Reduced miR-26b Expression in Megakaryocytes and Platelets Contributes to Elevated Level of Platelet Activation Status in Sepsis

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
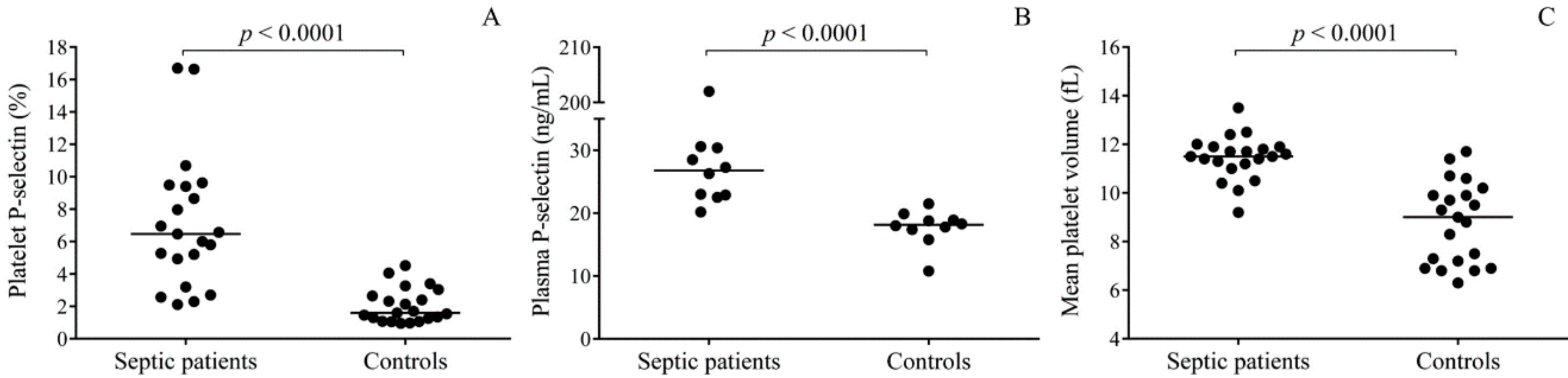
2.1. Baseline Characteristics of Study Groups
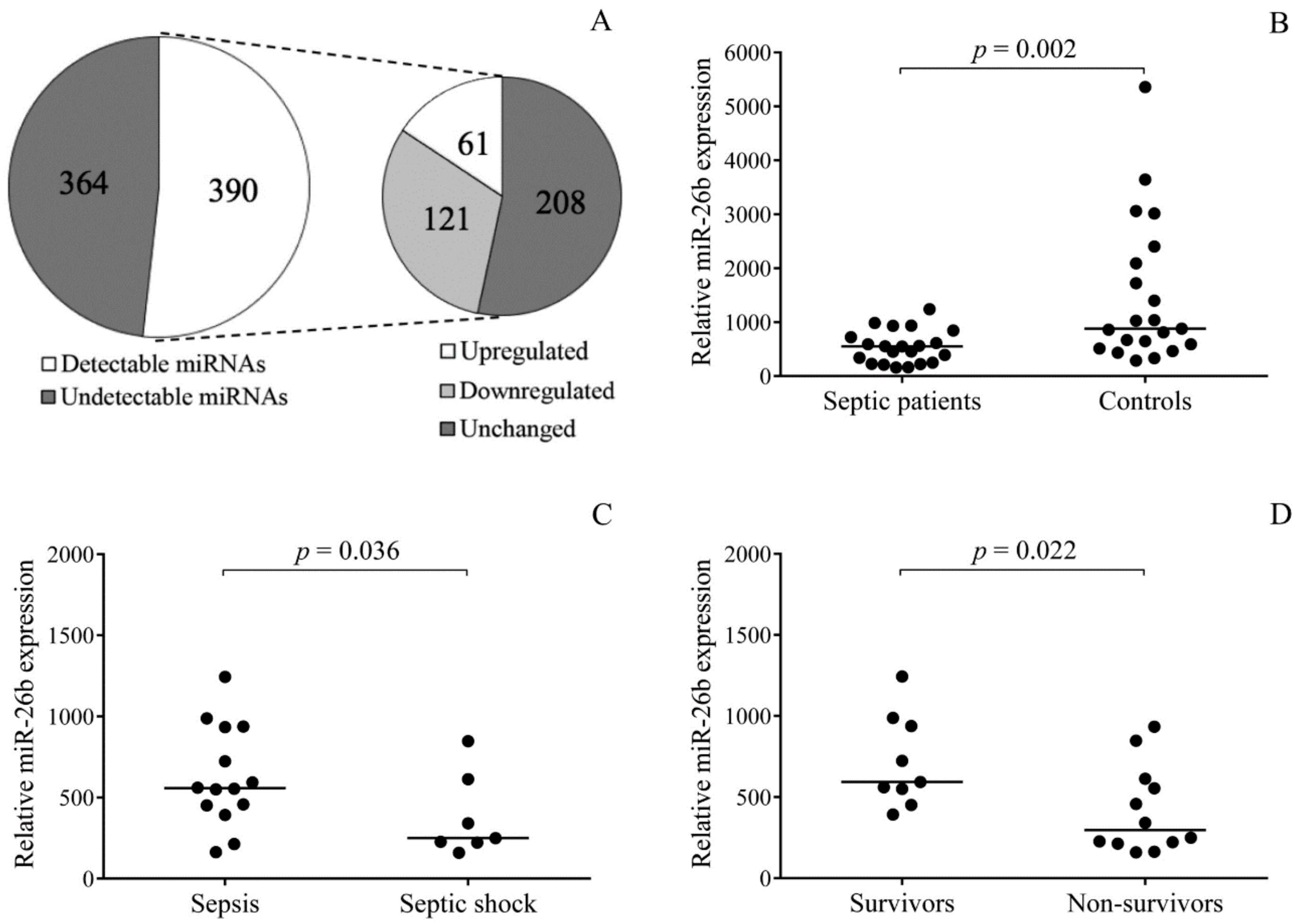
2.2. Sepsis-Induced Changes in Platelet miRNAs
2.3. Platelet miR-26b Expression Correlates with Sepsis Severity and Mortality
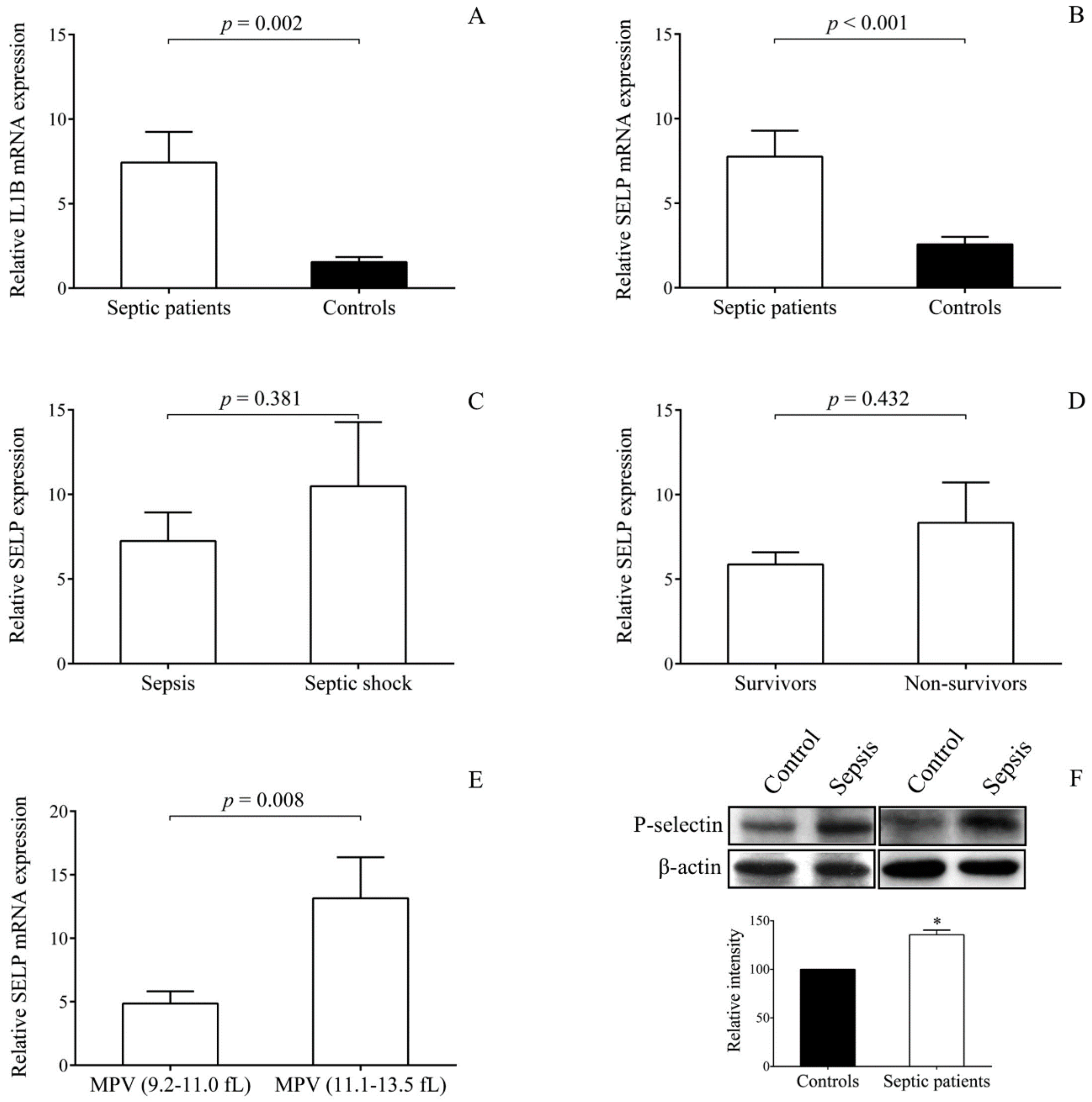
2.4. SELP Expression is Upregulated in Platelets with Sepsis
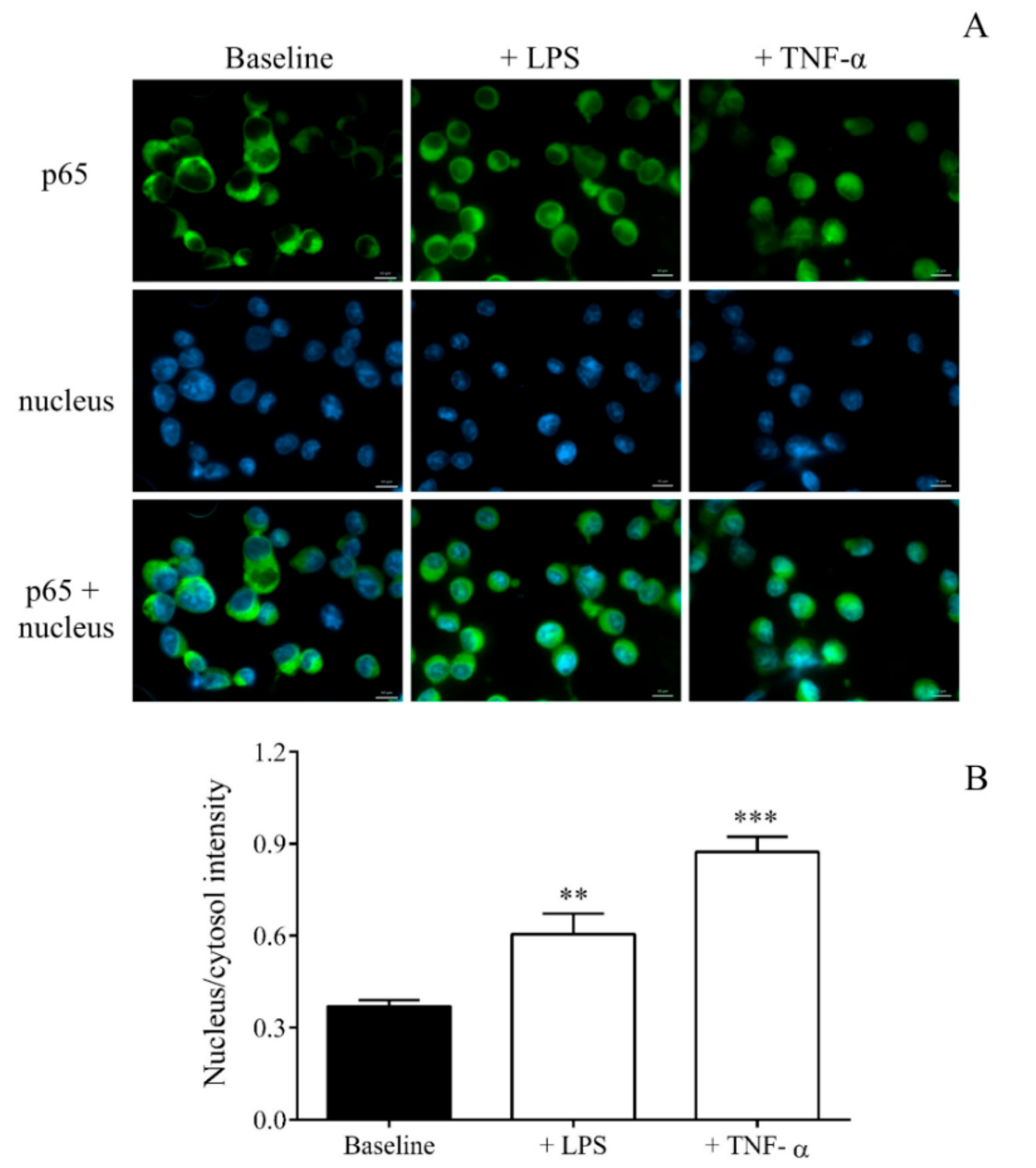
2.5. TLR4 Modulated Transcriptome in MEG-01 Cells Via Nuclear Factor-Kappa B (NF-κB) Pathway
2.6. Sepsis-Reduced miR-26b Results in Upregulated SELP Expression
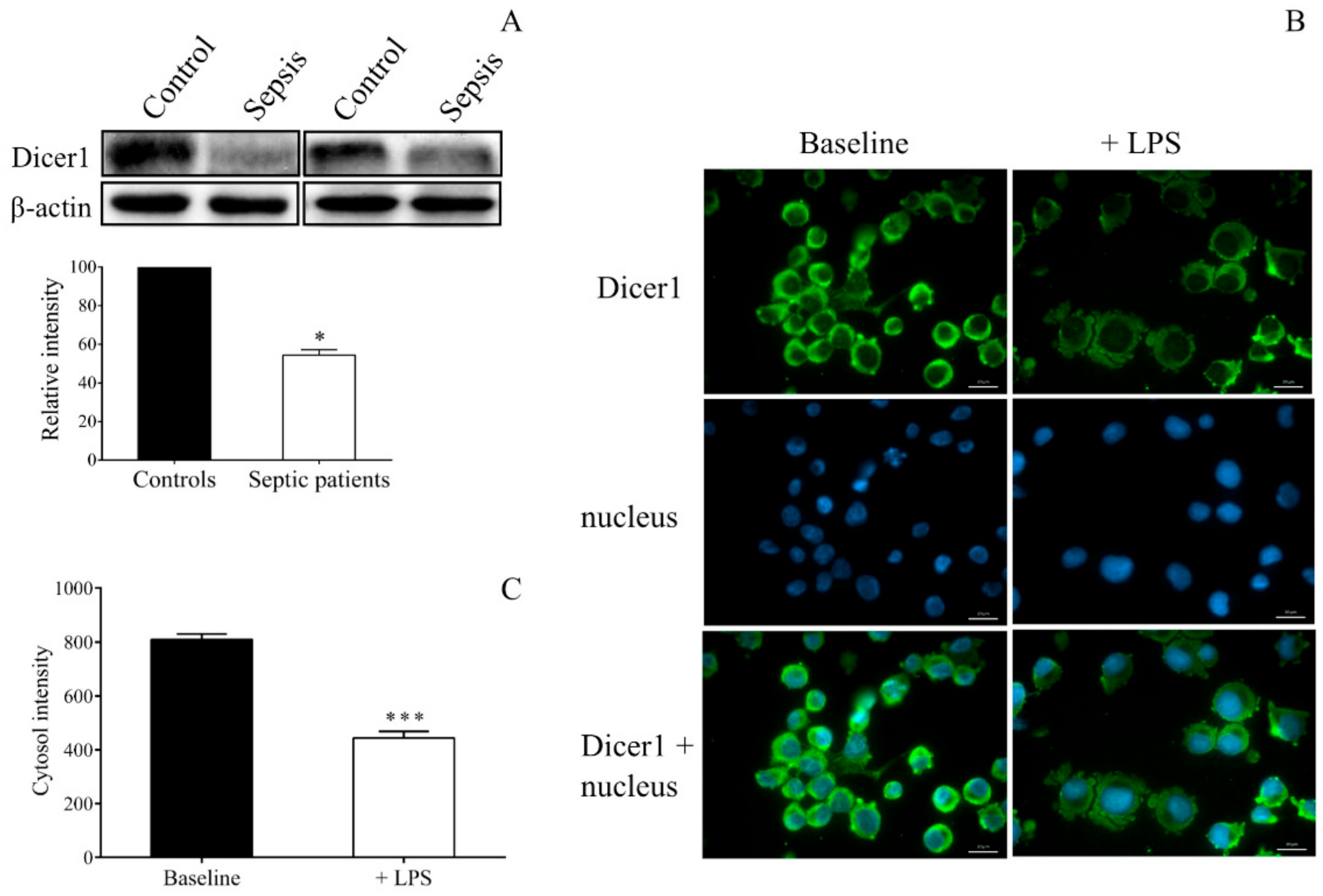
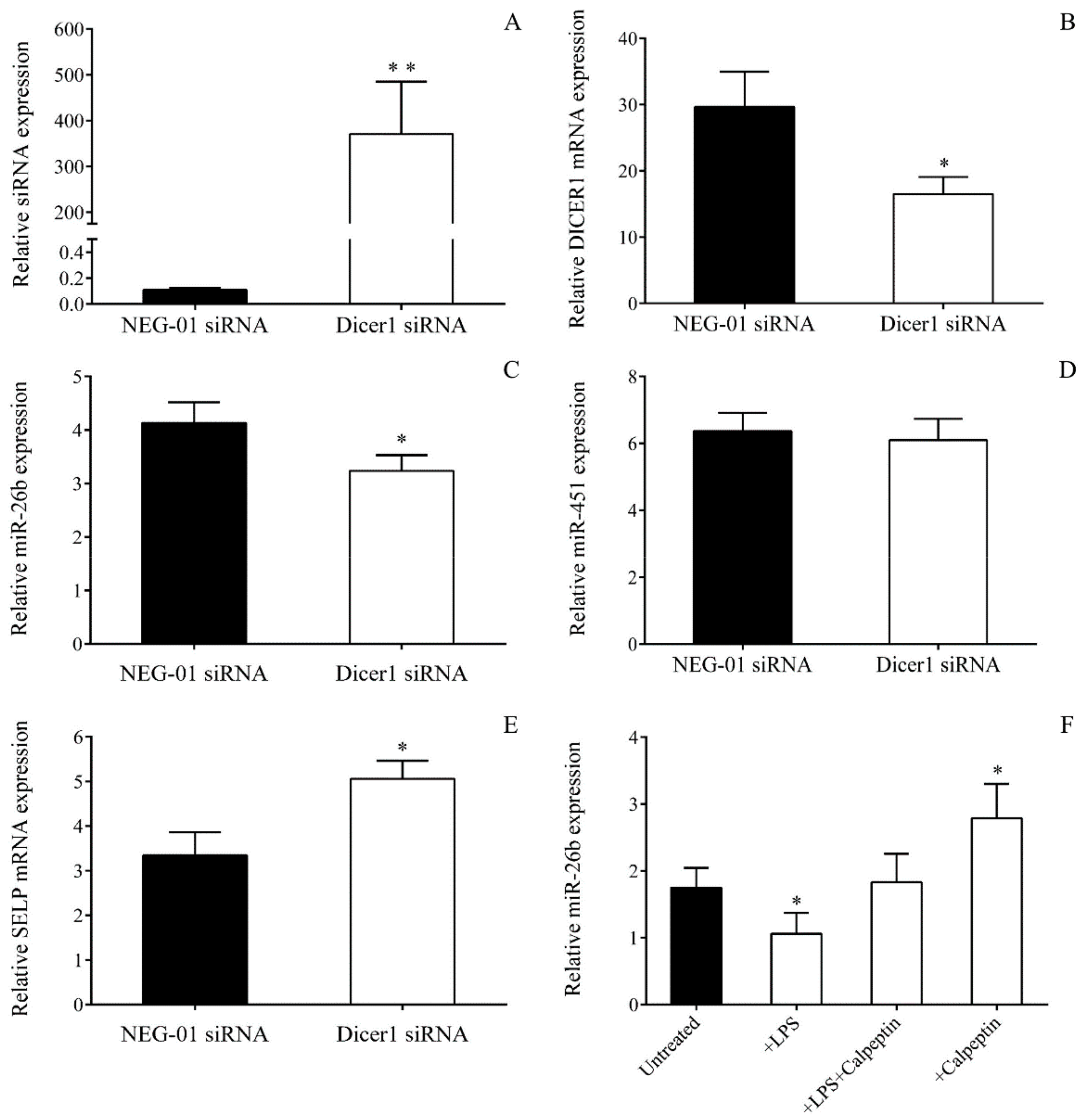
2.7. Decreased Dicer1 Level in Platelets and MKs Results in Abnormal miRNA Among Septic Conditions
3. Discussion
4. Materials and Methods
4.1. Study Design, Participants, and Blood Sample Preparation
4.2. Leukocyte-Depleted Platelet Preparation
4.3. Total RNA Extraction
4.4. TaqMan Open Array-Based miRNA Profiling in Septic Platelets
4.5. miRNA Specific Stem-Loop RT-qPCR Analysis
4.6. mRNA Specific RT-qPCR Analysis
4.7. In Vitro Activation of Normal Human Platelets by LPS
4.8. Culturing of MEG-01 Cells Mimicking Septic Conditions
4.9. RNA-Sequencing
4.10. Analysis of RNA-Seq Data
4.11. Gene Ontology Analysis
4.12. Transfection of MEG-01 Cells with miR-26b Mimic
4.13. Western Blot
4.14. Flow Cytometry
4.15. Fluorescence Microscopy
4.16. Downregulation of DICER1 Expression by siRNA Transfection in MEG-01 Cells
4.17. Analysis of Dicer Function on miRNA Level Through Calpain Inhibition in MEG-01 Cells Among Inflammatory Conditions
4.18. Other Laboratory Assays
4.19. Data Presentation and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MKs | Megakaryocytes |
| miRNA | MicroRNA |
| mRNA | Messenger RNA |
| TLRs | Toll-like receptors |
| IL-1β | Interleukin-1β |
| LPS | Lipopolysaccharide |
| GO | Gene ontology |
| ICU | Intensive care unit |
| RT | Room temperature |
| PRP | Platelet-rich plasma |
| LDP | Leukocyte-depleted platelet |
| UPL | Universal ProbeLibrary |
| cDNA | Complementary DNA |
| FBS | Fetal bovine serum |
| LBP | Lipoprotein binding protein |
| TNF-α | Tumor necrosis factor-α |
| NF-κB | Nuclear factor-kappa B |
| WBC | White blood cell |
| MPV | Mean platelet volume |
| CRP | C-reactive protein |
| PCT | Procalcitonin |
| DE | Differentially expressed |
| PLT | platelet |
| MAPK | mitogen-activated protein kinase |
| ERK1/2 | extracellular signal-regulated kinase 1/2 |
| JNK | c-Jun N-terminal kinase |
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Vardon-Bounes, F.; Ruiz, S.; Gratacap, M.P.; Garcia, C.; Payrastre, B.; Minville, V. Platelets Are Critical Key Players in Sepsis. Int. J. Mol. Sci. 2019, 20, 3494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russwurm, S.; Vickers, J.; Meier-Hellmann, A.; Spangenberg, P.; Bredle, D.; Reinhart, K.; Lösche, W. Platelet and leukocyte activation correlate with the severity of septic organ dysfunction. Shock 2002, 17, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Gawaz, M.; Dickfeld, T.; Bogner, C.; Fateh-Moghadam, S.; Neumann, F.J. Platelet function in septic multiple organ dysfunction syndrome. Intensive Care Med. 1997, 23, 379–385. [Google Scholar] [CrossRef]
- Akinosoglou, K.; Theodoraki, S.; Xanthopoulou, I.; Perperis, A.; Gkavogianni, T.; Pistiki, A.; Giamarellos-Bourboulis, E.; Gogos, C.A. Platelet reactivity in sepsis syndrome: Results from the PRESS study. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2503–2512. [Google Scholar] [CrossRef]
- Yaguchi, A.; Lobo, F.L.; Vincent, J.L.; Pradier, O. Platelet function in sepsis. J. Thromb. Haemost. 2004, 2, 2096–2102. [Google Scholar] [CrossRef]
- Adamzik, M.; Görlinger, K.; Peters, J.; Hartmann, M. Whole blood impedance aggregometry as a biomarker for the diagnosis and prognosis of severe sepsis. Crit Care 2012, 16, R204. [Google Scholar] [CrossRef] [Green Version]
- Assinger, A.; Schrottmaier, W.C.; Salzmann, M.; Rayes, J. Platelets in Sepsis: An Update on Experimental Models and Clinical Data. Front. Immunol. 2019, 10, 1687. [Google Scholar] [CrossRef]
- Kappelmayer, J.; Nagy, B., Jr.; Miszti-Blasius, K.; Hevessy, Z.; Setiadi, H. The emerging value of P-selectin as a disease marker. Clin. Chem. Lab. Med. 2004, 42, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Rondina, M.T.; Carlisle, M.; Fraughton, T.; Brown, S.M.; Miller, R.R., 3rd; Harris, E.S.; Weyrich, A.S.; Zimmerman, G.A.; Supiano, M.A.; Grissom, C.K. Platelet-monocyte aggregate formation and mortality risk in older patients with severe sepsis and septic shock. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Mosad, E.; Elsayh, K.I.; Eltayeb, A.A. Tissue factor pathway inhibitor and P-selectin as markers of sepsis-induced non-overt disseminated intravascular coagulopathy. Clin. Appl. Thromb. Hemost. 2011, 17, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Andonegui, G.; Kerfoot, S.M.; McNagny, K.; Ebbert, K.V.; Patel, K.D.; Kubes, P. Platelets express functional Toll-like receptor-4. Blood 2005, 106, 2417–2423. [Google Scholar] [CrossRef] [PubMed]
- Schattner, M. Platelet TLR4 at the crossroads of thrombosis and the innate immune response. J. Leukoc. Biol. 2019, 105, 873–880. [Google Scholar] [CrossRef]
- Beaulieu, L.M.; Lin, E.; Morin, K.M.; Tanriverdi, K.; Freedman, J.E. Regulatory effects of TLR2 on megakaryocytic cell function. Blood 2011, 117, 5963–5974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landry, P.; Plante, I.; Ouellet, D.L.; Perron, M.P.; Rousseau, G.; Provost, P. Existence of a microRNA pathway in anucleate platelets. Nat. Struct. Mol. Biol. 2009, 16, 961–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rondina, M.T.; Weyrich, A.S. Regulation of the genetic code in megakaryocytes and platelets. J. Thromb. Haemost. 2015, 13 Suppl. 1, S26–S32. [Google Scholar] [CrossRef]
- Nagalla, S.; Shaw, C.; Kong, X.; Kondkar, A.A.; Edelstein, L.C.; Ma, L.; Chen, J.; McKnight, G.S.; López, J.A.; Yang, L.; et al. Platelet microRNA-mRNA coexpression profiles correlate with platelet reactivity. Blood 2011, 117, 5189–5197. [Google Scholar] [CrossRef]
- Rowley, J.W.; Chappaz, S.; Corduan, A.; Chong, M.M.; Campbell, R.; Khoury, A.; Manne, B.K.; Wurtzel, J.G.; Michael, J.V.; Goldfinger, L.E.; et al. Dicer1-mediated miRNA processing shapes the mRNA profile and function of murine platelets. Blood 2016, 127, 1743–1751. [Google Scholar] [CrossRef] [Green Version]
- Miao, X.; Rahman, M.F.; Jiang, L.; Min, Y.; Tan, S.; Xie, H.; Lee, L.; Wang, M.; Malmström, R.E.; Lui, W.O.; et al. Thrombin-reduced miR-27b attenuates platelet angiogenic activities in vitro via enhancing platelet synthesis of anti-angiogenic thrombospondin-1. J. Thromb. Haemost. 2018, 16, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Shashkin, P.N.; Brown, G.T.; Ghosh, A.; Marathe, G.K.; McIntyre, T.M. Lipopolysaccharide is a direct agonist for platelet RNA splicing. J. Immunol. 2008, 181, 3495–3502. [Google Scholar] [CrossRef] [Green Version]
- Rondina, M.T.; Schwertz, H.; Harris, E.S.; Kraemer, B.F.; Campbell, R.A.; Mackman, N.; Grissom, C.K.; Weyrich, A.S.; Zimmerman, G.A. The septic milieu triggers expression of spliced tissue factor mRNA in human platelets. J. Thromb. Haemost. 2011, 9, 748–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, E.A.; Rowley, J.W.; Campbell, R.A.; Grissom, C.K.; Brown, S.M.; Beesley, S.J.; Schwertz, H.O.R.; Kosaka, Y.; Manne, B.K.; Krauel, K.; et al. Sepsis Alters the Transcriptional and Translational Landscape of Human and Murine Platelets. Blood 2019, 134, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Zampetaki, A.; Dudek, K.; Kaudewitz, D.; King, A.; Kirkby, N.S.; Crosby-Nwaobi, R.; Prokopi, M.; Drozdov, I.; Langley, S.R.; et al. Circulating microRNAs as novel biomarkers for platelet activation. Circ. Res. 2013, 112, 595–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.F.; Kristensen, S.D.; Mathur, A.; Grove, E.L.; Choudry, F.A. The causal role of megakaryocyte–platelet hyperactivity in acute coronary syndromes. Nat. Rev. Cardiol. 2012, 9, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, L.C.; McKenzie, S.E.; Shaw, C.; Holinstat, M.A.; Kunapuli, S.P.; Bray, P.F. MicroRNAs in platelet production and activation. J. Thromb. Haemost. 2013, 11 Suppl. 1, 340–350. [Google Scholar] [CrossRef]
- Fejes, Z.; Póliska, S.; Czimmerer, Z.; Káplár, M.; Penyige, A.; Gál Szabó, G.; Beke Debreceni, I.; Kunapuli, S.P.; Kappelmayer, J.; Nagy, B., Jr. Hyperglycaemia suppresses microRNA expression in platelets to increase P2RY12 and SELP levels in type 2 diabetes mellitus. Thromb. Haemost. 2017, 117, 529–542. [Google Scholar] [CrossRef]
- Beaulieu, L.M.; Freedman, J.E. The role of inflammation in regulating platelet production and function: Toll-like receptors in platelets and megakaryocytes. Thromb. Res. 2010, 125, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Randriamboavonjy, V.; Fleming, I. All cut up! The consequences of calpain activation on platelet function. Vascul. Pharmacol. 2012, 56, 210–215. [Google Scholar] [CrossRef]
- Montrucchio, G.; Bosco, O.; Del Sorbo, L.; Fascio Pecetto, P.; Lupia, E.; Goffi, A.; Omedè, P.; Emanuelli, G.; Camussi, G. Mechanisms of the priming effect of low doses of lipopoly-saccharides on leukocyte dependent platelet aggregation in whole blood. Thromb. Haemost. 2003, 90, 872–881. [Google Scholar] [CrossRef]
- Kappelmayer, J.; Beke Debreceni, I.; Vida, A.; Antal-Szalmás, P.; Clemetson, K.J.; Nagy, B., Jr. Distinct effects of Re- and S-forms of LPS on modulating platelet activation. J. Thromb. Haemost. 2013, 11, 775–778. [Google Scholar] [CrossRef]
- Jayachandran, M.; Brunn, G.J.; Karnicki, K.; Miller, R.S.; Owen, W.G.; Miller, V.M. In vivo effects of lipopolysaccharide and TLR4 on platelet production and activity: Implications for thrombotic risk. J. Appl. Physiol. (1985) 2007, 102, 429–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayachandran, M.; Miller, V.M.; Brunn, G.J.; Owen, W.G. Platelet response as a sentinel marker of toll-like receptor 4 activation in mice. Thromb. Res. 2010, 126, 414–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, A.; Dunoyer-Geindre, S.; Zapilko, V.; Nolli, S.; Reny, J.L.; Fontana, P. Functional Validation of microRNA-126-3p as a Platelet Reactivity Regulator Using Human Haematopoietic Stem Cells. Thromb. Haemost. 2019, 119, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, I.; Bhatlekar, S.; Manne, B.K.; Stoller, M.; Hugo, S.; Kong, X.; Ma, L.; Rondina, M.T.; Weyrich, A.S.; Edelstein, L.C.; et al. miR-15a-5p regulates expression of multiple proteins in the megakaryocyte GPVI signaling pathway. J. Thromb. Haemost. 2019, 17, 511–524. [Google Scholar] [CrossRef]
- Reithmair, M.; Buschmann, D.; Märte, M.; Kirchner, B.; Hagl, D.; Kaufmann, I.; Pfob, M.; Chouker, A.; Steinlein, O.K.; Pfaffl, M.W.; et al. Cellular and extracellular miRNAs are blood-compartment-specific diagnostic targets in sepsis. J. Cell Mol. Med. 2017, 21, 2403–2411. [Google Scholar] [CrossRef]
- Plé, H.; Landry, P.; Benham, A.; Coarfa, C.; Gunaratne, P.H.; Provost, P. The repertoire and features of human platelet microRNAs. PLoS ONE 2012, 7, e50746. [Google Scholar] [CrossRef]
- Sunderland, N.; Skroblin, P.; Barwari, T.; Huntley, R.P.; Lu, R.; Joshi, A.; Lovering, R.C.; Mayr, M. MicroRNA Biomarkers and Platelet Reactivity: The Clot Thickens. Circ. Res. 2017, 120, 418–435. [Google Scholar] [CrossRef] [Green Version]
- Szilágyi, B.; Fejes, Z.; Pócsi, M.; Kappelmayer, J.; Nagy, B., Jr. Role of sepsis modulated circulating microRNAs. EJIFCC 2019, 30, 128–145. [Google Scholar]
- Harrison, P.; Goodall, A.H. “Message in the platelet”—More than just vestigial mRNA! Platelets 2008, 19, 395–404. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, M.; Han, S.; Hwang, J.J.; Park, S.H.; Park, S.Y. An increase in mean platelet volume during admission can predict the prognoses of patients with pneumonia in the intensive care unit: A retrospective study. PLoS ONE 2018, 13, e0208715. [Google Scholar] [CrossRef]
- Vardon-Bounes, F.; Mémier, V.; Marcaud, M.; Jacquemin, A.; Hamzeh-Cognasse, H.; Garcia, C.; Series, J.; Sié, P.; Minville, V.; Gratacap, M.P.; et al. Platelet activation and prothrombotic properties in a mouse model of peritoneal sepsis. Sci. Rep. 2018, 8, 13536. [Google Scholar] [CrossRef]
- Kirschenbaum, L.A.; Adler, D.; Astiz, M.E.; Barua, R.S.; Saha, D.; Rackow, E.C. Mechanisms of platelet-neutrophil interactions and effects on cell filtration in septic shock. Shock 2002, 17, 508–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freishtat, R.J.; Natale, J.; Benton, A.S.; Cohen, J.; Sharron, M.; Wiles, A.A.; Ngor, W.M.; Mojgani, B.; Bradbury, M.; Degnan, A.; et al. Sepsis alters the megakaryocyte-platelet transcriptional axis resulting in granzyme B-mediated lymphotoxicity. Am J Respir Crit. Care Med. 2009, 179, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukai, N.; Nakayama, Y.; Ishi, S.; Ogawa, S.; Maeda, S.; Anada, N.; Murakami, S.; Mizobe, T.; Sawa, T.; Nakajima, Y. Changes in MicroRNA Expression Level of Circulating Platelets Contribute to Platelet Defect After Cardiopulmonary Bypass. Crit. Care Med. 2018, 46, e761–e767. [Google Scholar] [CrossRef]
- Elgheznawy, A.; Shi, L.; Hu, J.; Wittig, I.; Laban, H.; Pircher, J.; Mann, A.; Provost, P.; Randriamboavonjy, V.; Fleming, I. Dicer cleavage by calpain determines platelet microRNA levels and function in diabetes. Circ. Res. 2015, 117, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, J.; Blume, M.; Petit-Teixeira, E.; Hugo Teixeira, V.; Steiner, A.; Quente, E.; Wolfram, G.; Scholz, M.; Pierlot, C.; Migliorini, P.; et al. Cellular adhesion gene SELP is associated with rheumatoid arthritis and displays differential allelic expression. PLoS ONE 2014, 9, e103872. [Google Scholar] [CrossRef] [Green Version]
- Tregouet, D.A.; Barbaux, S.; Escolano, S.; Tahri, N.; Golmard, J.L.; Tiret, L.; Cambien, F. Specific haplotypes of the P-selectin gene are associated with myocardial infarction. Hum. Mol. Genet. 2002, 11, 2015–2023. [Google Scholar] [CrossRef] [Green Version]
- Bone, R.C.; Balk, R.A.; Cerra, F.B.; Dellinger, R.P.; Fein, A.M.; Knaus, W.A.; Schein, R.M.; Sibbald, W.J. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest 1992, 101, 1644–1655. [Google Scholar] [CrossRef] [Green Version]
- Czimmerer, Z.; Hulvely, J.; Simandi, Z.; Varallyay, E.; Havelda, Z.; Szabo, E.; Varga, A.; Dezso, B.; Balogh, M.; Horvath, A.; et al. A versatile method to design stem-loop primer-based quantitative PCR assays for detecting small regulatory RNA molecules. PLoS ONE 2013, 8, e55168. [Google Scholar] [CrossRef] [Green Version]
- Fejes, Z.; Czimmerer, Z.; Szük, T.; Póliska, S.; Horváth, A.; Balogh, E.; Jeney, V.; Váradi, J.; Fenyvesi, F.; Balla, G.; et al. Endothelial cell activation is attenuated by everolimus via transcriptional and post-transcriptional regulatory mechanisms after drug-eluting coronary stenting. PLoS ONE 2018, 13, e0197890. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Septic Patients (n = 21) | Controls (n = 21) | p Value |
|---|---|---|---|
| Age (years) | 64 (51–70) | 58 (42–65) | n.s. |
| Male/female gender (n) | 16/5 | 14/7 | n.s. |
| WBC count (G/L) | 11.4 (8.3–16.2) | 7.6 (6.2–8.9) | p < 0.001 |
| PLT count (G/L) | 218 (175–264) | 332 (290–365) | p < 0.01 |
| Serum CRP (mg/L) | 210.5 ± 98.2 | 1.4 ± 1.0 | p < 0.001 |
| Serum PCT (µg/L) | 27.4 ± 11.7 | n/a | - |
| SOFA-score | 11 (9–13) | n/a | - |
| Sepsis/septic shock (n) | 14/7 | n/a | - |
| ICU length of stay (days) | 25.1 ± 14.4 | n/a | - |
| 28-day mortality (n) | 9 | n/a | - |
| Source of infection—pneumonia (n) | 18 | n/a | - |
| Source of infection—urinary tract (n) | 3 | n/a | - |
| Organism—Gram-positive bacteria (n) | 6 | n/a | - |
| Organism—Gram-negative bacteria (n) | 12 | n/a | - |
| Unknown infection (n) | 3 | n/a | - |
| Anti-platelet therapy (n) | 16 | 12 | n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szilágyi, B.; Fejes, Z.; Póliska, S.; Pócsi, M.; Czimmerer, Z.; Patsalos, A.; Fenyvesi, F.; Rusznyák, Á.; Nagy, G.; Kerekes, G.; et al. Reduced miR-26b Expression in Megakaryocytes and Platelets Contributes to Elevated Level of Platelet Activation Status in Sepsis. Int. J. Mol. Sci. 2020, 21, 866. https://doi.org/10.3390/ijms21030866
Szilágyi B, Fejes Z, Póliska S, Pócsi M, Czimmerer Z, Patsalos A, Fenyvesi F, Rusznyák Á, Nagy G, Kerekes G, et al. Reduced miR-26b Expression in Megakaryocytes and Platelets Contributes to Elevated Level of Platelet Activation Status in Sepsis. International Journal of Molecular Sciences. 2020; 21(3):866. https://doi.org/10.3390/ijms21030866
Chicago/Turabian StyleSzilágyi, Bernadett, Zsolt Fejes, Szilárd Póliska, Marianna Pócsi, Zsolt Czimmerer, Andreas Patsalos, Ferenc Fenyvesi, Ágnes Rusznyák, György Nagy, György Kerekes, and et al. 2020. "Reduced miR-26b Expression in Megakaryocytes and Platelets Contributes to Elevated Level of Platelet Activation Status in Sepsis" International Journal of Molecular Sciences 21, no. 3: 866. https://doi.org/10.3390/ijms21030866
APA StyleSzilágyi, B., Fejes, Z., Póliska, S., Pócsi, M., Czimmerer, Z., Patsalos, A., Fenyvesi, F., Rusznyák, Á., Nagy, G., Kerekes, G., Berhés, M., Szűcs, I., Kunapuli, S. P., Kappelmayer, J., & Nagy, B., Jr. (2020). Reduced miR-26b Expression in Megakaryocytes and Platelets Contributes to Elevated Level of Platelet Activation Status in Sepsis. International Journal of Molecular Sciences, 21(3), 866. https://doi.org/10.3390/ijms21030866