Differences in the Inflammatory Response of White Adipose Tissue and Adipose-Derived Stem Cells


 ,
, 
Abstract
:1. Introduction
2. Results
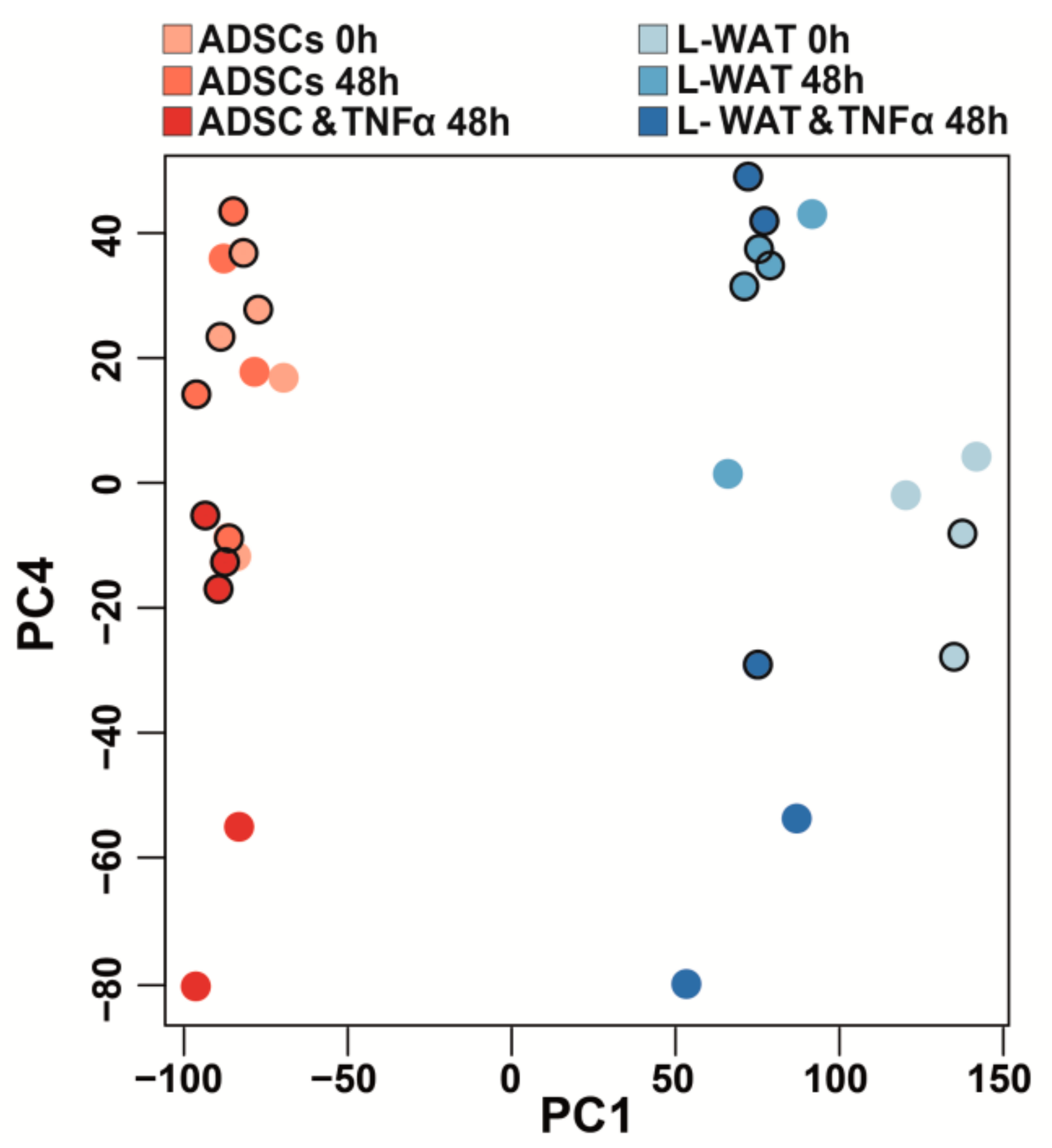
2.1. Gene Expression after TNFα Treatment of ADSCs Is More Homogenous in Comparison to L-WAT
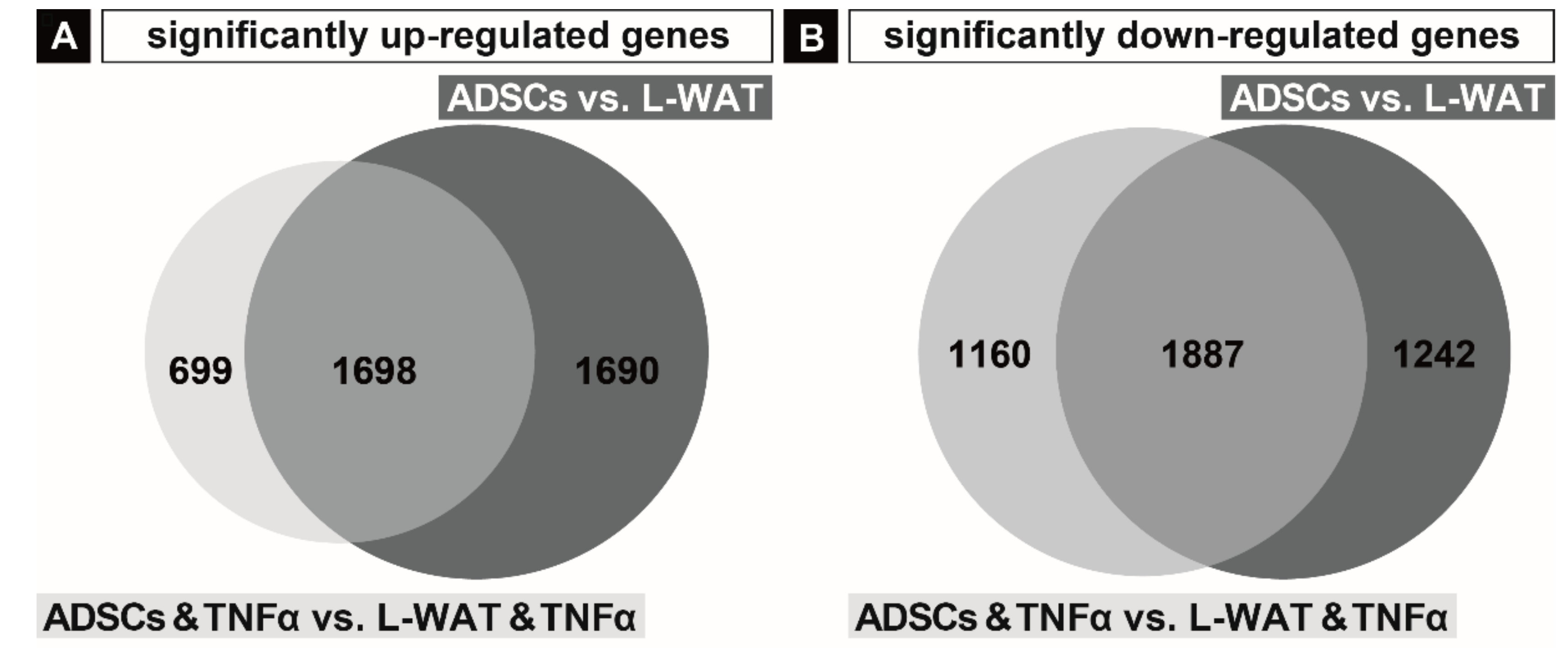
2.2. Gene Expression Regulation in Fat and ADSCs Treated with TNFα
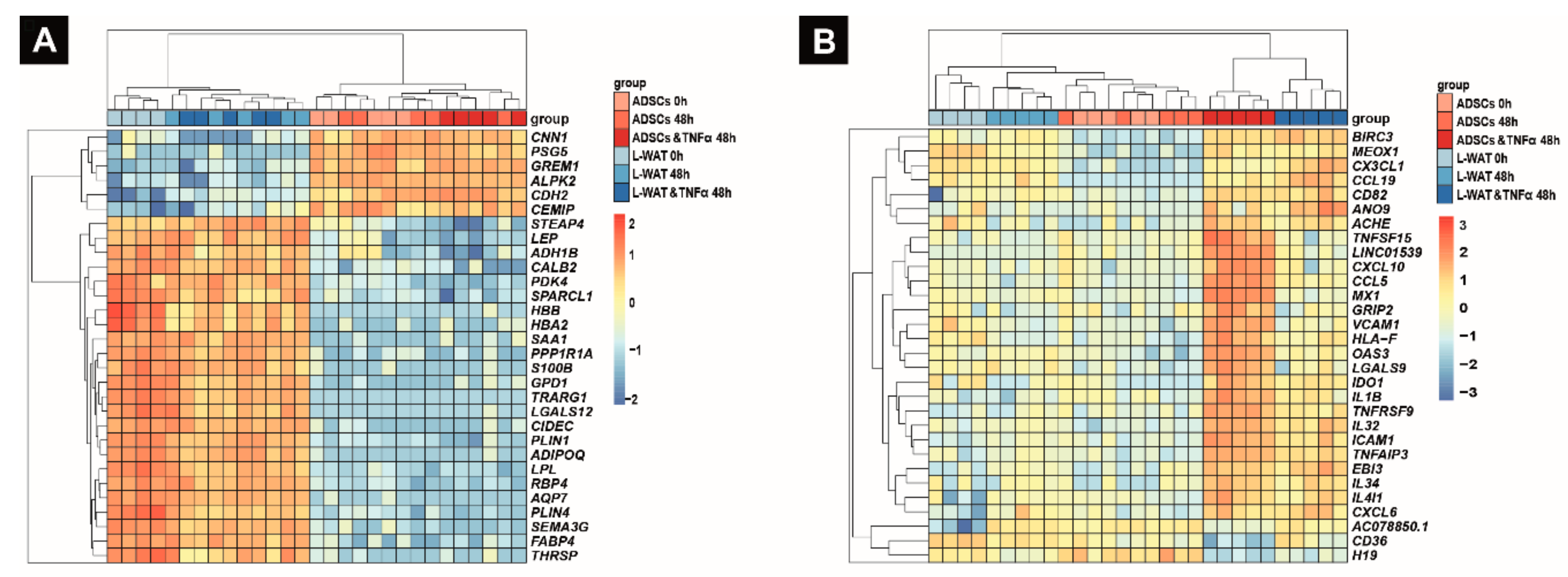
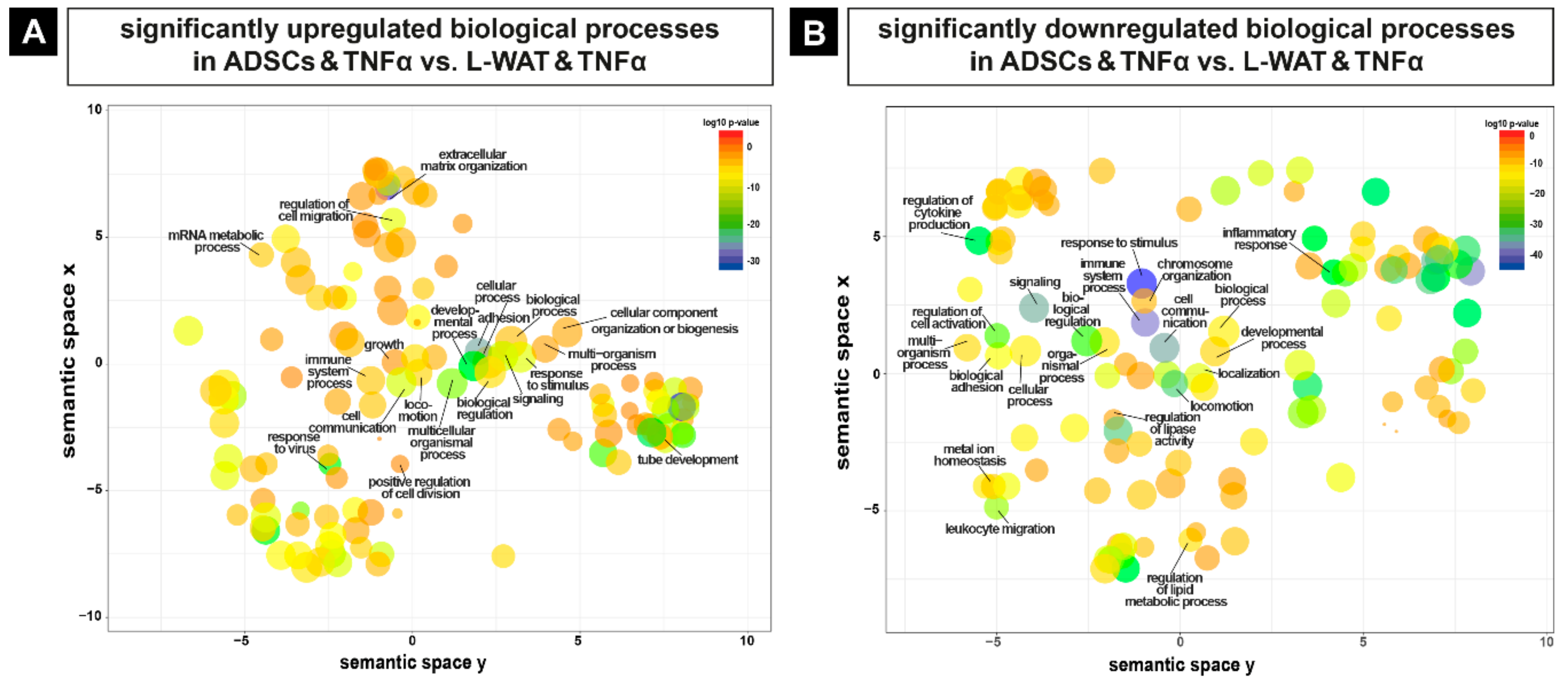
2.3. Biological Pathways that Are Regulated Upon TNFα Exposure in L-WAT and ADSCs
3. Discussion
4. Materials and Methods
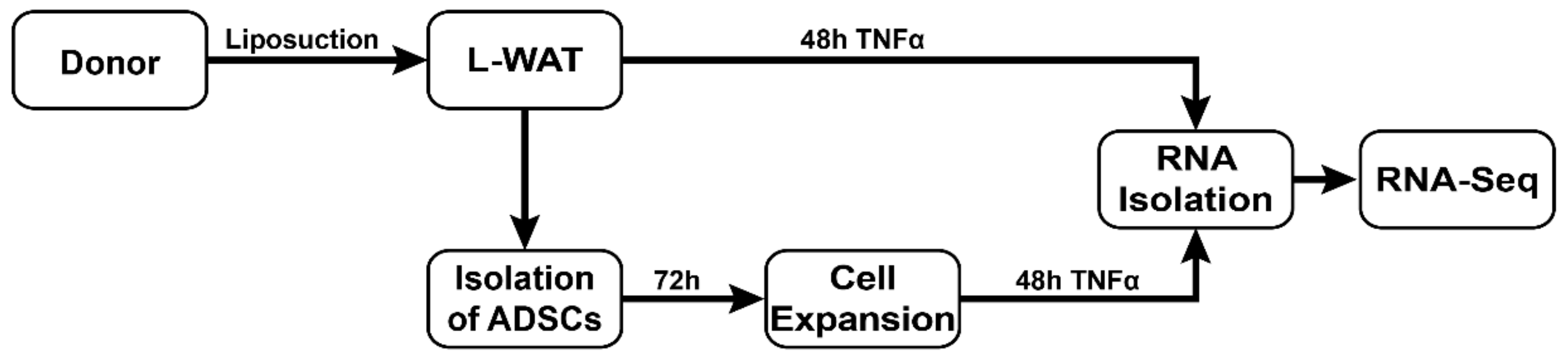
4.1. Ethics Statement and Sample Acquisition
4.2. Preparation of White Adipose Tissue, Cell Isolation and Culture Conditions
4.3. Deep RNA-Sequencing and Bioinformatics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADSCs | adipose-derived stem cells |
| GO | gene ontology |
| GSEA | gene set enrichment analysis |
| HBV | hepatitis B virus |
| HCV | hepatitis C virus |
| HIV | human immunodeficiency virus |
| IFN | Interferon |
| IL | Interleukin |
| L-WAT | liposuctioned white adipose tissue |
| PCA | principal component analysis |
| SVF | stromal vascular fraction |
| TNFα | tumor necrosis factor alpha |
| WAT | white adipose tissue |
References
- Cavaillon, J.M. Pro-versus anti-inflammatory cytokines: Myth or reality. Cell. Mol. Biol. 2001, 47, 695–702. [Google Scholar] [PubMed]
- Van Meegeren, M.E.R.; Roosendaal, G.; Jansen, N.W.D.; Wenting, M.J.G.; Van Wesel, A.C.W.; Van Roon, J.A.G.; Lafeber, F.P.J.G. IL-4 alone and in combination with IL-10 protects against blood-induced cartilage damage. Osteoarthr. Cartil. 2012, 20, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relic, B.; Guicheux, J.; Mezin, F.; Lubberts, E.; Togninalli, D.; Garcia, I.; van den Berg, W.B.; Guerne, P.-A. Il-4 and IL-13, but not IL-10, protect human synoviocytes from apoptosis. J. Immunol. 2001, 166, 2775–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuerwegh, A.J.; Dombrecht, E.J.; Stevens, W.J.; Van Offel, J.F.; Bridts, C.H.; De Clerck, L.S. Influence of pro-inflammatory (IL-1α, IL-6, TNF-α, IFN-γ) and anti-inflammatory (IL-4) cytokines on chondrocyte function. Osteoarthr. Cartil. 2003, 11, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Dieppe, P. Inflammation in osteoarthritis. Rheumatology 1978, 17, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Waldner, M.; Zhang, W.; James, I.B.; Allbright, K.; Havis, E.; Bliley, J.M.; Almadori, A.; Schweizer, R.; Plock, J.A.; Washington, K.M.; et al. Characteristics and immunomodulating functions of adipose-derived and bone marrow-derived mesenchymal stem cells across defined human leukocyte antigen barriers. Front. Immunol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mun, C.H.; Kang, M.I.; Shin, Y.D.; Kim, Y.; Park, Y.B. The Expression of Immunomodulation-Related Cytokines and Genes of Adipose- and Bone Marrow-Derived Human Mesenchymal Stromal Cells from Early to Late Passages. Tissue Eng. Regen. Med. 2018, 15, 771–779. [Google Scholar] [CrossRef]
- Yoshizumi, Y.; Yukawa, H.; Iwaki, R.; Fujinaka, S.; Kanou, A.; Kanou, Y.; Yamada, T.; Nakagawa, S.; Ohara, T.; Nakagiri, K.; et al. Immunomodulatory Effects of Adipose Tissue-Derived Stem Cells on Concanavalin A-Induced Acute Liver Injury in Mice. Cell Med. 2016, 9, 21–33. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Smorlesi, A.; Frontini, A.; Giordano, A.; Cinti, S. The adipose organ: White-brown adipocyte plasticity and metabolic inflammation. Obes. Rev. 2012, 13, 83–96. [Google Scholar] [CrossRef]
- Meyer, J.; Engelmann, R.; Kamp, G.; Peters, K. Human adipocytes and CD34 + cells from the stromal vascular fraction of the same adipose tissue differ in their energy metabolic enzyme configuration. Exp. Cell Res. 2019, 380, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Pak, J.; Lee, J.H.; Kartolo, W.A.; Lee, S.H. Cartilage Regeneration in Human with Adipose Tissue-Derived Stem Cells: Current Status in Clinical Implications. Biomed Res. Int. 2016, 2016, 4702674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiekman, M.; van Dongen, J.A.; Willemsen, J.C.; Hoppe, D.L.; van der Lei, B.; Harmsen, M.C. The power of fat and its adipose-derived stromal cells: Emerging concepts for fibrotic scar treatment. J. Tissue Eng. Regen. Med. 2017, 11, 3220–3235. [Google Scholar] [CrossRef]
- Pak, J.; Lee, J.H.; Park, K.S.; Jeong, B.C.; Lee, S.H. Regeneration of Cartilage in Human Knee Osteoarthritis with Autologous Adipose Tissue-Derived Stem Cells and Autologous Extracellular Matrix. Biores. Open Access 2016, 5, 192–200. [Google Scholar] [CrossRef]
- Ter Huurne, M.; Schelbergen, R.; Blattes, R.; Blom, A.; De Munter, W.; Grevers, L.C.; Jeanson, J.; Noël, D.; Casteilla, L.; Jorgensen, C.; et al. Antiinflammatory and chondroprotective effects of intraarticular injection of adipose-derived stem cells in experimental osteoarthritis. Arthritis Rheum. 2012, 64, 3604–3613. [Google Scholar] [CrossRef]
- Heydarkhan-Hagvall, S.; Schenke-Layland, K.; Yang, J.Q.; Heydarkhan, S.; Xu, Y.; Zuk, P.A.; MacLellan, W.R.; Beygui, R.E. Human adipose stem cells: A potential cell source for cardiovascular tissue engineering. Cells Tissues Organs 2008, 187, 263–274. [Google Scholar] [CrossRef]
- Moustaki, M.; Papadopoulos, O.; Verikokos, C.; Karypidis, D.; Masud, D.; Kostakis, A.; Papastefanaki, F.; Roubelakis, M.; Perrea, D. Application of adipose-derived stromal cells in fat grafting: Basic science and literature review (Review). Exp. Ther. Med. 2017, 2415–2423. [Google Scholar] [CrossRef] [Green Version]
- Stojanović, S.; Najman, S. The Effect of Conditioned Media of Stem Cells Derived from Lipoma and Adipose Tissue on Macrophages’ Response and Wound Healing in Indirect Co-culture System In Vitro. Int. J. Mol. Sci. 2019, 20, 1671. [Google Scholar] [CrossRef] [Green Version]
- Kawata, Y.; Tsuchiya, A.; Seino, S.; Watanabe, Y.; Kojima, Y.; Ikarashi, S.; Tominaga, K.; Yokoyama, J.; Yamagiwa, S.; Terai, S. Early injection of human adipose tissue-derived mesenchymal stem cell after inflammation ameliorates dextran sulfate sodium-induced colitis in mice through the induction of M2 macrophages and regulatory T cells. Cell Tissue Res. 2019, 376, 257–271. [Google Scholar] [CrossRef]
- Leto Barone, A.A.; Khalifian, S.; Lee, W.P.A.; Brandacher, G. Immunomodulatory Effects of Adipose-Derived Stem Cells: Fact or Fiction? Biomed. Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Puissant, B.; Barreau, C.; Bourin, P.; Clavel, C.; Corre, J.; Bousquet, C.; Taureau, C.; Cousin, B.; Abbal, M.; Laharrague, P.; et al. Immunomodulatory effect of human adipose tissue-derived adult stem cells: Comparison with bone marrow mesenchymal stem cells. Br. J. Haematol. 2005, 129, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Huaman, O.; Bahamonde, J.; Cahuascanco, B.; Jervis, M.; Palomino, J.; Torres, C.G.; Peralta, O.A. Immunomodulatory and immunogenic properties of mesenchymal stem cells derived from bovine fetal bone marrow and adipose tissue. Res. Vet. Sci. 2019, 124, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Guo, B.; Xie, J.; Deng, S.; Fu, N.; Lin, S.; Li, G.; Lin, Y.; Cai, X. Crosstalk between adipose-derived stem cells and chondrocytes: When growth factors matter. Bone Res. 2016, 4, 15036. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Kabata, T.; Hayashi, K.; Maeda, T.; Kajino, Y.; Iwai, S.; Fujita, K.; Hasegawa, K.; Inoue, D.; Sugimoto, N.; et al. The paracrine effect of adipose-derived stem cells inhibits osteoarthritis progression. BMC Musculoskelet. Disord. 2015, 16, 236. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Thacker, R.I.; Hall, B.E.; Kong, R.; Granneman, J.G. Exploring the activated adipogenic niche. Cell Cycle 2014, 13, 184–190. [Google Scholar] [CrossRef] [Green Version]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Santalla, M.; Manche, P.; Menta, R.; López-Belmonte, J.; Bueren, J.; Dalemans, W. Human Adipose-Derived Mesenchymal Stem Cells Modulate Experimental Autoimmune Arthritis by Modifying Early Adaptive T Cell Responses. Stem Cells 2015, 33, 3493–3503. [Google Scholar]
- Baharlou, R.; Rashidi, N.; Ahmadi-Vasmehjani, A.; Khoubyari, M.; Sheikh, M.; Erfanian, S. Immunomodulatory Effects of Human Adipose Tissue-derived Mesenchymal Stem Cells on T Cell Subsets in Patients with Rheumatoid Arthritis. Iran. J. Allergy, Asthma Immunol. 2019, 18, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Chien, C.M.; Chen, Y.W.; Chen, C.C.; Wu, Y.C.; Huang, S.H.; Lee, S.S.; Lai, C.S.; Lin, S.D.; Wang, C.J.; Kuo, Y.R. Adipose-Derived Stem Cell Modulation of T-Cell Regulation Correlates with Heme Oxgenase-1 Pathway Changes. Plast. Reconstr. Surg. 2016, 138, 1015–1023. [Google Scholar] [CrossRef]
- Stasch, T.; Hoehne, J.; Huynh, T.; De Baerdemaeker, R.; Grandel, S.; Herold, C. Débridement and Autologous Lipotransfer for Chronic Ulceration of the Diabetic Foot and Lower Limb Improves Wound Healing. Plast. Reconstr. Surg. 2015, 136, 1357–1366. [Google Scholar] [CrossRef]
- Lin, G.; Wang, G.; Liu, G.; Yang, L.-J.; Chang, L.-J.; Lue, T.F.; Lin, C.-S. Treatment of Type 1 Diabetes With Adipose Tissue–Derived Stem Cells Expressing Pancreatic Duodenal Homeobox 1. Stem Cells Dev. 2009, 18, 1399–1406. [Google Scholar] [CrossRef]
- Stessuk, T.; Puzzi, M.B.; Chaim, E.A.; Alves, P.C.M.; de Paula, E.V.; Forte, A.; Izumizawa, J.M.; Oliveira, C.C.; Frei, F.; Ribeiro-Paes, J.T. Platelet-rich plasma (PRP) and adipose-derived mesenchymal stem cells: Stimulatory effects on proliferation and migration of fibroblasts and keratinocytes in vitro. Arch. Dermatol. Res. 2016, 308, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Doornaert, M.; Colle, J.; De Maere, E.; Declercq, H.; Blondeel, P. Autologous fat grafting: Latest insights. Ann. Med. Surg. 2019, 37, 47–53. [Google Scholar] [CrossRef]
- Reinisch, K.B.; Zuk, G.; Raptis, D.A.; Bueter, M.; Guggenheim, M.; Stasch, T.; Palma, A.F. Autologous lipotransfer for bone defects secondary to osteomyelitis: A report of a novel method and systematic review of the literature. Int. Wound J. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zellner, E.G.; Pfaff, M.J.; Steinbacher, D.M. Fat Grafting in Primary Cleft Lip Repair. Plast. Reconstr. Surg. 2015, 135, 1449–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascali, M.; Quarato, D.; Marianetti, T.; Carinci, F. Malar region rejuvenation through non-invasive techniques: Hyaluronic acid fillers and lipofilling. J. Biol. Regul. Homeost. Agents 2017, 31, 1–7. [Google Scholar] [PubMed]
- Russe, E.; Kholosy, H.; Weitgasser, L.; Brandstetter, M.; Traintinger, H.; Neureiter, J.; Wechselberger, G.; Schoeller, T. Autologous fat grafting for enhancement of breast reconstruction with a transverse myocutaneous gracilis flap: A cohort study. J. Plast. Reconstr. Aesthetic Surg. 2018, 71, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Lazzeri, D.; Grassetti, L.; Silvestri, A.; Perdanasari, A.T.; Han, S.; Torresetti, M.; Di Benedetto, G.; Castello, M.F. Three-dimensional superficial liposculpture of the hips, flank, and thighs. Plast. Reconstr. Surg. Glob. Open 2015, 3, 25–35. [Google Scholar] [CrossRef]
- Chang, J.; Liao, Z.; Lu, M.; Meng, T.; Han, W.; Ding, C. Systemic and local adipose tissue in knee osteoarthritis. Osteoarthr. Cartil. 2018, 26, 864–871. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Keenan, S.R.; Lynn, J.; McEwan, J.C.; Beck, C.W. Gremlin1 induces anterior-posterior limb bifurcations in developing Xenopus limbs but does not enhance limb regeneration. Mech. Dev. 2015, 138, 256–267. [Google Scholar] [CrossRef]
- Woalder Calponin Isoforms CNN1, CNN2 and CNN3: Regulators for Actin Cytoskeleton Functions in Smooth Muscle and Non-Muscle Cells. Physiol. Behav. 2017, 176, 139–148.
- Hofsteen, P.; Strash, N.; Palpant, N.; Moon, R.T.; Pabon, L.; Murry, E.; Hofsteen, P.; Robitaille, A.M.; Strash, N.; Palpant, N.; et al. ALPK2 Promotes Cardiogenesis in Zebrafish and Human Pluripotent Stem Cells ALPK2 Promotes Cardiogenesis in Zebrafish and Human Pluripotent Stem Cells. ISCIENCE 2018, 2, 88–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wei, Y.; Yan, Y.; Wang, H.; Yang, J.; Zheng, Z.; Zha, J.; Bo, P.; Tang, Y.; Guo, X.; et al. CircDOCK1 suppresses cell apoptosis via inhibition of miR-196a-5p by targeting BIRC3 in OSCC. Oncol. Rep. 2018, 39, 951–966. [Google Scholar] [CrossRef] [Green Version]
- Skuntz, S.; Mankoo, B.; Nguyen, M.-T.T.; Hustert, E.; Nakayama, A.; Tournier-Lasserve, E. Lack of the mesodermal homeodomain protein MEOX1 disrupts sclerotome polarity and leads to a remodeling of the cranio-cervical joints of the axial skeleton. Bone 2008, 23, 383–395. [Google Scholar]
- Zhuang, Q.; Cheng, K.; Ming, Y. CX3CL1/CX3CR1 Axis, as the Therapeutic Potential in Renal Diseases: Friend or Foe? Curr. Gene Ther. 2018, 17, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Hauser, M.A.; Legler, D.F. Common and biased signaling pathways of the chemokine receptor CCR7 elicited by its ligands CCL19 and CCL21 in leukocytes. J. Leukoc. Biol. 2016, 99, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Jayasuriya, C.T. Role of Inflammation in Osteoarthritis. Rheumatol. Curr. Res. 2013, 03, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Jun, I.; Park, H.S.; Piao, H.; Han, J.W.; An, M.J.; Yun, B.G.; Zhang, X.; Cha, Y.H.; Shin, Y.K.; Yook, J.I.; et al. ANO9/TMEM16j promotes tumourigenesis via EGFR and is a novel therapeutic target for pancreatic cancer. Br. J. Cancer 2017, 117, 1798–1809. [Google Scholar] [CrossRef]
- Kalliolias, G.D.; Ivashkiv, L.B. Program, T.D. strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef]
- Uccelli, A.; de Rosbo, N.K. The immunomodulatory function of mesenchymal stem cells: Mode of action and pathways. Ann. N. Y. Acad. Sci. 2015, 1351, 114–126. [Google Scholar] [CrossRef]
- Frese, L.; Dijkman, P.E.; Hoerstrup, S.P. Adipose tissue-derived stem cells in regenerative medicine. Transfus. Med. Hemotherapy 2016, 43, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Kochumon, S.; Al-Ozairi, E.; Tuomilehto, J.; Ahmad, R. Association between Adipose Tissue Interleukin-33 and Immunometabolic Markers in Individuals with Varying Degrees of Glycemia. Dis. Markers 2019, 2019, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef]
- Wu, L.; Cai, X.; Zhang, S.; Karperien, M.; Lin, Y. Regeneration of articular cartilage by adipose tissue derived mesenchymal stem cells: Perspectives from stem cell biology and molecular medicine. J. Cell. Physiol. 2013, 228, 938–944. [Google Scholar] [CrossRef]
- Locker, M.; Kellermann, O.; Boucquey, M.; Khun, H.; Huerre, M.; Poliard, A. Paracrine and Autocrine Signals Promoting Full Chondrogenic Differentiation of a Mesoblastic Cell Line. J. Bone Miner. Res. 2003, 19, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, S.; Jennewein, M.; Bubel, M.; Guthoerl, S.; Pohlemann, T.; Oberringer, M. Interacting adipose-derived stem cells and microvascular endothelial cells provide a beneficial milieu for soft tissue healing. Mol. Biol. Rep. 2020, 1, 111–122. [Google Scholar] [CrossRef]
- Lombardi, F.; Palumbo, P.; Augello, F.R.; Cifone, M.G.; Cinque, B.; Giuliani, M. Secretome of adipose tissue-derived stem cells (ASCs) as a novel trend in chronic non-healing wounds: An overview of experimental in vitro and in vivo studies and methodological variables. Int. J. Mol. Sci. 2019, 20, 3721. [Google Scholar] [CrossRef] [Green Version]
- Campanella, C.; Caruso Bavisotto, C.; Logozzi, M.; Marino Gammazza, A.; Mizzoni, D.; Cappello, F.; Fais, S. On the choice of the extracellular vesicles for therapeutic purposes. Int. J. Mol. Sci. 2019, 20, 236. [Google Scholar] [CrossRef] [Green Version]
- Ragni, E.; Orfei, C.P.; De Luca, P.; Colombini, A.; Viganò, M.; Lugano, G.; de Girolamo, L. Identification of miRNA Reference Genes in Extracellular Vesicles from Adipose Derived Mesenchymal Stem Cells for Studying Osteoarthritis. Int. J. Mol. Sci. 2019, 20, 1108. [Google Scholar] [CrossRef] [Green Version]
- Tofiño-Vian, M.; Guillén, M.I.; Pérez Del Caz, M.D.; Castejón, M.A.; Alcaraz, M.J. Extracellular vesicles from adipose-derived mesenchymal stem cells downregulate senescence features in osteoarthritic osteoblasts. Oxid. Med. Cell. Longev. 2017, 2017, 7197598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schelbergen, R.F.; van Dalen, S.; ter Huurne, M.; Roth, J.; Vogl, T.; Noël, D.; Jorgensen, C.; van den Berg, W.B.; van de Loo, F.A.; Blom, A.B.; et al. Treatment efficacy of adipose-derived stem cells in experimental osteoarthritis is driven by high synovial activation and reflected by S100A8/A9 serum levels. Osteoarthr. Cartil. 2014, 22, 1158–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, H.; Tanaka, H.; Katoh, K.; Nakamura, H.; Nagashima, M.; Yoshino, S. Characterization of infiltrating T cells and Th1/Th2-type cytokines in the synovium of patients with osteoarthritis. Osteoarthr. Cartil. 2002, 10, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.; Butler, P.E.; Seifalian, A.M. Adipose-derived stem cells for clinical applications: A review. Cell Prolif. 2011, 44, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Domenis, R.; Cifù, A.; Quaglia, S.; Pistis, C.; Moretti, M.; Vicario, A.; Parodi, P.C.; Fabris, M.; Niazi, K.R.; Soon-Shiong, P.; et al. Pro inflammatory stimuli enhance the immunosuppressive functions of adipose mesenchymal stem cells-derived exosomes. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Seo, Y.; Shin, T.H.; Kim, H.S. Current Strategies to Enhance Adipose Stem Cell Function: An Update. Int. J. Mol. Sci. 2019, 20, 3827. [Google Scholar] [CrossRef] [Green Version]
- Taha, S.; Saller, M.M.; Haas, E.; Farkas, Z.; Aszodi, A.; Giunta, R.; Volkmer, E. Adipose-derived stem/progenitor cells from lipoaspirate: A comparison between the Liivage200-5 liposuction system and the Body-Jet liposuction system. J. Plas. Reconstr. Aesthet. Surg. 2020, 73, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome, 2011. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014, 15, 1–17. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ADSCs vs. ADSCs and TNFα | ||
|---|---|---|
| Gene symbol | Description | logFC |
| CXCL10 | C-X-C motif chemokine ligand 10 | 10.08 |
| CXCL11 | C-X-C motif chemokine ligand 11 | 9.67 |
| CCL5 | C-C motif chemokine ligand 5 | 9.60 |
| CXCL8 | C-X-C motif chemokine ligand 8 | 9.38 |
| LINC01539 | long intergenic non-protein coding RNA 1539 | 8.49 |
| PLA2G2A | phospholipase A2 group IIA | −6.22 |
| WISP2 | WNT1 inducible signaling pathway protein 2 | −6.47 |
| TNNT3 | troponin T3, fast skeletal type | −6.48 |
| ASPN | Asporin | −7.06 |
| H19 | H19, imprinted maternally expressed transcript | −8.17 |
| L-WAT vs. L-WAT and TNFα | ||
| Gene symbol | Description | logFC |
| CCL22 | C-C motif chemokine ligand 22 | 5.68 |
| ANO9 | anoctamin 9 | 5.30 |
| MMP9 | matrix metallopeptidase 9 | 5.17 |
| EBI3 | Epstein–Barr virus induced 3 | 4.94 |
| CCL5 | C-C motif chemokine ligand 5 | 4.68 |
| ECSCR | endothelial cell surface expressed chemotaxis and apoptosis regulator | −2.97 |
| AC091939.1 | novel transcript | −3.13 |
| MNDA | myeloid cell nuclear differentiation antigen | −3.57 |
| CA4 | carbonic anhydrase 4 | −4.08 |
| AC002546.1 | novel transcript | −4.13 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taha, S.; Volkmer, E.; Haas, E.; Alberton, P.; Straub, T.; David-Rus, D.; Aszodi, A.; Giunta, R.; Saller, M.M. Differences in the Inflammatory Response of White Adipose Tissue and Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2020, 21, 1086. https://doi.org/10.3390/ijms21031086
Taha S, Volkmer E, Haas E, Alberton P, Straub T, David-Rus D, Aszodi A, Giunta R, Saller MM. Differences in the Inflammatory Response of White Adipose Tissue and Adipose-Derived Stem Cells. International Journal of Molecular Sciences. 2020; 21(3):1086. https://doi.org/10.3390/ijms21031086
Chicago/Turabian StyleTaha, Sara, Elias Volkmer, Elisabeth Haas, Paolo Alberton, Tobias Straub, Diana David-Rus, Attila Aszodi, Riccardo Giunta, and Maximilian Michael Saller. 2020. "Differences in the Inflammatory Response of White Adipose Tissue and Adipose-Derived Stem Cells" International Journal of Molecular Sciences 21, no. 3: 1086. https://doi.org/10.3390/ijms21031086
APA StyleTaha, S., Volkmer, E., Haas, E., Alberton, P., Straub, T., David-Rus, D., Aszodi, A., Giunta, R., & Saller, M. M. (2020). Differences in the Inflammatory Response of White Adipose Tissue and Adipose-Derived Stem Cells. International Journal of Molecular Sciences, 21(3), 1086. https://doi.org/10.3390/ijms21031086