Surgical Wound Fluids from Patients with Breast Cancer Reveal Similarities in the Biological Response Induced by Intraoperative Radiation Therapy and the Radiation-Induced Bystander Effect—Transcriptomic Approach

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
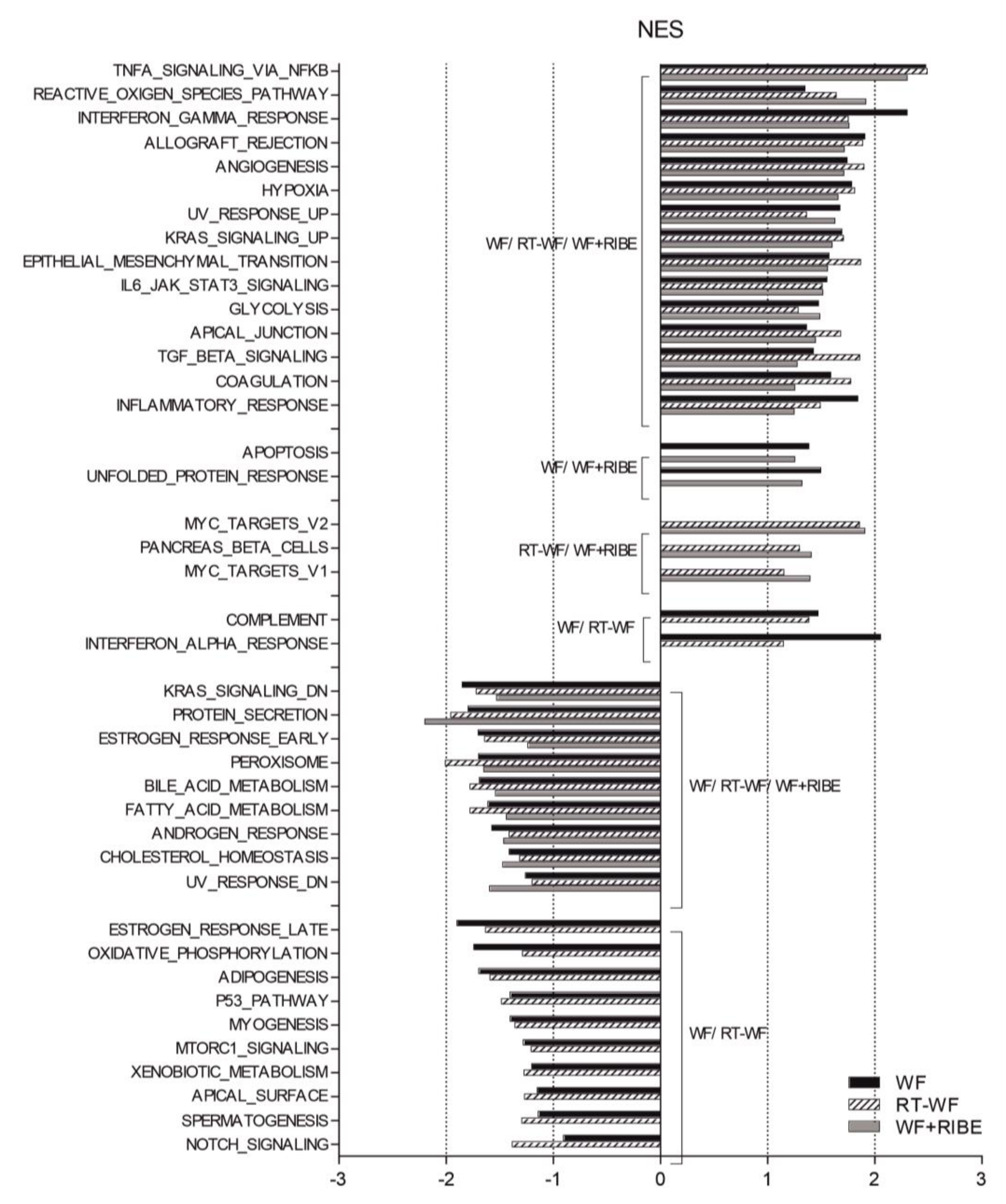
2.1. Breast Cancer Cells Treated with Surgical Wound Fluids Are Associated with an Enhancement of Processes Associated with Tumorigenicity and Metastatic Potential
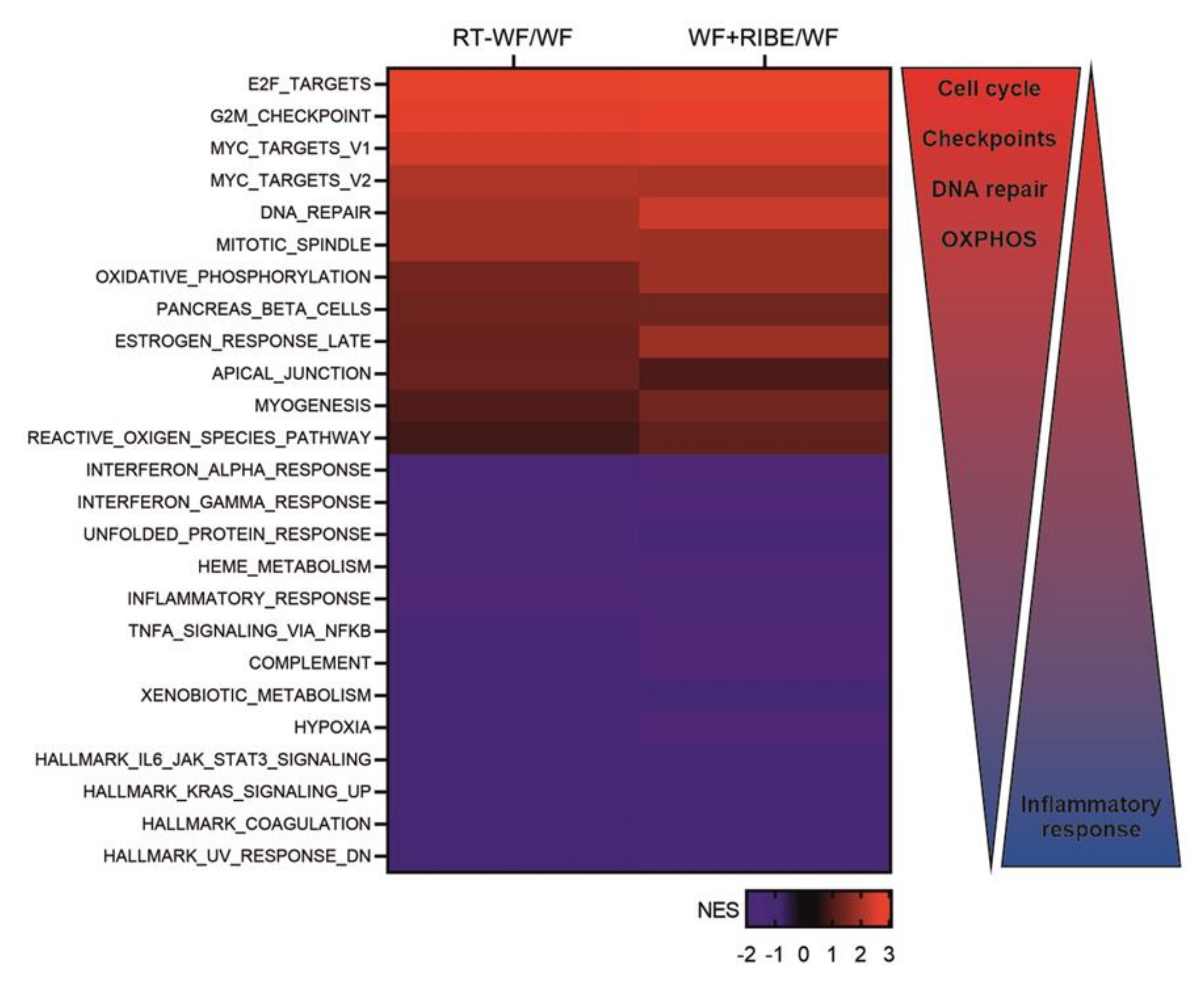
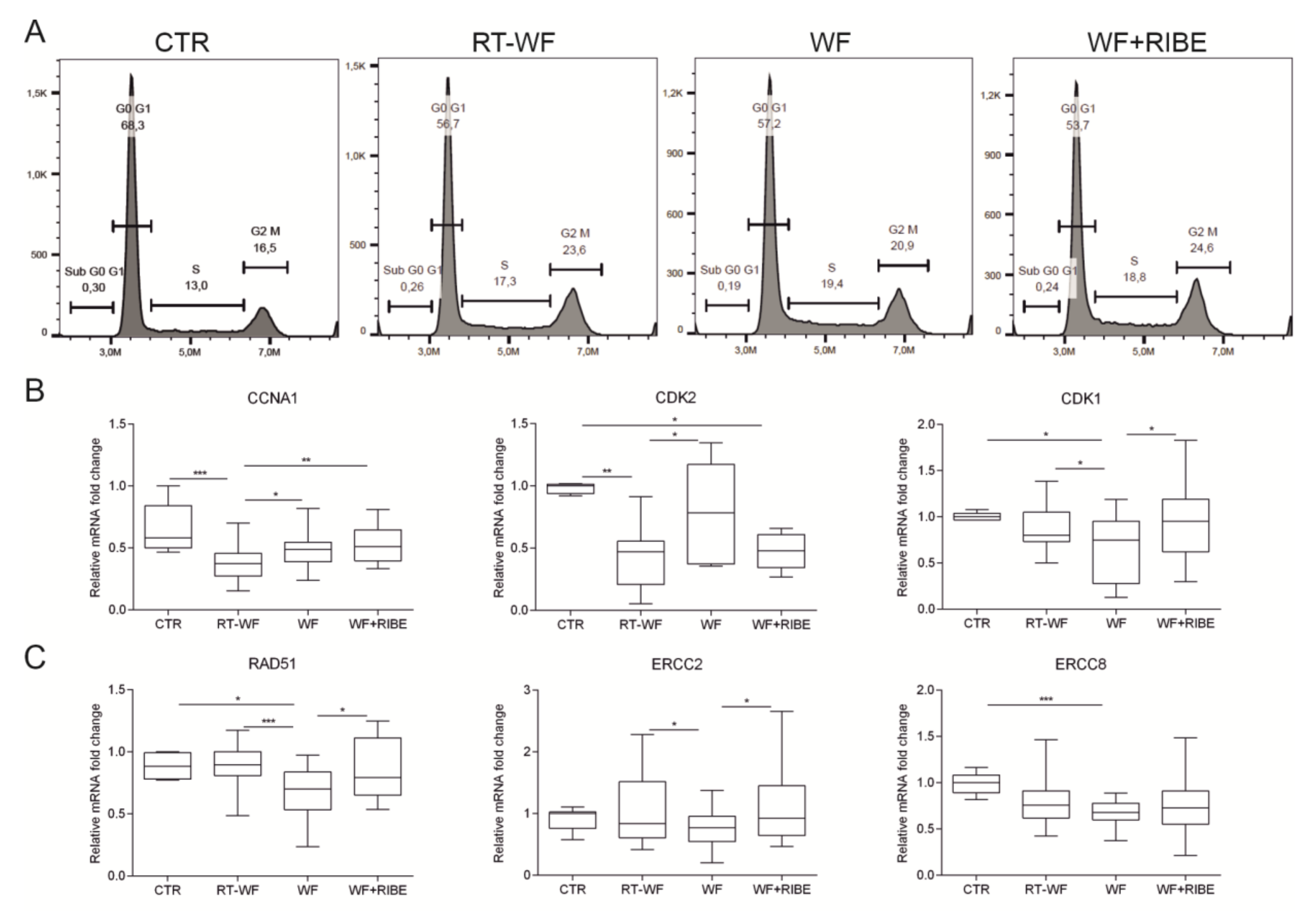
2.2. The Biological Processes in Cancer Cells Enriched in RT-WF and WF+RIBE-Treated Cells Are Similar and Related to Cell Proliferation, Cell Division, and DNA Damage Responses
2.3. Surgical Wound Fluids Alter the Metabolism of the MDA-MB-468 Cell Line
3. Discussion
4. Materials and Methods
4.1. Wound Fluid Collection
4.2. Cell Culture
4.3. Radiation-Induced Bystander Effect Medium (RIBE)
4.4. Cell Treatment
4.5. RNA Isolation
4.6. Microarray Expression Study
4.7. Microarray Data Analysis
4.8. Gene Set Enrichment Analysis (GSEA)
4.9. Real-Time Quantitative PCR RT-qPCR
4.10. Flow Cytometry
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ATP5G1 | ATP synthase membrane subunit c locus 1 |
| ATP5J2 | ATP synthase membrane subunit f |
| BC | breast cancer |
| BCS | breast conserving surgery |
| COX4I1 | cytochrome c oxidase subunit 4I1 |
| COX6B1 | cytochrome c oxidase subunit 6B1 |
| EBRT | external beam radiotherapy |
| EMT | epithelial to mesenchymal transition |
| ER | estrogen receptor |
| ES | enrichment score |
| FBS | fetal bovine serum |
| FC | fold change |
| GEO | Gene Expression Omnibus |
| GSEA | gene set enrichment analysis |
| HR | homologous recombination |
| IORT | intraoperative radiation therapy |
| IR | ionizing radiation |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LDH | lactic acid dehydrogenase |
| NDUFB2 | NADH:ubiquinone oxidoreductase subunit B2 |
| NDUFB7 | NADH:ubiquinone oxidoreductase subunit B7 |
| NER | nucleotide excision repair |
| NES | normalized enrichment score |
| PR | progesterone receptor |
| RIBE | radiation induced bystander effect |
| RT-WF | surgical wound fluids collected from patients after breast conserving surgery followed by IORT |
| SWF | surgical wound fluids |
| TNBC | triple-negative breast cancer |
| WF | surgical wound fluids collected from patients after breast conserving surgery |
References
- McGale, P.; Taylor, C.; Correa, C.; Cutter, D.; Duane, F.; Ewertz, M.; Wang, Y. Effect of radiotherapy after mastectomy and axillary surgery on 10-year recurrence and 20-year breast cancer mortality: Meta-analysis of individual patient data for 8135 women in 22 randomised trials. Lancet 2014, 383, 2127–2135. [Google Scholar] [CrossRef]
- Faverly, D.R.; Hendriks, J.H.; Holland, R. Breast carcinomas of limited extent: Frequency, radiologic-pathologic characteristics, and surgical margin requirements. Cancer 2001, 91, 647–659. [Google Scholar] [CrossRef]
- Lemanski, C.; Azria, D.; Thezenas, S.; Gutowski, M.; Saint-Aubert, B.; Rouanet, P.; Dubois, J.B. Intraoperative radiotherapy given as a boost for early breast cancer: Long-term clinical and cosmetic results. Int. J. Radiat. Oncol. Biol. Phys. 2006, 64, 1410–1415. [Google Scholar] [CrossRef]
- Sperk, E.; Welzel, G.; Keller, A.; Kraus-Tiefenbacher, U.; Gerhardt, A.; Sütterlin, M.; Wenz, F. Late radiation toxicity after intraoperative radiotherapy (IORT) for breast cancer: Results from the randomized phase III trial TARGIT A. Breast Cancer Res. Treat. 2012, 135, 253–260. [Google Scholar] [CrossRef]
- Fastner, G.; Sedlmayer, F.; Merz, F.; Deutschmann, H.; Reitsamer, R.; Menzel, C.; Mirri, A. IORT with electrons as boost strategy during breast conserving therapy in limited stage breast cancer: Long term results of an ISIORT pooled analysis. Radiother. Oncol. 2013, 108, 279–286. [Google Scholar] [CrossRef]
- Belletti, B.; Vaidya, J.S.; D’Andrea, S.; Entschladen, F.; Roncadin, M.; Lovat, F.; Veronesi, A. Targeted intraoperative radiotherapy impairs the stimulation of breast cancer cell proliferation and invasion caused by surgical wounding. Clin. Cancer Res. 2008, 14, 1325–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demicheli, R.; Valagussa, P.; Bonadonna, G. Does surgery modify growth kinetics of breast cancer micrometastases? Br. J. Cancer 2001, 85, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Agresti, R.; Triulzi, T.; Sasso, M.; Ghirelli, C.; Aiello, P.; Rybinska, I.; Bianchi, F. Wound Healing Fluid Reflects the Inflammatory Nature and Aggressiveness of Breast Tumors. Cells 2019, 8, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krall, J.A.; Reinhardt, F.; Mercury, O.A.; Pattabiraman, D.R.; Brooks, M.W.; Dougan, M.; Weinberg, R.A. The systemic response to surgery triggers the outgrowth of distant immune-controlled tumors in mouse models of dormancy. Sci. Transl. Med. 2018, 10, eaan3464. [Google Scholar] [CrossRef] [Green Version]
- Turdo, F.; Bianchi, F.; Gasparini, P.; Sandri, M.; Sasso, M.; De Cecco, L.; Agresti, R. CDCP1 is a novel marker of the most aggressive human triple-negative breast cancers. Oncotarget 2016, 7, 69649–69665. [Google Scholar] [CrossRef] [Green Version]
- Kulcenty, K.; Piotrowski, I.; Wróblewska, J.P.; Wasiewicz, J.; Suchorska, W.M. The Composition of Surgical Wound Fluids from Breast Cancer Patients is Affected by Intraoperative Radiotherapy Treatment and Depends on the Molecular Subtype of Breast Cancer. Cancers 2019, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Piotrowski, I.; Kulcenty, K.; Murawa, D.; Suchorska, W. Surgical wound fluids from patients treated with intraoperative radiotherapy induce radiobiological response in breast cancer cells. Med. Oncol. 2018, 36, 14. [Google Scholar] [CrossRef] [Green Version]
- Kulcenty, K.; Piotrowski, I.; Zaleska, K.; Wichtowski, M.; Wróblewska, J.; Murawa, D.; Suchorska, W.M. Wound fluids collected postoperatively from patients with breast cancer induce epithelial to mesenchymal transition but intraoperative radiotherapy impairs this effect by activating the radiation-induced bystander effect. Sci. Rep. 2019, 9, 7891. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B. Aerobic glycolysis and high level of lactate in cancer metabolism and microenvironment. Genes Dis. 2017, 4, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Troester, M.A.; Lee, M.H.; Carter, M.; Fan, C.; Cowan, D.W.; Perez, E.R.; Schneider, S.S. Activation of host wound responses in breast cancer microenvironment. Clin. Cancer Res. 2009, 15, 7020–7028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliabue, E.; Agresti, R.; Carcangiu, M.L.; Ghirelli, C.; Morelli, D.; Campiglio, M.; Ménard, S. Role of HER2 in wound-induced breast carcinoma proliferation. Lancet 2003, 362, 527–533. [Google Scholar] [CrossRef]
- Piotrowski, I.; Kulcenty, K.; Wichtowski, M.; Murawa, D.; Suchorska, W. Intraoperative Radiotherapy of Breast Cancer and Its Biological Effects. Breast Care 2017, 12, 109–113. [Google Scholar] [CrossRef]
- Ren, B.; Cam, H.; Takahashi, Y.; Volkert, T.; Terragni, J.; Young, R.A.; Dynlacht, B.D. E2F integrates cell cycle progression with DNA repair, replication, and G(2)/M checkpoints. Genes Dev. 2002, 16, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Warburg, O.; Wind, F.; Negelein, E. The Metabolism of Tumors in the Body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Estrella, V.; Chen, T.; Lloyd, M.; Wojtkowiak, J.; Cornnell, H.H.; Ibrahim-Hashim, A.; Johnson, J. Acidity generated by the tumor microenvironment drives local invasion. Cancer Res. 2013, 73, 1524–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.H.; Qiu, J.; O’Sullivan, D.; Buck, M.D.; Noguchi, T.; Curtis, J.D.; Tonc, E. Metabolic Competition in the Tumor Microenvironment Is a Driver of Cancer Progression. Cell 2015, 162, 1229–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Tan, M.; Cai, Q. The Warburg effect in tumor progression: Mitochondrial oxidative metabolism as an anti-metastasis mechanism. Cancer Lett. 2015, 356, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polyak, K.; Weinberg, R.A. Transitions between epithelial and mesenchymal states: Acquisition of malignant and stem cell traits. Nat. Rev. Cancer 2009, 9, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; PZhou, B. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Pitroda, S.P.; Wakim, B.T.; Sood, R.F.; Beveridge, M.G.; Beckett, M.A.; MacDermed, D.M.; Khodarev, N.N. STAT1-dependent expression of energy metabolic pathways links tumour growth and radioresistance to the Warburg effect. BMC Med. 2009, 7, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodarev, N.N.; Beckett, M.; Labay, E.; Darga, T.; Roizman, B.; Weichselbaum, R.R. STAT1 is overexpressed in tumors selected for radioresistance and confers protection from radiation in transduced sensitive cells. Proc. Natl. Acad. Sci. USA 2004, 101, 1714–1719. [Google Scholar] [CrossRef] [Green Version]
- Jopek, K.; Tyczewska, M.; Celichowski, P.; Malendowicz, L.K.; Rucinski, M. Transcriptome Profile in Unilateral Adrenalectomy-Induced Compensatory Adrenal Growth in the Rat. Int. J. Mol. Sci. 2018, 19, 1111. [Google Scholar] [CrossRef] [Green Version]
- Stelcer, E.; Kulcenty, K.; Rucinski, M.; Jopek, K.; Richter, M.; Trzeciak, T.; Suchorska, W.M. Chondrogenic differentiation in vitro of hiPSCs activates pathways engaged in limb development. Stem Cell Res. 2018, 30, 53–60. [Google Scholar] [CrossRef]
- Stelcer, E.; Kulcenty, K.; Rucinski, M.; Jopek, K.; Trzeciak, T.; Richter, M.; Suchorska, W.M. Expression of Pluripotency Genes in Chondrocyte-Like Cells Differentiated from Human Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2018, 19, 550. [Google Scholar] [CrossRef]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy—Analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, B.S.; Irizarry, R.A. A framework for oligonucleotide microarray preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.I.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucl. Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Functions Simplifying Presentation of PCA Models in a 3D Interactive Representation Using ‘rgl’. 2017. Available online: https://cran.r-project.org/web/packages/pca3d (accessed on 28 April 2019).
- Package “Corrplot”: Visualization of a Correlation Matrix. 2019. Available online: https://github.com/taiyun/corrplot (accessed on 28 April 2019).
- Damian, D.; Gorfine, M. Statistical concerns about the GSEA procedure. Nat. Genet. 2004, 36, 663. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Mesirov, J.P. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Comparisons | NES | NOM p-Val | FDR q-Val |
|---|---|---|---|---|
| Glycolysis | RT-WF vs. CTR | 1.29 | 0.035 | 0.074 |
| WF vs. CTR | 1.48 | 0.002 | 0.017 | |
| WF+RIBE vs. CTR | 1.49 | 0.002 | 0.024 | |
| Fatty acid metabolism | RT-WF vs. CTR | −1.78 | 0.000 | 0.004 |
| WF vs. CTR | −1.61 | 0.000 | 0.004 | |
| WF+RIBE vs. CTR | −1.44 | 0.009 | 0.042 | |
| Oxidative phosphorylation | RT-WF vs. WF | 1.42 | 0.005 | 0.043 |
| WF+RIBE vs. WF | 1.88 | 0.000 | 0.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulcenty, K.; Piotrowski, I.; Rucinski, M.; Wroblewska, J.P.; Jopek, K.; Murawa, D.; Suchorska, W.M. Surgical Wound Fluids from Patients with Breast Cancer Reveal Similarities in the Biological Response Induced by Intraoperative Radiation Therapy and the Radiation-Induced Bystander Effect—Transcriptomic Approach. Int. J. Mol. Sci. 2020, 21, 1159. https://doi.org/10.3390/ijms21031159
Kulcenty K, Piotrowski I, Rucinski M, Wroblewska JP, Jopek K, Murawa D, Suchorska WM. Surgical Wound Fluids from Patients with Breast Cancer Reveal Similarities in the Biological Response Induced by Intraoperative Radiation Therapy and the Radiation-Induced Bystander Effect—Transcriptomic Approach. International Journal of Molecular Sciences. 2020; 21(3):1159. https://doi.org/10.3390/ijms21031159
Chicago/Turabian StyleKulcenty, Katarzyna, Igor Piotrowski, Marcin Rucinski, Joanna Patrycja Wroblewska, Karol Jopek, Dawid Murawa, and Wiktoria Maria Suchorska. 2020. "Surgical Wound Fluids from Patients with Breast Cancer Reveal Similarities in the Biological Response Induced by Intraoperative Radiation Therapy and the Radiation-Induced Bystander Effect—Transcriptomic Approach" International Journal of Molecular Sciences 21, no. 3: 1159. https://doi.org/10.3390/ijms21031159
APA StyleKulcenty, K., Piotrowski, I., Rucinski, M., Wroblewska, J. P., Jopek, K., Murawa, D., & Suchorska, W. M. (2020). Surgical Wound Fluids from Patients with Breast Cancer Reveal Similarities in the Biological Response Induced by Intraoperative Radiation Therapy and the Radiation-Induced Bystander Effect—Transcriptomic Approach. International Journal of Molecular Sciences, 21(3), 1159. https://doi.org/10.3390/ijms21031159