Low Dose of Penfluridol Inhibits VEGF-Induced Angiogenesis

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Cell Lines and Culture Procedures
2.3. MTT Cytotoxicity Assay
2.4. Cell Migration Assay
2.5. Tube Formation Assay
2.6. In Vivo Matrigel Plug and Hemoglobin Assay
2.7. Cancer Cell Paracrine Effect on Angiogenesis
2.8. Wound Healing Assay
2.9. Immunoblot Analysis
2.10. Statistical Analysis
3. Results
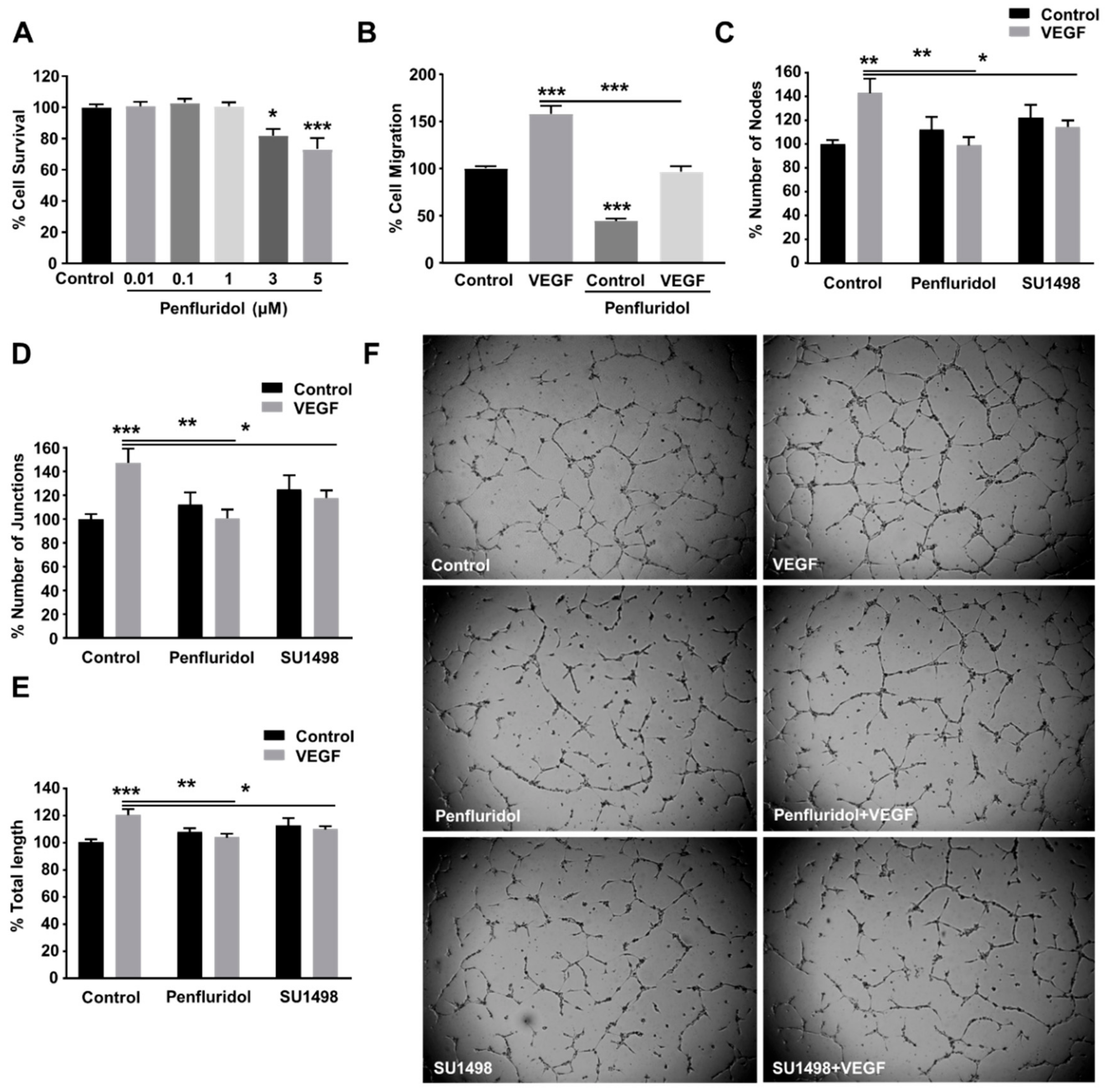
3.1. Identification of Non-Toxic Penfluridol Concentrations
3.2. Low Concentration of Penfluridol Inhibits Endothelial Cell Migration and Tube Formation In Vitro
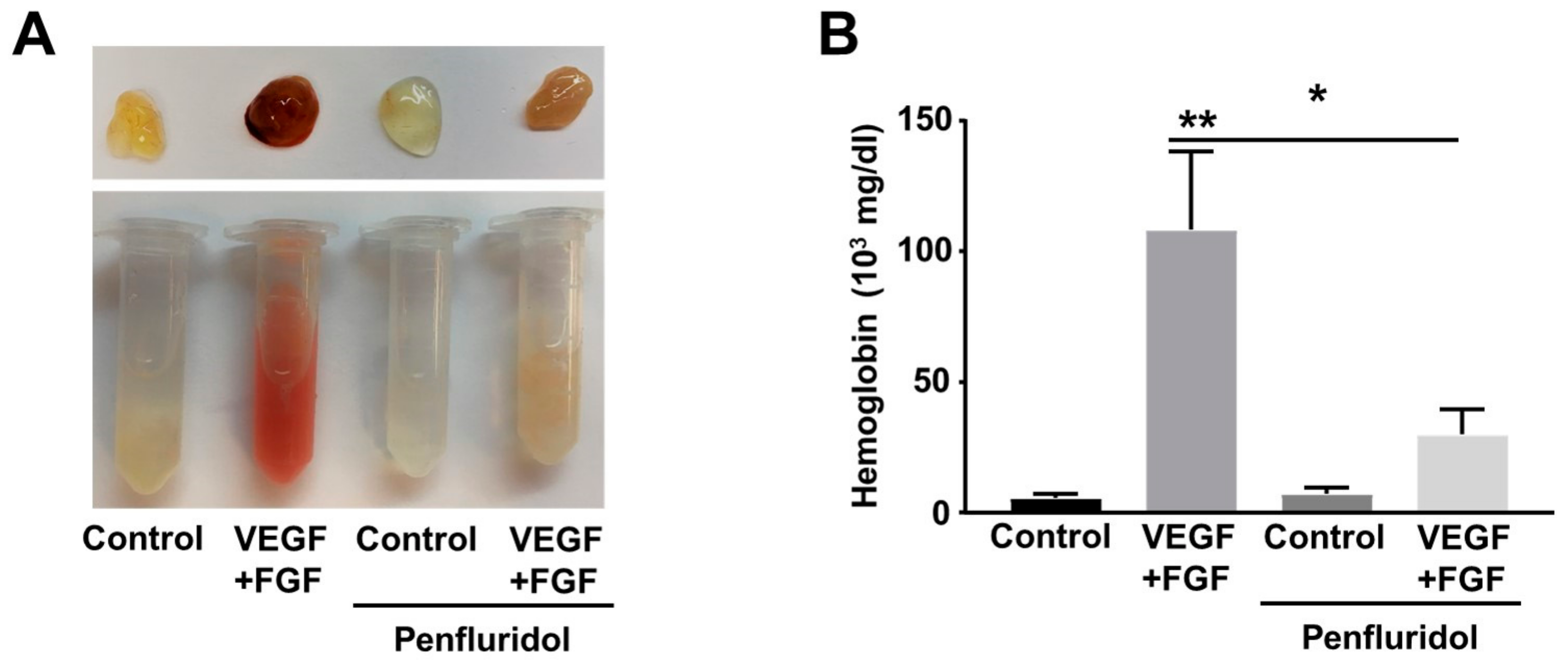
3.3. Low Dose of Penfluridol Blocks In Vivo Angiogenesis
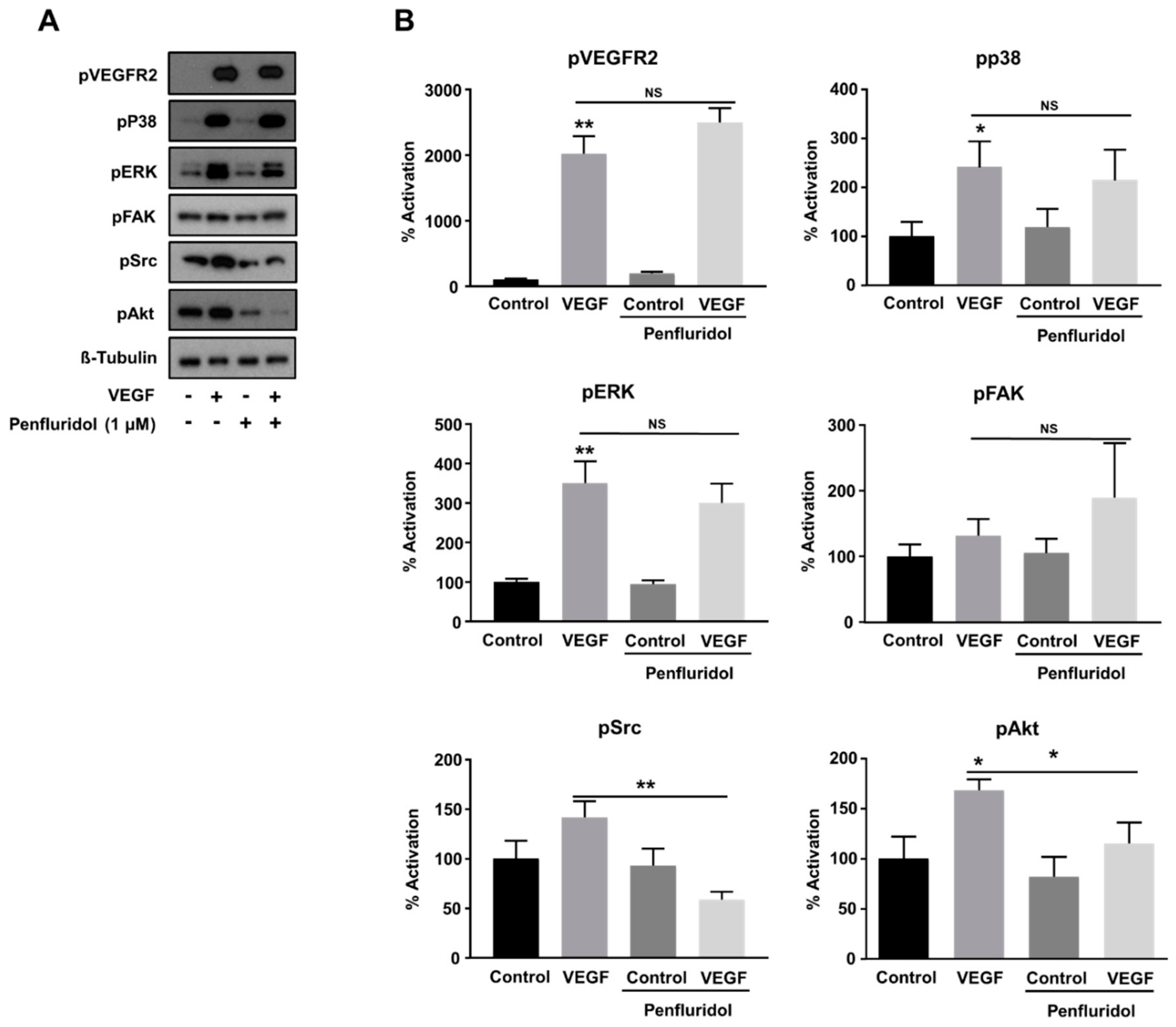
3.4. Low Concentration of Penfluridol Affects the Downstream VEGF-Signaling Pathway in Endothelial Cells
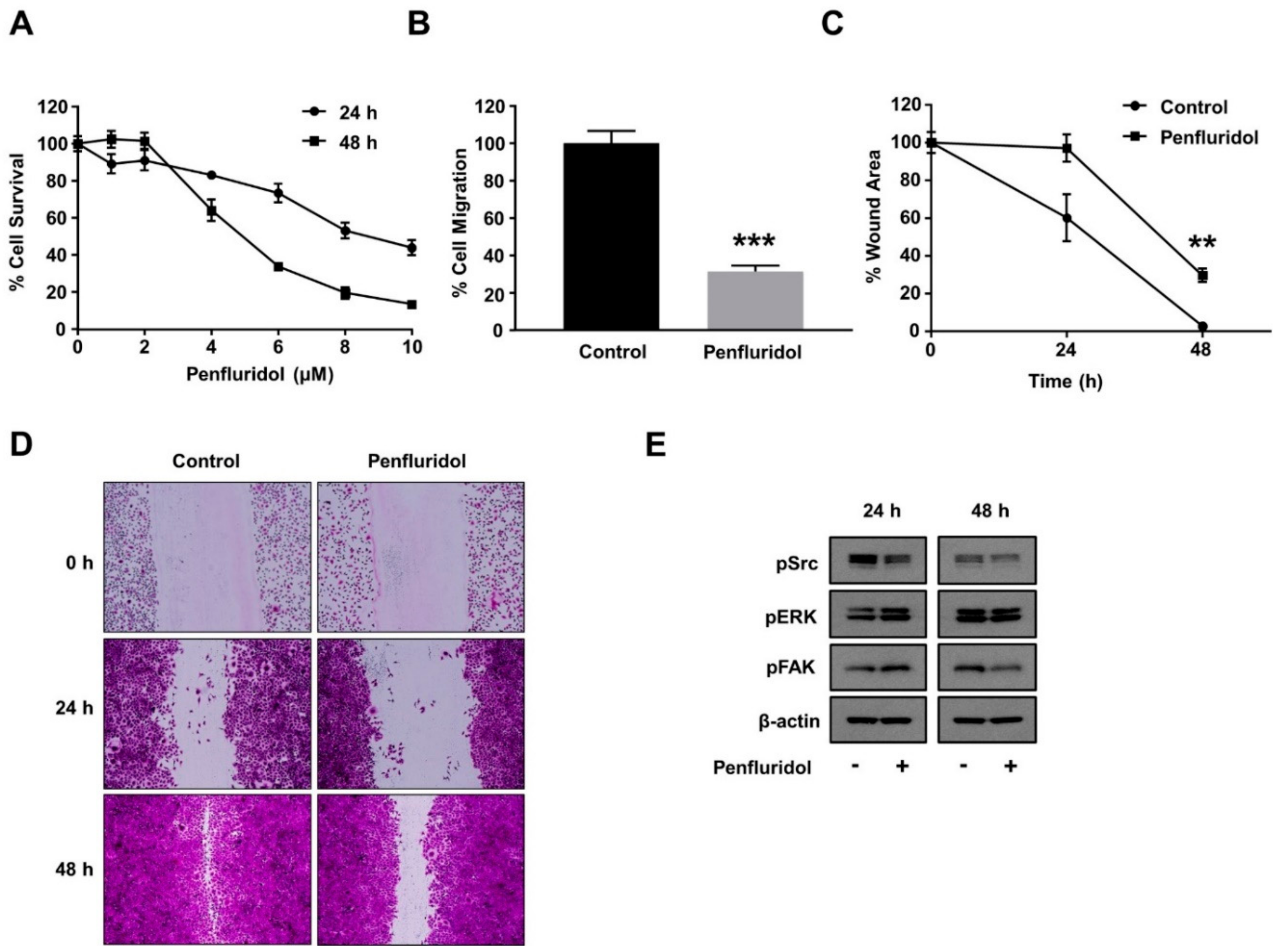
3.5. Effect of Low Concentration of Penfluridol on Breast Cancer Cell Functions and Basal Src Activation
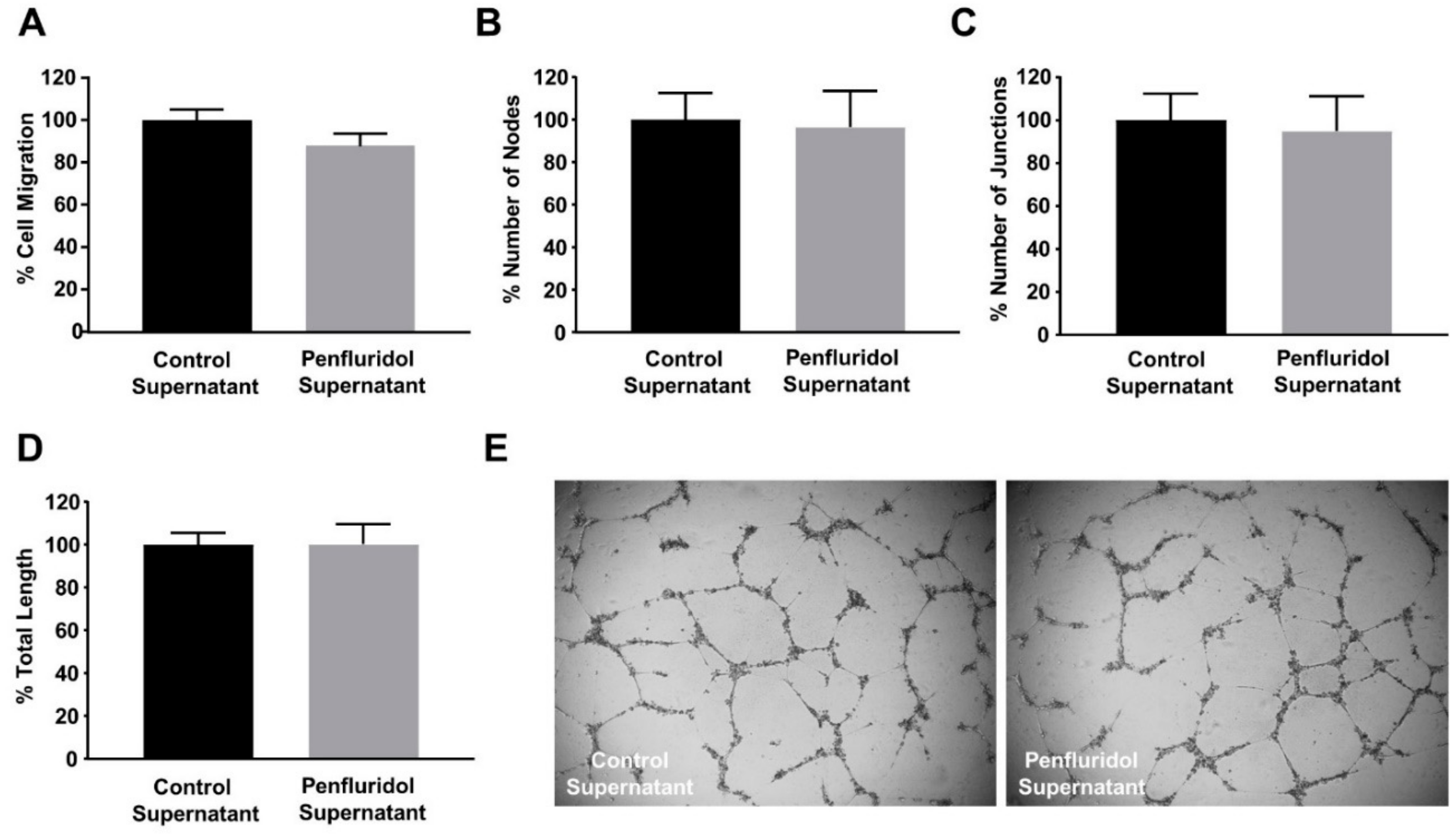
3.6. The Effect of Low Concentration of Penfluridol on Endothelial and Breast Cancer Cells Are Not Dependent on Cancer Cell-Derived Paracrine Effect on Endothelial Cells
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Ataollahi, M.R.; Sharifi, J.; Paknahad, M.R.; Paknahad, A. Breast cancer and associated factors: a review. J. Med. Life 2015, 8, 6–11. [Google Scholar] [PubMed]
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Thun, M.J. Cancer statistics. 2009. CA Cancer J. Clin. 2009, 59, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Rosso, K.; Nathanson, S.D. Pathogenesis, prevention, diagnosis and treatment of breast cancer. World J. Clin. Oncol. 2014, 5, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Bielenberg, D.R.; Zetter, B.R. The Contribution of Angiogenesis to the Process of Metastasis. Cancer J. 2015, 21, 267–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P. Mechanisms of angiogenesis and arteriogenesis. Nat. Med. 2000, 6, 389–395. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis: an organizing principle for drug discovery? Nat. Rev. Drug Discov. 2007, 6, 273–286. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Dudley, A.C. Tumor endothelial cells. Cold Spring Harb. Perspect. Med. 2012, 2, a006536. [Google Scholar] [CrossRef]
- Walker, C.; Mojares, E.; Del Rio Hernandez, A. Role of Extracellular Matrix in Development and Cancer Progression. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Wong, P.P.; Bodrug, N.; Hodivala-Dilke, K.M. Exploring Novel Methods for Modulating Tumor Blood Vessels in Cancer Treatment. Curr. Biol. 2016, 26, R1161–R1166. [Google Scholar] [CrossRef] [PubMed]
- Mattheolabakis, G.; Mikelis, C.M. Nanoparticle Delivery and Tumor Vascular Normalization: The Chicken or The Egg? Front. Oncol. 2019, 9, 1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, Y. Tumor angiogenesis and anti-angiogenic therapy. Keio J. Med. 2012, 61, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, L.S. VEGF-targeted therapy: therapeutic potential and recent advances. Oncologist 2005, 10, 382–391. [Google Scholar] [CrossRef]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2019, 76, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Zahra, F.T.; Sajib, M.S.; Ichiyama, Y.; Akwii, R.G.; Tullar, P.E.; Cobos, C.; Minchew, S.A.; Doci, C.L.; Zheng, Y.; Kubota, Y.; et al. Endothelial RhoA GTPase is essential for in vitro endothelial functions but dispensable for physiological in vivo angiogenesis. Sci. Rep. 2019, 9, 11666. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Koch, S.; Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Cold Spring Harb. Perspect. Med. 2012, 2, a006502. [Google Scholar] [CrossRef]
- Claesson-Welsh, L.; Welsh, M. VEGFA and tumour angiogenesis. J. Intern. Med. 2013, 273, 114–127. [Google Scholar] [CrossRef]
- Jaszai, J.; Schmidt, M.H.H. Trends and Challenges in Tumor Anti-Angiogenic Therapies. Cells 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Joyce, J.A. Therapeutic targeting of the tumor microenvironment. Cancer Cell 2005, 7, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roma-Rodrigues, C.; Mendes, R.; Baptista, P.V.; Fernandes, A.R. Targeting Tumor Microenvironment for Cancer Therapy. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.I.; Larson, C.; Treul, S.J. Study of penfluridol and chlorpromazine in the treatment of chronic schizophrenia. J. Clin. Pharmacol. 1982, 22, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Nikvarz, N.; Vahedian, M.; Khalili, N. Chlorpromazine versus penfluridol for schizophrenia. Cochrane Database Syst. Rev. 2017, 9, CD011831. [Google Scholar] [CrossRef] [PubMed]
- Pich, J. Review: Chlorpromazine Versus Penfluridol for Schizophrenia. Issues Ment Health Nurs. 2018, 39, 814–815. [Google Scholar] [CrossRef] [PubMed]
- Zur, E. Penfluridol, a Unique Psychiatric Medicine for the Treatment of Chronic Schizophrenia. Int. J. Pharm. Compd. 2019, 23, 113–119. [Google Scholar] [PubMed]
- Ranjan, A.; Gupta, P.; Srivastava, S.K. Penfluridol: An Antipsychotic Agent Suppresses Metastatic Tumor Growth in Triple-Negative Breast Cancer by Inhibiting Integrin Signaling Axis. Cancer Res. 2016, 76, 877–890. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Srivastava, S.K. Penfluridol suppresses pancreatic tumor growth by autophagy-mediated apoptosis. Sci. Rep. 2016, 6, 26165. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Srivastava, S.K. Penfluridol suppresses glioblastoma tumor growth by Akt-mediated inhibition of GLI1. Oncotarget 2017, 8, 32960–32976. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, E.; Li, X.; Safe, S. Penfluridol Represses Integrin Expression in Breast Cancer through Induction of Reactive Oxygen Species and Downregulation of Sp Transcription Factors. Mol. Cancer Ther. 2017, 16, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Mikelis, C.; Sfaelou, E.; Koutsioumpa, M.; Kieffer, N.; Papadimitriou, E. Integrin alpha(v)beta(3) is a pleiotrophin receptor required for pleiotrophin-induced endothelial cell migration through receptor protein tyrosine phosphatase beta/zeta. FASEB J. 2009, 23, 1459–1469. [Google Scholar] [CrossRef] [PubMed]
- Hossian, A.; Sajib, M.S.; Tullar, P.E.; Mikelis, C.M.; Mattheolabakis, G. Multipronged activity of combinatorial miR-143 and miR-506 inhibits Lung Cancer cell cycle progression and angiogenesis in vitro. Sci. Rep. 2018, 8, 10495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokolakis, G.; Mikelis, C.; Papadimitriou, E.; Courty, J.; Karetsou, E.; Katsoris, P. Effect of heparin affin regulatory peptide on the expression of vascular endothelial growth factor receptors in endothelial cells. In Vivo 2006, 20, 629–635. [Google Scholar]
- Mikelis, C.M.; Palmby, T.R.; Simaan, M.; Li, W.; Szabo, R.; Lyons, R.; Martin, D.; Yagi, H.; Fukuhara, S.; Chikumi, H.; et al. PDZ-RhoGEF and LARG are essential for embryonic development and provide a link between thrombin and LPA receptors and Rho activation. J. Biol. Chem. 2013, 288, 12232–12243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikelis, C.; Lamprou, M.; Koutsioumpa, M.; Koutsioubas, A.G.; Spyranti, Z.; Zompra, A.A.; Spiliopoulos, N.; Vradis, A.A.; Katsoris, P.; Spyroulias, G.A.; et al. A peptide corresponding to the C-terminal region of pleiotrophin inhibits angiogenesis in vivo and in vitro. J. Cell. Biochem. 2011, 112, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, G.; Martinelli, M.; Courty, J.; Cascone, I. Angiogenesis analyzer for ImageJ. In Proceedings of the 4th ImageJ User and Developer Conference Proceedings, Luxembourg, 24–26 October 2012; pp. 198–201. [Google Scholar]
- Doci, C.L.; Mikelis, C.M.; Lionakis, M.S.; Molinolo, A.A.; Gutkind, J.S. Genetic Identification of SEMA3F as an Antilymphangiogenic Metastasis Suppressor Gene in Head and Neck Squamous Carcinoma. Cancer Res. 2015, 75, 2937–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, D.H.; Kim, M.R.; Jang, J.H.; Na, H.J.; Lee, S. A Review of Anti-Angiogenic Targets for Monoclonal Antibody Cancer Therapy. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Ferla, R.; Bonomi, M.; Otvos, L., Jr.; Surmacz, E. Glioblastoma-derived leptin induces tube formation and growth of endothelial cells: comparison with VEGF effects. BMC Cancer 2011, 11, 303. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.Q.; Lee, D.Y.; Zioncheck, T.F. Vascular endothelial growth factor governs endothelial nitric-oxide synthase expression via a KDR/Flk-1 receptor and a protein kinase C signaling pathway. J. Biol. Chem. 1999, 274, 33057–33063. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, D.C.; Bryan, B.A. Anti-angiogenic therapy: adapting strategies to overcome resistant tumors. J. Cell. Biochem. 2010, 111, 543–553. [Google Scholar] [CrossRef]
- Bergers, G.; Hanahan, D. Modes of resistance to anti-angiogenic therapy. Nat. Rev. Cancer 2008, 8, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhide, R.S.; Lombardo, L.J.; Hunt, J.T.; Cai, Z.W.; Barrish, J.C.; Galbraith, S.; Jeyaseelan, R., Sr.; Mortillo, S.; Wautlet, B.S.; Krishnan, B.; et al. The antiangiogenic activity in xenograft models of brivanib, a dual inhibitor of vascular endothelial growth factor receptor-2 and fibroblast growth factor receptor-1 kinases. Mol. Cancer Ther. 2010, 9, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danastas, K.; Combes, V.; Lindsay, L.A.; Grau, G.E.; Thompson, M.B.; Murphy, C.R. VEGF111: new insights in tissue invasion. Front. Physiol. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Zhou, W. The Emerging Regulation of VEGFR-2 in Triple-Negative Breast Cancer. Front. Endocrinol. 2015, 6, 159. [Google Scholar] [CrossRef] [Green Version]
- Soares, B.G.; Lima, M.S. Penfluridol for schizophrenia. Cochrane Database Syst. Rev. 2006. [Google Scholar] [CrossRef]
- Gupta, N.; Gupta, P.; Srivastava, S.K. Penfluridol overcomes paclitaxel resistance in metastatic breast cancer. Sci. Rep. 2019, 9, 5066. [Google Scholar] [CrossRef]
- Ranjan, A.; German, N.; Mikelis, C.; Srivenugopal, K.; Srivastava, S.K. Penfluridol induces endoplasmic reticulum stress leading to autophagy in pancreatic cancer. Tumour. Biol. 2017, 39, 1010428317705517. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Wright, S.; Srivastava, S.K. Immune consequences of penfluridol treatment associated with inhibition of glioblastoma tumor growth. Oncotarget 2017, 8, 47632–47641. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Guan, J.L. Signal transduction by focal adhesion kinase in cancer. Cancer Metastasis Rev. 2009, 28, 35–49. [Google Scholar] [CrossRef]
- Eliceiri, B.P.; Paul, R.; Schwartzberg, P.L.; Hood, J.D.; Leng, J.; Cheresh, D.A. Selective requirement for Src kinases during VEGF-induced angiogenesis and vascular permeability. Mol. Cell 1999, 4, 915–924. [Google Scholar] [CrossRef]
- Sun, Z.; Li, X.; Massena, S.; Kutschera, S.; Padhan, N.; Gualandi, L.; Sundvold-Gjerstad, V.; Gustafsson, K.; Choy, W.W.; Zang, G.; et al. VEGFR2 induces c-Src signaling and vascular permeability in vivo via the adaptor protein TSAd. J. Exp. Med. 2012, 209, 1363–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrosa, A.R.; Bodrug, N.; Gomez-Escudero, J.; Carter, E.P.; Reynolds, L.E.; Georgiou, P.N.; Fernandez, I.; Lees, D.M.; Kostourou, V.; Alexopoulou, A.N.; et al. Tumor Angiogenesis Is Differentially Regulated by Phosphorylation of Endothelial Cell Focal Adhesion Kinase Tyrosines-397 and -861. Cancer Res. 2019, 79, 4371–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Wu, L.; Zhou, X.; Zhou, N.; Zhuang, Q.; Yang, J.; Dai, J.; Wang, H.; Chen, S.; Mao, W. Cryptotanshinone inhibits VEGF-induced angiogenesis by targeting the VEGFR2 signaling pathway. Microvasc. Res. 2017, 111, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Toutounchian, J.J.; Pagadala, J.; Miller, D.D.; Baudry, J.; Park, F.; Chaum, E.; Morales-Tirado, V.; Yates, C.R. Novel Small Molecule JP-153 Targets the Src-FAK-Paxillin Signaling Complex to Inhibit VEGF-Induced Retinal Angiogenesis. Mol. Pharmacol. 2017, 91, 1–13. [Google Scholar] [CrossRef]
- Zhao, L.N.; Wang, P.; Liu, Y.H.; Cai, H.; Ma, J.; Liu, L.B.; Xi, Z.; Li, Z.Q.; Liu, X.B.; Xue, Y.X. MiR-383 inhibits proliferation, migration and angiogenesis of glioma-exposed endothelial cells in vitro via VEGF-mediated FAK and Src signaling pathways. Cell Signal. 2017, 30, 142–153. [Google Scholar] [CrossRef]
- Bolos, V.; Gasent, J.M.; Lopez-Tarruella, S.; Grande, E. The dual kinase complex FAK-Src as a promising therapeutic target in cancer. Onco Targets Ther. 2010, 3, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Ashraf-Uz-Zaman, M.; Sajib, M.S.; Cucullo, L.; Mikelis, C.M.; German, N.A. Analogs of penfluridol as chemotherapeutic agents with reduced central nervous system activity. Bioorg. Med. Chem. Lett. 2018, 28, 3652–3657. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.Y.; Lee, C.C.; Chou, Y.M.; Su, C.Y.; Chou, F.H. The incidence and relative risk of stroke in patients with schizophrenia: a five-year follow-up study. Schizophr. Res. 2012, 138, 41–47. [Google Scholar] [CrossRef]
- Chou, F.H.; Tsai, K.Y.; Su, C.Y.; Lee, C.C. The incidence and relative risk factors for developing cancer among patients with schizophrenia: a nine-year follow-up study. Schizophr. Res. 2011, 129, 97–103. [Google Scholar] [CrossRef]
- Barak, Y.; Achiron, A.; Mandel, M.; Mirecki, I.; Aizenberg, D. Reduced cancer incidence among patients with schizophrenia. Cancer 2005, 104, 2817–2821. [Google Scholar] [CrossRef] [PubMed]
- Fond, G.; Macgregor, A.; Attal, J.; Larue, A.; Brittner, M.; Ducasse, D.; Capdevielle, D. Antipsychotic drugs: pro-cancer or anti-cancer? A systematic review. Med. Hypotheses 2012, 79, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.; Soares, R.; Figueiredo-Braga, M.; Coelho, R. Schizophrenia and cancer: is angiogenesis a missed link? Life Sci. 2014, 97, 91–95. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srivastava, S.; Zahra, F.T.; Gupta, N.; Tullar, P.E.; Srivastava, S.K.; Mikelis, C.M. Low Dose of Penfluridol Inhibits VEGF-Induced Angiogenesis. Int. J. Mol. Sci. 2020, 21, 755. https://doi.org/10.3390/ijms21030755
Srivastava S, Zahra FT, Gupta N, Tullar PE, Srivastava SK, Mikelis CM. Low Dose of Penfluridol Inhibits VEGF-Induced Angiogenesis. International Journal of Molecular Sciences. 2020; 21(3):755. https://doi.org/10.3390/ijms21030755
Chicago/Turabian StyleSrivastava, Suyash, Fatema Tuz Zahra, Nehal Gupta, Paul E. Tullar, Sanjay K. Srivastava, and Constantinos M. Mikelis. 2020. "Low Dose of Penfluridol Inhibits VEGF-Induced Angiogenesis" International Journal of Molecular Sciences 21, no. 3: 755. https://doi.org/10.3390/ijms21030755
APA StyleSrivastava, S., Zahra, F. T., Gupta, N., Tullar, P. E., Srivastava, S. K., & Mikelis, C. M. (2020). Low Dose of Penfluridol Inhibits VEGF-Induced Angiogenesis. International Journal of Molecular Sciences, 21(3), 755. https://doi.org/10.3390/ijms21030755