–116. [Google Scholar] [CrossRef]
- Wang, G.; Shen, Y.; Cheng, G.; Bo, H.; Lin, J.; Zheng, M.; Li, J.; Zhao, Y.; Li, W. Lysyl Oxidase Gene G473A Polymorphism and Cigarette Smoking in Association with a High Risk of Lung and Colorectal Cancers in a North Chinese Population. Int. J. Environ. Res. Public Heal. 2016, 13, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csiszar, K.; Fong, S.F.T.; Ujfalusi, A.; Krawetz, S.A.; Salvati, E.P.; MacKenzie, J.W.; Boyd, C.D. Somatic mutations of the lysyl oxidase gene on chromosome 5q23.1 in colorectal tumors. Int. J. Cancer 2002, 97, 636–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Cong, J.-L.; Qu, L.-Y.; Jiang, L.; Wang, Y. Association between Lysyl Oxidase G473A Polymorphism and Ovarian Cancer in the Han Chinese Population. J. Int. Med. Res. 2012, 40, 917–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Cueva, A.; Emmerling, M.; Lim, S.L.; Yang, S.; Trackman, P.C.; Sonenshein, G.E.; Kirsch, K.H. A polymorphism in the lysyl oxidase propeptide domain accelerates carcinogen-induced cancer. Carcinogenesis 2018, 39, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Hecht, J.R.; Benson, A.B.; Vyushkov, D.; Yang, Y.; Bendell, J.; Verma, U. A Phase II, Randomized, Double-Blind, Placebo-Controlled Study of Simtuzumab in Combination with FOLFIRI for the Second-Line Treatment of Metastatic KRAS Mutant Colorectal Adenocarcinoma. Oncologist 2017, 22, 243. [Google Scholar] [CrossRef] [Green Version]
- Benson, A.B.; Wainberg, Z.A.; Hecht, J.R.; Vyushkov, D.; Dong, H.; Bendell, J.; Kudrik, F. A Phase II Randomized, Double-Blind, Placebo-Controlled Study of Simtuzumab or Placebo in Combination with Gemcitabine for the First-Line Treatment of Pancreatic Adenocarcinoma. Oncologist 2017, 22, 241. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-E.; Kim, Y. A Tissue-specific Variant of the Human Lysyl Oxidase-like Protein 3 (LOXL3) Functions as an Amine Oxidase with Substrate Specificity. J. Biol. Chem. 2006, 281, 37282–37290. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kim, S.-S.; Jung, S.T.; Park, J.-Y.; Yoo, H.-W.; Ko, J.; Csiszar, K.; Choi, S.-Y.; Kim, Y. Expression and Purification of Enzymatically Active Forms of the Human Lysyl Oxidase-like Protein 4. J. Biol. Chem. 2003, 278, 52071–52074. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.T.; Kim, M.S.; Seo, J.Y.; Kim, H.C.; Kim, Y. Purification of enzymatically active human lysyl oxidase and lysyl oxidase-like protein from Escherichia coli inclusion bodies. Protein Expr. Purif. 2003, 31, 240–246. [Google Scholar] [CrossRef]
- Tang, S.S.; Trackman, P.C.; Kagan, H.M. Reaction of aortic lysyl oxidase with beta-aminopropionitrile. J. Biol. Chem. 1983, 258, 4331–4338. [Google Scholar]
- Trackman, P.C.; Kagan, H.M. Nonpeptidyl amine inhibitors are substrates of lysyl oxidase. J. Biol. Chem. 1979, 254, 7831–7836. [Google Scholar] [PubMed]
- Bondareva, A.; Downey, C.M.; Ayres, F.; Liu, W.; Boyd, S.K.; Hallgrímsson, B.; Jirik, F.R. The Lysyl Oxidase Inhibitor, β-Aminopropionitrile, Diminishes the Metastatic Colonization Potential of Circulating Breast Cancer Cells. PLoS ONE 2009, 4, e5620. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, S.; Li, W.; Chen, J.; Xiao, X.; Wang, Y.; Yan, G.; Chen, L. Inactivation of lysyl oxidase by β-aminopropionitrile inhibits hypoxia-induced invasion and migration of cervical cancer cells. Oncol. Rep. 2012, 29, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Zhang, N.; Liu, H.; Zhao, L.; Liu, J.; Wan, J.; Wu, W.; Lei, H.; Liu, R.; Han, M. Lysyl oxidase inhibition via β-aminoproprionitrile hampers human umbilical vein endothelial cell angiogenesis and migration in vitro. Mol. Med. Rep. 2018, 17, 5029–5036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, M.; Adamo, H.; Bergh, A.; Bergström, S.H. Inhibition of Lysyl Oxidase and Lysyl Oxidase-Like Enzymes Has Tumour-Promoting and Tumour-Suppressing Roles in Experimental Prostate Cancer. Sci. Rep. 2016, 6, 19608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Zhu, C.-C.; Ni, B.; Zhang, Z.-Z.; Jiang, S.-H.; Hu, L.-P.; Wang, X.; Zhang, X.-X.; Huang, P.-Q.; Yang, Q.; et al. Lysyl oxidase promotes liver metastasis of gastric cancer via facilitating the reciprocal interactions between tumor cells and cancer associated fibroblasts. EBioMedicine 2019, 49, 157–171. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.B.; Ikenaga, N.; Peng, Z.; Sverdlov, D.Y.; Greenstein, A.; Smith, V.; Schuppan, D.; Popov, Y.V. Lysyl oxidase activity contributes to collagen stabilization during liver fibrosis progression and limits spontaneous fibrosis reversal in mice. FASEB J. 2015, 30, 1599–1609. [Google Scholar] [CrossRef]
- Wang, X.; Huang, W.; Liu, G.; Cai, W.; Millard, R.W.; Wang, Y.; Chang, J.; Peng, T.; Fan, G.-C. Cardiomyocytes mediate anti-angiogenesis in type 2 diabetic rats through the exosomal transfer of miR-320 into endothelial cells. J. Mol. Cell. Cardiol. 2014, 74, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, K.; Liu, Y.; Xu, Y.; Zhang, F.; Yang, H.; Liu, J.; Pan, T.; Chen, J.; Wu, M.; et al. Exosomes mediate the cell-to-cell transmission of IFN-α-induced antiviral activity. Nat. Immunol. 2013, 14, 793–803. [Google Scholar] [CrossRef]
- Kulshreshtha, A.; Ahmad, T.; Agrawal, A.; Ghosh, B. Proinflammatory role of epithelial cell–derived exosomes in allergic airway inflammation. J. Allergy Clin. Immunol. 2013, 131, 1194–1203.e14. [Google Scholar] [CrossRef]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of MicroRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, H.M.; Vaysberg, M.; Mikels, A.; McCauley, S.; Velayo, A.C.; Garcia, C.; Smith, V. Modulation of Lysyl Oxidase-like 2 Enzymatic Activity by an Allosteric Antibody Inhibitor. J. Biol. Chem. 2010, 285, 20964–20974. [Google Scholar] [CrossRef] [Green Version]
- Barry-Hamilton, V.; Spangler, R.; Marshall, D.; McCauley, S.A.; Rodriguez, H.M.; Oyasu, M.; Mikels, A.; Vaysberg, M.; Ghermazien, H.; Wai, C.; et al. Allosteric inhibition of lysyl oxidase-like-2 impedes the development of a pathologic microenvironment. Nat. Med. 2010, 16, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Muir, A.J.; Levy, C.; Janssen, H.L.; Montano-Loza, A.J.; Shiffman, M.L.; Caldwell, S.; Luketic, V.; Ding, D.; Jia, C.; McColgan, B.J.; et al. Simtuzumab for Primary Sclerosing Cholangitis: Phase 2 Study Results With Insights on the Natural History of the Disease. Hepatology 2019, 69, 684–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilter, H.C.; Findlay, A.D.; Perryman, L.; Yow, T.T.; Moses, J.; Zahoor, A.; Turner, C.I.; Deodhar, M.; Foot, J.S.; Zhou, W.; et al. The lysyl oxidase like 2/3 enzymatic inhibitor, PXS-5153A, reduces crosslinks and ameliorates fibrosis. J. Cell. Mol. Med. 2018, 23, 1759–1770. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, J.H.; Rowbottom, M.W.; Lonergan, D.; Darlington, J.; Prodanovich, P.; King, C.D.; Evans, J.F.; Bain, G. Small Molecule Lysyl Oxidase-like 2 (LOXL2) Inhibitors: The Identification of an Inhibitor Selective for LOXL2 over LOX. ACS Med. Chem. Lett. 2017, 8, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Leung, L.M.H.; Niculescu-Duvaz, D.; Smithen, D.; Lopes, F.; Callens, C.; McLeary, R.; Saturno, G.; Davies, L.; Aljarah, M.; Brown, M.; et al. Anti-metastatic Inhibitors of Lysyl Oxidase (LOX): Design and Structure-Activity Relationships. J. Med. Chem. 2019, 62, 5863–5884. [Google Scholar] [CrossRef]
- Smithen, D.A.; Leung, L.M.H.; Challinor, M.; Lawrence, R.; Tang, H.; Niculescu-Duvaz, D.; Pearce, S.P.; McLeary, R.; Lopes, F.; Aljarah, M.; et al. 2-Aminomethylene-5-sulfonylthiazole Inhibitors of Lysyl Oxidase (LOX) and LOXL2 Show Significant Efficacy in Delaying Tumor Growth. J. Med. Chem. 2019, 63, 2308–2324. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, X.; Huang, Y.; Ma, Y.; Jin, X.; Wang, H.; Wang, J. Inhibition of lysyl oxidase expression by dextran sulfate affects invasion and migration of gastric cancer cells. Int. J. Mol. Med. 2018, 42, 2737–2749. [Google Scholar] [CrossRef]
- Chen, X.; Kou, Y.; Lu, Y.; Pu, Y. Salidroside ameliorated hypoxia-induced tumorigenesis of BxPC-3 cells via downregulating hypoxia-inducible factor (HIF)-1α and LOXL2. J. Cell. Biochem. 2019, 121, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, X.; Zhao, P.; Tong, B.; Wei, Z.; Dai, Y. Escin Ia suppresses the metastasis of triple-negative breast cancer by inhibiting epithelial-mesenchymal transition via down-regulating LOXL2 expression. Oncotarget 2016, 7, 23684–23699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morisawa, A.; Okui, T.; Shimo, T.; Ibaragi, S.; Okusha, Y.; Ono, M.; Nguyen, T.T.H.; Hassan, N.M.M.; Sasaki, A. Ammonium tetrathiomolybdate enhances the antitumor effects of cetuximab via the suppression of osteoclastogenesis in head and neck squamous carcinoma. Int. J. Oncol. 2018, 52, 989–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, W.; Liu, J.; Gao, Y.; Zhong, G.; Ding, B.; Xu, L.; Fan, W. MicroRNA regulation of liver cancer stem cells. Am. J. Cancer Res. 2018, 8, 1126–1141. [Google Scholar] [PubMed]
- Ye, M.-F.; Zhang, J.-G.; Guo, T.-X.; Pan, X.-J. MiR-504 inhibits cell proliferation and invasion by targeting LOXL2 in non small cell lung cancer. Biomed. Pharmacother. 2018, 97, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, W.; Yang, J.; Huang, J.; Kang, G.; Hu, H.; Xie, S. Downregulation of lysyl oxidase-like 4 LOXL4 by miR-135a-5p promotes lung cancer progression in vitro and in vivo. J. Cell. Physiol. 2019, 234, 18679–18687. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Liu, G.; Huang, J.; Hu, H.; Jiang, W. miR-210 promotes lung adenocarcinoma proliferation, migration, and invasion by targeting lysyl oxidase-like 4. J. Cell. Physiol. 2019, 234, 14050–14057. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LOX Family Member | Patients | Expression Level in Tumor | Clinical Relevance | PMID |
|---|---|---|---|---|
| LOX | HCC | Up | Prognostic marker for high recurrence rate and poor OS | 30919528 |
| HCC | Up | Prognostic marker for poor OS and DFS | 26048020 | |
| LOXL2 | HCC | Up | Prognostic marker for poor OS | 28449718 |
| HCC | Up | Prognostic marker for poor OS and DFS | 29620290 | |
| HCC | Up | Prognostic marker for poor OS and RFS | 29938458 | |
| HCC | Up | Prognostic marker for poor OS and DFS | 30506621 | |
| CCA | Up | Prognostic marker for poor OS and DFS | 31322171 | |
| CCA | Up | Prognostic marker for poor OS and DFS | 27363654 | |
| HCC | Up | Serum LOXL2 as an excellent differential marker | 25048396 | |
| LOXL4 | HCC | Up | Prognostic marker for poor OS | 33068461 |
| HCC | Up | High LOXL4 indicates poor OS and DFS | 30704479 | |
| HCC | Down | High LOXL4 indicates high recurrence rate and poor OS | 26097573 |
| Liver Cancer Context | Donor or Approach | Recipient | Signaling Pathway | Biological Activity | PMID |
|---|---|---|---|---|---|
| HCC cell line (SK-hep-1, MHCC-97-H, Hep-G2, SSMC-7721, Huh7) | -- | -- | LOX→MAPK→VEGF | ↓ proliferation; ↓migration; ↓ invasion | 26048020 |
| HCC cell line (SK-hep-1, Hep-G2) | Ad-LOX-PP | HCC | LOX-PP→p-ERK LOX-PP┤MMP-2, MMP9 | ↑ apoptosis; ↓ proliferation ↓ migration; ↓ invasion | 24573150 |
| Xenograft using Sphere-derived TIC (HuH-7) | TIC | EC | LOX→ VEGF | ↑ angiogenesis | 30720077 |
| HCC cell line (SK-hep-1) | -- | -- | HIF1→LOX→ TWIST/Vimentin/Slug | ↑ EMT ↑ cell migration and invasion | 30919528 |
| Liver Cancer Context | Donor or Stimulant | Recipient | Signaling Pathway | Biological Activity | PMID |
|---|---|---|---|---|---|
| HCC cell lines (Huh7 and Hep3B) | HCC | EC | LOXL2→ Snail ┤FBP1┤HIF-1α→VEGF | ↑ Proliferation; ↑ glycolysis; ↑ angiogenesis | 32323822 |
| Mφ (THP-1 cell ine) | ↑ Matrix stiffness | Mφ | integrin β5 →FAK→MEK1/2-ERK1/2→ HIF-1α→LOXL2 | ↑ M2 phenotype of Mφ | 32964626 |
| HBx-transfected HCC (HepG2, Hep3B, MHCC97L) | HCC | collagen | HBx→HIF-1α→LOXL2 | ↑ ECM collagen cross-link ↑ Cell invasion ↑ Tumor growth ↑ Metastasis | 29799025 |
| HCC cell line (HepG2) | -- | -- | HIF-1α→LOXL2→E-cadherin/Vimentin | ↑ EMT ↑ cell migration and invasion ↑ Vasculogenic mimicry | 28449718 |
| HCC cell lines (MHCC97L) | HCC | Collagen; BMDC | HIF-1α→LOXL2 TGF-β→SMAD4→LOXL2 miR-26a ┤LOXL2 miR-29a ┤LOXL2 | ↑ tissue stiffness ↑ local metastasis ↑ BMDC requitement to pre-metastatic site ↑ distant metastasis | 25048396 |
| HCC cell lines (MHCC97L) | HCC | Lung fibroblast; BMDC | ↑ Matrix stiffness→ integrin→JNK→c-Jun→LOXL2 (HCC) LOXL2→AKT→MMP9, CXCL12 (lung fibroblast) | ↑ HCC adhere to lung fibrobalst ↑ BMDC migration ↑ HCC migration | 29728125 |
| CCA cell lines (QBC939 and RBE) | CCA | EC | LOXL2-GATA6 interaction→ VEGFA | ↑ Tumor growth ↑ angiogenesis | 31322171 |
| QBC939 | CCA | -- | 67LR→LOXL2 | ↑ cell invasion ↑ tumor metastasis | 24794791 |
| Liver Cancer Context | Donor | Recipient | Signaling Pathway | Biological Activity | PMID |
|---|---|---|---|---|---|
| Mouse treated with CDAA diet and CCl4 | HCC | Mφ | LOXL4 (HCC)→H2O2→IFNAR1→STAT1/3→PD-L1 (Mφ) ┤ cytotoxicity (T cell) | ↑ Immunosuppressive phenotype of Mφ ↓ T cell cytotoxicity; ↑ Tumor growth | 33068461 |
| HCC cell line xenograft | HCC | HCC | TFG-β→exosomal LOXL4→FAK→Src | ↑ cell migration and invasion ↑ intrahepatic and pulmonary metastasis | 30704479 |
| HCC | EC | Exosomal LOXL4→ HUVEC migration and tube formation | ↑ Angiogenesis | 30704479 |
| Agents | Biological Property | Targets of Action | Disease Model | PMID |
|---|---|---|---|---|
| BAPN | Small-molecule inhibitor | (-) LOX, LOXL1-4 | HCC | 30720077 |
| (-) LOXL2 | HCC | 29620290 | ||
| (-) LOX | Liver metastasis of GC | 31678002 | ||
| (-) LOX, LOXL1-4 | Liver fibrosis | 26700732 | ||
| GW4869 | N-SMase inhibitor | Exosome-mediated transfer of LOXL4 | HCC | 30704479 |
| pterostilbene/curcumin analogues | Stilbene/curcuminoids compounds | (-) LOX | HCC | 23560895 |
| AB0023 | mAb | (-) LOXL2 | Liver fibrosis | 28073888 |
| (-) LOXL2 | Liver fibrosis | 20818376 | ||
| LOXL2-IN-1 hydrochloride | Small-molecule inhibitor | (-) LOXL2 | HCC | 32323822 |
| PXS-5153A | Small-molecule inhibitor | (-) LOXL2/3 | Liver fibrosis | 30536539 |
| 5-aza-CR | DNA methylation Inhibitor | (+) LOXL4 | HCC | 30728460 |
| CCT365623 | Small-molecule inhibitor | (-) LOX | Lung metastasis of BC | 31070916 |
| AMTz-21b | Small-molecule inhibitor | (-) LOX, LOXL2 | Lung metastasis of BC | 31430136 |
| Salidroside | Glucoside of tyrosol | (-) LOX, LOXL1-4 | Lung metastasis of PC | 31162697 |
| escin Ia | Subclass of SFAC | (-) LOXL2 | Lung metastasis of BC | 27008697 |
| ammonium tetrathiomolybdate | Copper chelator | (-) LOX | Bone invasion of HNSCC | 29328370 |
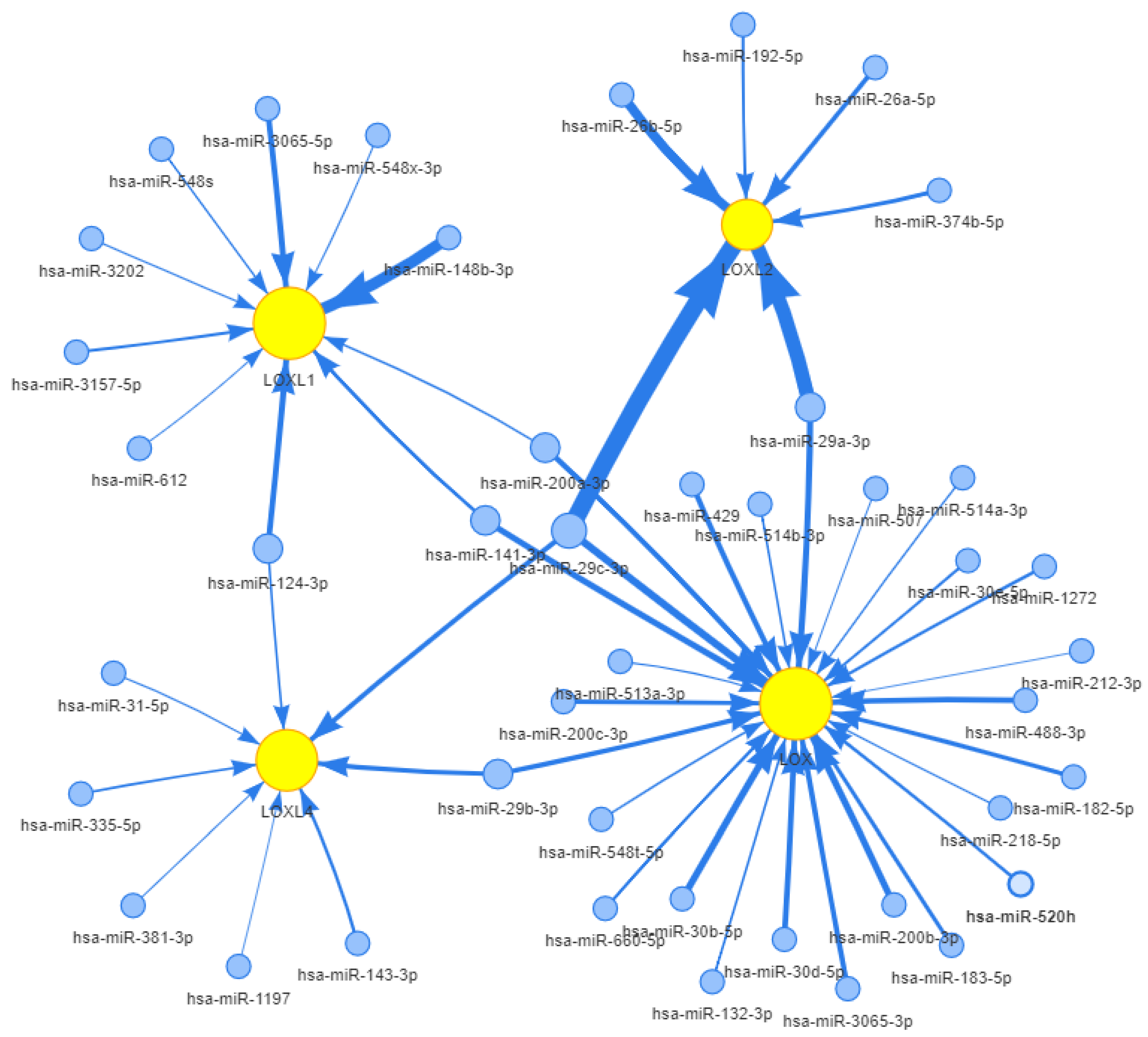
| miR-26a, miR-29a | Non-coding RNAs | (-) LOXL2 | HCC | 25048396 |
| miR-26a/b, miR-29a/b/c, miR-218 | Non-coding RNAs | (-) LOXL2 | HNSCC | 26490187 |
| miR-26a/b, miR-29a/b/c, miR-218 | Non-coding RNAs | (-) LOXL2 | PC | 27278788 |
| miR-26a/b | Non-coding RNAs | (-) LOXL2 | RCC | 26983694 |
| miR-142 | Non-coding RNAs | (-) LOX | BC | 32415208 |
| miR-30a | Non-coding RNAs | (-) LOX | ATC | 25488748 |
| miR-29a/b/c | Non-coding RNAs | (-) LOXL2 | LSCC | 26676674 |
| miR-29a | Non-coding RNAs | (-) LOXL2 | NSCLC | 27488440 |
| miR-30b | Non-coding RNAs | (-) LOX | GCCL | 31093946 |
| miR-135a | Non-coding RNAs | (-) LOXL4 | NSCLC | 30993701 |
| miR-504 | Non-coding RNAs | (-) LOXL2 | NSCLC | 29156517 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, H.-Y.; Li, C.-J.; Yang, Y.-L.; Huang, Y.-H.; Hsiau, Y.-T.; Chu, P.-Y. Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer. Int. J. Mol. Sci. 2020, 21, 9751. https://doi.org/10.3390/ijms21249751
Lin H-Y, Li C-J, Yang Y-L, Huang Y-H, Hsiau Y-T, Chu P-Y. Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer. International Journal of Molecular Sciences. 2020; 21(24):9751. https://doi.org/10.3390/ijms21249751
Chicago/Turabian StyleLin, Hung-Yu, Chia-Jung Li, Ya-Ling Yang, Ying-Hsien Huang, Ya-Tze Hsiau, and Pei-Yi Chu. 2020. "Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer" International Journal of Molecular Sciences 21, no. 24: 9751. https://doi.org/10.3390/ijms21249751
APA StyleLin, H. -Y., Li, C. -J., Yang, Y. -L., Huang, Y. -H., Hsiau, Y. -T., & Chu, P. -Y. (2020). Roles of Lysyl Oxidase Family Members in the Tumor Microenvironment and Progression of Liver Cancer. International Journal of Molecular Sciences, 21(24), 9751. https://doi.org/10.3390/ijms21249751