Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
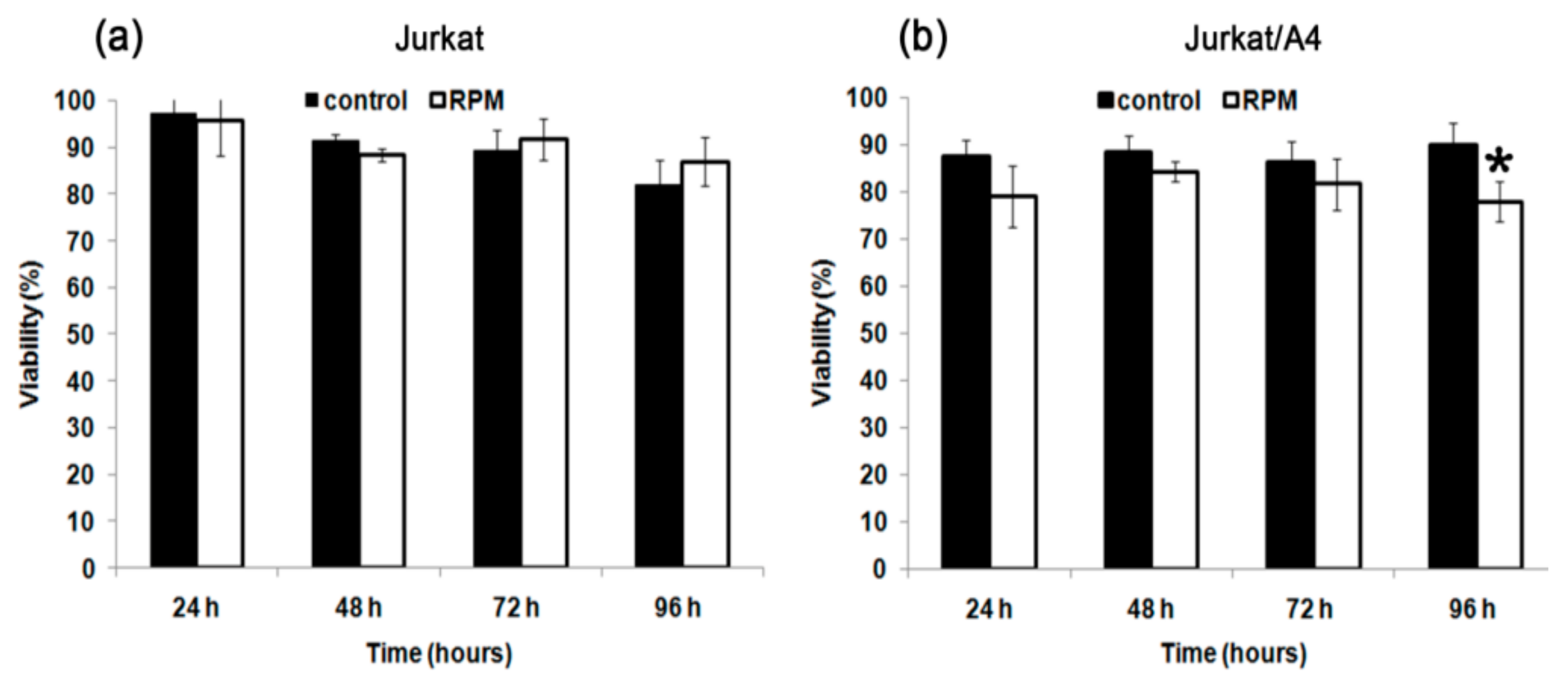
2.1. Effect of Simulated Microgravity on Cell Viability of Jurkat/A4 Cells
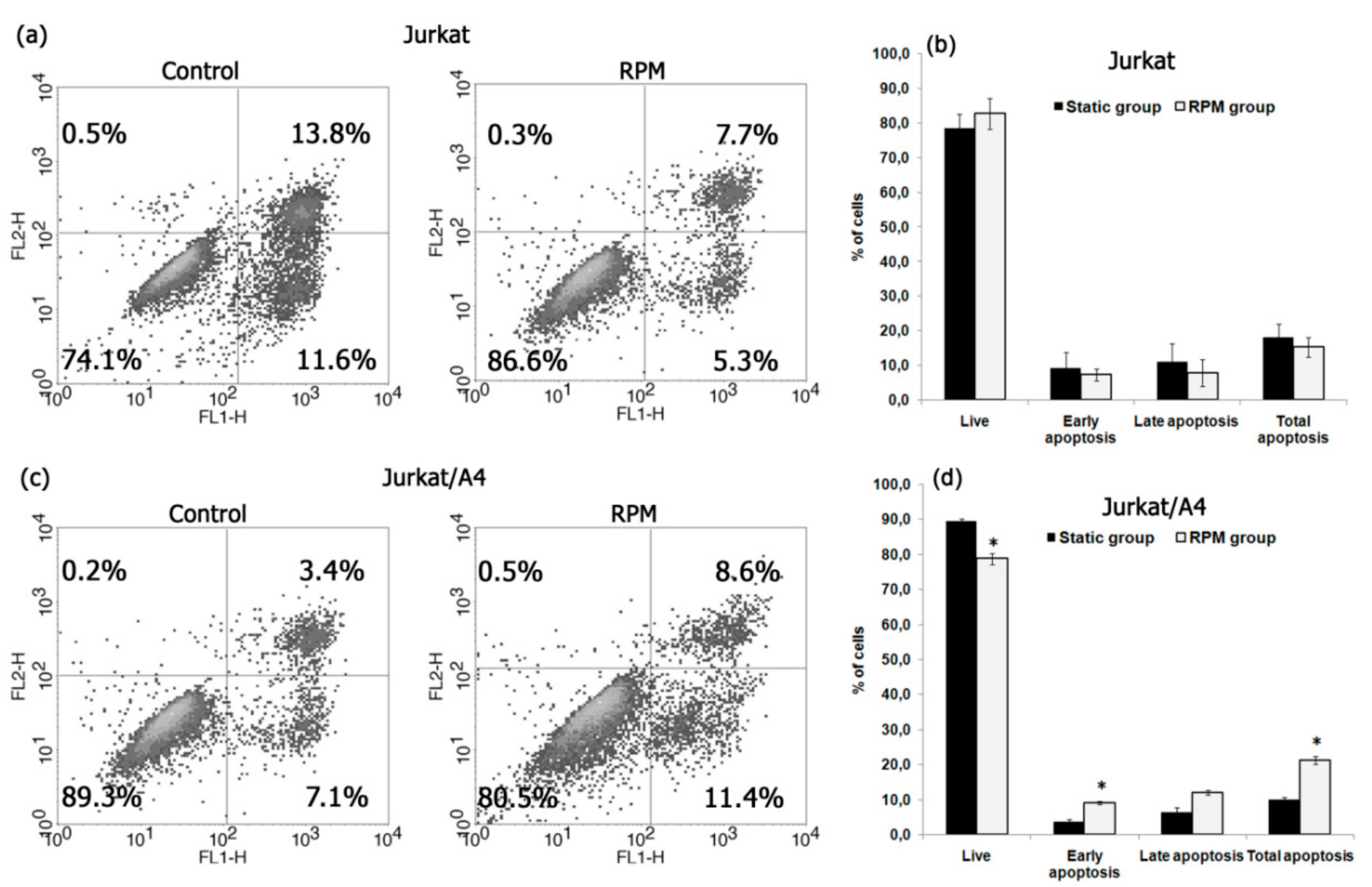
2.2. Simulated Microgravity Induced Apoptosis of Jurkat/A4 Cells
2.3. Simulated Microgravity Disturbed Cell Cycle of Jurkat/A4 Cells
2.4. Simulated Microgravity Changed the Expression of the Surface Marker ICAM-3 in Jurkat/A4 Cells
2.5. The Effect of Simulated Microgravity on the Morphology and Cytoskeleton of Jurkat/A4 Cells
2.6. The Effect of Simulated Microgravity on Certain Apoptotic Proteins of Jurkat/A4 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RPM
4.3. Cell Viability Analysis
4.4. Cell-Cycle Analysis
4.5. Apoptosis Analysis
4.6. Flow Cytometry Immunophenotyping
4.7. Observation of Morphological Changes
4.8. Western Blotting
4.9. Immunocytochemistry
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| Abs | Antibodies |
| BCL2 | B-cell lymphoma 2 |
| BNIP3 | BCL2 interacting protein 3 |
| CD50 | ICAM-3 also known as cluster of differentiation (CD)50 |
| CD95 | Apo-1/Fas (CD95), a receptor of the TNF Receptor (TNFR) family |
| CDDP | cis-diamminedichloridoplatinum(II) |
| Con-A | Concavalin A |
| DAPI | 4′,6-diamidino-2-phenylindole |
| ERK1 | Extracellular signal-regulated kinase1 |
| FAK | Focal adhesion kinase |
| FAS | FS-7-associated surface antigen, also known as apoptosis antigen 1 (APO-1) |
| FBS | Fetal bovine serum |
| FITC | Fluorescein isothiocyanate |
| ICAM-3 | Inter-cellular adhesion molecule 3 |
| LC3 | Microtubule-associated protein light chain 3-II/I ratio |
| MAPK | Mitogen-activated protein kinase |
| MEG-01 | megakaryoblastic cells |
| MFI | Mean fluorescence intensity |
| MCS | Multicellular spheroid |
| mTORC1 | Mammalian target of rapamycin Complex 1 |
| NF-κB | Nuclear factor-kappa B |
| NP-40 | Nonyl Phenoxypolyethoxylethanol-40 |
| PBS | Phosphate-buffered saline |
| PI | Propidium-iodide |
| RhoA | Ras homolog family member A |
| RPM | Random positioning machine |
| RT | Room temperature |
| SDS | Sodium-dodecyl sulfate |
| SDS-PAGE | SDS -polyacrylamide gel electrophoresis |
| TBST | Tris-buffered saline buffer with Tween 20 |
| VCAM1 | Vascular cell adhesion protein 1 |
References
- Pietsch, J.; Bauer, J.; Egli, M.; Infanger, M.; Wise, P.; Ulbrich, C.; Grimm, D. The effects of weightlessness on the human organism and mammalian cells. Curr. Mol. Med. 2011, 11, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, A.I.; Kalinin, Y.T.; Buravkova, L.B.; Mitichkin, O.V. Space cell physiology and space biotechnology in Russia. Adv. Space Biol. Med. 2002, 8, 215–236. [Google Scholar] [PubMed]
- Vernikos, J.; Schneider, V.S. Space, gravity and the physiology of aging: Parallel or convergent disciplines? A mini-review. Gerontology 2010, 56, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, C.; Westphal, K.; Pietsch, J. Characterization of human chondrocytes exposed to simulated microgravity. Cell. Physiol. Biochem. 2010, 25, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M. The Cytoskeleton, apoptosis, and gene expression in T lymphocytes and other mammalian cells exposed to altered gravity. Adv. Space Biol. Med. 2002, 8, 77–128. [Google Scholar]
- Chen, J. Tumor Cells in Microgravity. In Into Space—A Journey of How Humans Adapt and Live in Microgravity; IntechOpen: London, UK, 2018; pp. 259–268. [Google Scholar]
- Uva, B.M. Microgravity-induced apoptosis in cultured glial cells. Eur. J. Histochem. 2002, 46, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Infanger, M. Induction of three dimensional assembly and increase in apoptosis of human endothelial cells by simulated microgravity: Impact of vascular endothelial grow. Apoptosis 2006, 11, 749. [Google Scholar] [CrossRef]
- Infanger, M.; Kossmeh, P.; Shakibaei, M.; Bauer, J.; Kossmehl-Zorn, S.; Cogoli, A.; Curcio, F.; Oksche, A.; Wehland, M.; Kreutz, R.; et al. Simulated weightlessness changes the cytoskeleton and extracellular matrix proteins in papillary thyroid carcinoma cells. Cell Tissue Res. 2006, 324, 267–277. [Google Scholar] [CrossRef]
- Grimm, D.; Bauer, J.; Kossmehl, P.; Shakibaei, M.; Schönberger, J. Simulated microgravity alters differentiation and increases apoptosis in human follicular thyroid carcinoma cells. FASEB J. 2002, 16, 604–606. [Google Scholar] [CrossRef]
- Grimm, D.; Bauer, J.; Ulbrich, C.; Westphal, K.; Wehland, M. Different responsiveness of endothelial cells to vascular endothelial growth factor and basic fibroblast growth factor added to culture media under gravity and simulated microgravity. Tissue Eng. Part A 2010, 16, 1559–1573. [Google Scholar] [CrossRef]
- Uva, B.M.; Masini, M.A.; Sturla, M.; Prato, P.; Passalacqua, M. Clinorotation-induced weightlessness influences the cytoskeleton of glial cells in culture. Brain Res. 2002, 934, 132–139. [Google Scholar] [CrossRef]
- Li, J.; Zhang, S.; Chen, J.; Du, T.Y.; Wang, Y.C.; Wang, Z.R. Modeled microgravity causes changes in the cytoskeleton and focal adhesions, and decreases in migration in malignant human MCF-7 cells. Protoplasma 2009, 238, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Borst, A.G.; van Loon, J.J.W.A. Technology and development for the random positioning machine, RPM. Microgravity Sci. Technol. 2009, 21, 1161–1165. [Google Scholar] [CrossRef]
- Hauschild, S.; Tauber, S.; Lauber, B.; Layer, L.E.; Ullrich, O. T cell regulation in microgravity—The current knowledge from in vitro experiments conducted in space, parabolic flights and ground-based facilities. Acta Astronaut. 2014, 104, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Morabito, C.; Lanuti, P.; Caprara, G.A.; Marchisio, M.; Bizzarri, M.; Guarnieri, S.; Mariggiò, M.A. Physiological Responses of Jurkat Lymphocytes to Simulated Microgravity Condition. Int. J. Mol. Sci. 2019, 20, 1892. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight alters microtubules and increases apoptosis in human lymphocytes (Jurkat). FASEB J. 1998, 12, 1007–1018. [Google Scholar] [CrossRef]
- Ullrich, O.; Thiel, C.S. Gravitational force: Triggered stress in cells of the immune system. In Stress Challenges, and Immunity in Space: From Mechanisms to Monitoring and Preventive Strategies; Chouk’er, A., Ed.; Springer: Berlin, Germany, 2012; pp. 187–202. [Google Scholar]
- Krüger, M.; Melnik, D.; Kopp, S.; Buken, C.; Sahana, J.; Bauer, J.; Wehland, M.; Hemmersbach, R.; Corydon, T.J.; Infanger, M.; et al. Fighting Thyroid Cancer with Microgravity Research. Int. J. Mol. Sci. 2019, 20, 2553. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, T.; Tanimoto, K.; Shrestha, L.; Imura, T.; Takahashi, S.; Sueda, T.; Hirohashi, N.; Hiyama, E.; Yuge, L. Simulated microgravity enhances CDDP-induced apoptosis signal via p53-independent mechanisms in cancer cells. PLoS ONE 2019, 14, e0219363. [Google Scholar] [CrossRef] [Green Version]
- Aleshcheva, G.; Bauer, J.; Hemmersbach, R.; Slumstrup, L.; Wehland, M.; Infanger, M.; Grimm, D. Scafold-free tissue formation under real and simulated microgravity conditions. Basic Clin. Pharmacol. Toxicol. 2016, 119, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Prodanov, L.; Van Loon, J.J.; Te Riet, J.; Jansen, J.A.; Walboomers, X.F. Nanostructured substrate conformation can decrease osteoblastlike cell dysfunction in simulated microgravity conditions. J. Tissue Eng. Regen. Med. 2014, 8, 978–988. [Google Scholar] [CrossRef]
- Riwaldt, S.; Pietsch, J.; Sickmann, A. Identification of proteins involved in inhibition of spheroid formation under microgravity. Proteomics 2015, 15, 2945–2952. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Q.; Gastman, B.R.; Wieckowski, E.; Goldstein, L.A.; Rabinovitz, A.; Yin, X.-M.; Rabinowich, H. Apoptosis-resistant mitochondria in T cells selected for resistance to Fas signaling. J. Biol. Chem. 2001, 276, 3610–3619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigorito, E.; Plaza, S.; Mir, L.; Mongay, L.; Vinas, O.; Serra-Pages, C.; Vives, J. Contributions of p53 and PMA to irradiation induced apoptosis in Jurkat cells. Hematol. Cell 1999, 41, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Sokolovskaya, A.A.; Zabotina, T.N.; Blokhin, D.Yu.; Kadagidze, Z.G.; Baryshnikov, A.Yu. CD95-deficient cells of Jurkat/A4 subline are resistant to drug-induced apoptosis. Exp. Oncol. 2001, 23, 175–180. [Google Scholar]
- Mikhailov, A.; Sokolovskaya, A.A.; Yegutkin, G.G.; Amdahl, H.; West, A.; Yagita, H.; Lahesmaa, R.; Thompson, L.F.; Jalkanen, S.; Blokhin, D.; et al. CD73 participates in cellular multiresistance program and protects against TRAIL-induced apoptosis. J. Immunol. 2008, 181, 464–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakely, W.F.; Miller, A.C.; Grace, M.B.; McLeland, C.B.; Luo, L.; Muderhwa, J.M.; Miner, V.L.; Prasanna, P.G. Radiation biodosimetry: Applications for spaceflight. Adv. Space Res. 2003, 31, 1487–1493. [Google Scholar] [CrossRef]
- Thiel, C.S.; Huge, A.; Hauschild, S.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B. Stability of gene expression in human T cells in different gravity environments is clustered in chromosomal region 11p15.4. Npj Microgravity. 2017, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Buken, C.; Sahana, J.; Corydon, T.J.; Melnik, D.; Bauer, J.; Wehland, M.; Kruger, M.; Balk, S.; Abuagela, N.; Infanger, M.; et al. Morphological and Molecular Changes in Juvenile Normal Human Fibroblasts Exposed to Simulated Microgravity. Sci. Rep. 2019, 9, 11882. [Google Scholar] [CrossRef]
- Nassef, M.Z.; Kopp, S.; Wehland, M.; Melnik, D.; Sahana, J.; Kruger, M.; Corydon, T.J.; Oltmann, H.; Schmitz, B.; Schutte, A.; et al. Real Microgravity Influences the Cytoskeleton and Focal Adhesions in Human Breast Cancer Cells. Int. J. Mol. Sci. 2019, 20, 3156. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, A.; Grimm, D.; Sahana, J.; Bauer, J.; Kruger, M.; Infanger, M.; Magnusson, N.E. Key Proteins Involved in Spheroid Formation and Angiogenesis in Endothelial Cells After Long-Term Exposure to stimulated Microgravity. Cell Physiol. Biochem. 2018, 45, 429–445. [Google Scholar] [CrossRef]
- Coinu, R.; Chiaviello, A.; Galleri, G.; Franconi, F.; Crescenzi, E.; Palumbo, G. Exposure to modeled microgravity induces metabolic idleness in malignant human MCF-7 and normal murine VSMC cells. FEBS Lett. 2006, 580, 2465–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, M.; Magakia, T.; Okazakia, T.; Kawahara, Y.; Manabe, T.; Yuge, L.; Kurisu, K. Effects of simulated microgravity on proliferation and chemosensitivity in malignant glioma cells. Neurosci. Lett. 2009, 463, 54–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolovskaya, A.A.; Korneeva, E.A.; Kolesov, D.V.; Moskovtsev, A.A.; Kubatiev, A.A. Inhibition of cell cycle progression and changes in surface markers in MEG-01 megakaryoblastic cells exposed to the random positioning machine. Microgravity Sci. Technol. 2019. [Google Scholar] [CrossRef]
- Paulsen, K.; Tauber, S.; Dumrese, C.; Bradacs, G.; Simmet, D.M.; Golz, N.; Hauschild, S.; Raig, C.; Engeli, S.; Gutewort, A.; et al. Regulation of ICAM-1 in cells of the monocyte/macrophage system in microgravity. Biomed. Res. Int. 2015, 538786. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Yu.; Sang, C.; Paulsen, K.; Arenz, A.; Zhao, Z.; Jia, X.; Ullrich, O.; Zhuang, F. ICAM-1 expression and organization in human endothelial cells is sensitive to gravity. Acta Astronautica 2010, 67, 1073–1080. [Google Scholar] [CrossRef]
- Buravkova, L.; Romanov, Y.; Rykova, M.; Grigorieva, O.; Merzlikina, N. “Cell-to-cell interactions in changed gravity: Ground-based and flight experiments”. Acta Astronautica 2005, 57, 67–74. [Google Scholar] [CrossRef]
- Buravkova, L.B.; Rudimov, E.G.; Andreeva, E.R.; Grigoriev, A. The ICAM-1 expression level determines the susceptibility of human endothelial cells to simulated microgravity. J. Cell. Biochem. 2018, 119, 2875–2885. [Google Scholar] [CrossRef]
- Moffatt, O.D.; Devitt, A.; Bell, E.D.; Simmons, D.L.; Gregory, C.D. Macrophage recognition of ICAM-3 on apoptotic leukocytes. J. Immunol. 1999, 162, 6800–6810. [Google Scholar]
- Torr, E.E.; Gardner, D.H.; Thomas, L.; Goodall, D.M.; Bielemeier, A.; Willetts, R.; Griffiths, H.R.; Marshall, L.J.; Devitt, A. Apoptotic cell-derived ICAM-3 promotes both macrophage chemoattraction to and tethering of apoptotic cells. Cell Death Differ. 2012, 19, 671–679. [Google Scholar] [CrossRef]
- Aleshcheva, G.; Wehland, M.; Sahana Bauer, J.; Corydon, T.J.; Hemmersbach, R.; Frett, T.; Egli, M.; Infanger, M.; Grosse, J. Moderate alterations of the cytoskeleton in human chondrocytes after short-term microgravity produced by parabolic flight maneuvers could be prevented by up-regulation of bmp-2 and sox-9. FASEB J. 2015, 29, 2303–2314. [Google Scholar] [CrossRef]
- Bizzarri, M.; Masiello, M.G.; Giuliani, A.; Cucina, A. Gravity constraints drive biological systems toward specific organization patterns: Commitment of cell specification is constrained by physical cues. Bioessays 2018, 40, 1700138. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Tang, X.; Umeshappa, C.S.; Ma, H.; Gao, H.; Deng, Y.; Freywald, A.; Xiang, J. Simulated Microgravity Promotes Cell Apoptosis Through Suppressing Uev1A/TICAM/TRAF/NF-κB-Regulated Anti-Apoptosis and p53/PCNA- and ATM/ATR-Chk1/2-Controlled DNA-Damage Response Pathways. J. Cell Biochem. 2016, 117, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, T.; Biggi, F.; Bassi, A.M.; Ferrando, S.; Gallus, L.; Loiacono, F.; Ravera, S.; Rottigni, M.; Scarfì, S.; Strollo, F. Simulated microgravity induces nuclear translocation of Bax and BCL-2 in glial cultured C6 cells. Heliyon 2019, 5, e01798. [Google Scholar] [CrossRef] [Green Version]
- Cavalcanti, G.B., Jr.; Scheiner, M.A.M.; Simões Magluta, E.P.; Vasconcelos, F.C.; Klumb, C.E.; Maia, R.C. p53 flow cytometry evaluation in leukemias: Correlation to factors affecting clinical outcome. Cytom. Part B 2010, 78, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Guadamillas, M.C.; Cerezo, A.; del Pozo, M.A. Overcoming anoikis—Pathways to anchorage in dependent growth in cancer. J. Cell Sci. 2011, 124, 3189–3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhwani, V.J.; Kale, R.K.; Singh, R.P. Microgravity Alters Cancer Growth and Progression. Curr. Cancer Drug Targets. 2014, 14, 394–406. [Google Scholar]
- Albi, E.; Krüger, M.; Hemmersbach, R.; Lazzarini, A.; Cataldi, S.; Codini, M.; Beccari, T.; Ambesi Impiombato, F.S.; Curcio, F. Impact of gravity on thyroid cells. Int. J. Mol. Sci. 2017, 18, 972. [Google Scholar] [CrossRef] [Green Version]
- Braddock, M. From Target identification to Drug Development in Space: Using the Microgravity Assist. Curr. Drug Discov. Technol. 2019, 16. [Google Scholar] [CrossRef]
- Zanoni, M.; Piccinini, F.; Arienti, C.; Zamagni, A.; Santi, S.; Polico, R.; Bevilacqua, A.; Tesei, A. 3D tumor spheroid models for in vitro therapeutic screening: A systematic approach to enhance the biological relevance of data obtained. Sci. Rep. 2016, 6, 19103. [Google Scholar] [CrossRef]
- Yeung, C.K.; Koenig, P.; Countryman, S.; Thummel, K.E.; Himmelfarb, J.; Kelly, E.J. Tissue Chips in Space—Challenges and Opportunities. Clin. Transl. Sci. 2020, 13, 8–10. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokolovskaya, A.; Korneeva, E.; Zaichenko, D.; Virus, E.; Kolesov, D.; Moskovtsev, A.; Kubatiev, A. Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine. Int. J. Mol. Sci. 2020, 21, 855. https://doi.org/10.3390/ijms21030855
Sokolovskaya A, Korneeva E, Zaichenko D, Virus E, Kolesov D, Moskovtsev A, Kubatiev A. Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine. International Journal of Molecular Sciences. 2020; 21(3):855. https://doi.org/10.3390/ijms21030855
Chicago/Turabian StyleSokolovskaya, Alisa, Ekaterina Korneeva, Danila Zaichenko, Edward Virus, Dmitry Kolesov, Aleksey Moskovtsev, and Aslan Kubatiev. 2020. "Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine" International Journal of Molecular Sciences 21, no. 3: 855. https://doi.org/10.3390/ijms21030855
APA StyleSokolovskaya, A., Korneeva, E., Zaichenko, D., Virus, E., Kolesov, D., Moskovtsev, A., & Kubatiev, A. (2020). Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine. International Journal of Molecular Sciences, 21(3), 855. https://doi.org/10.3390/ijms21030855