Nitric Oxide and Hydrogen Peroxide Signaling in Extractive Shiraia Fermentation by Triton X-100 for Hypocrellin A Production


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
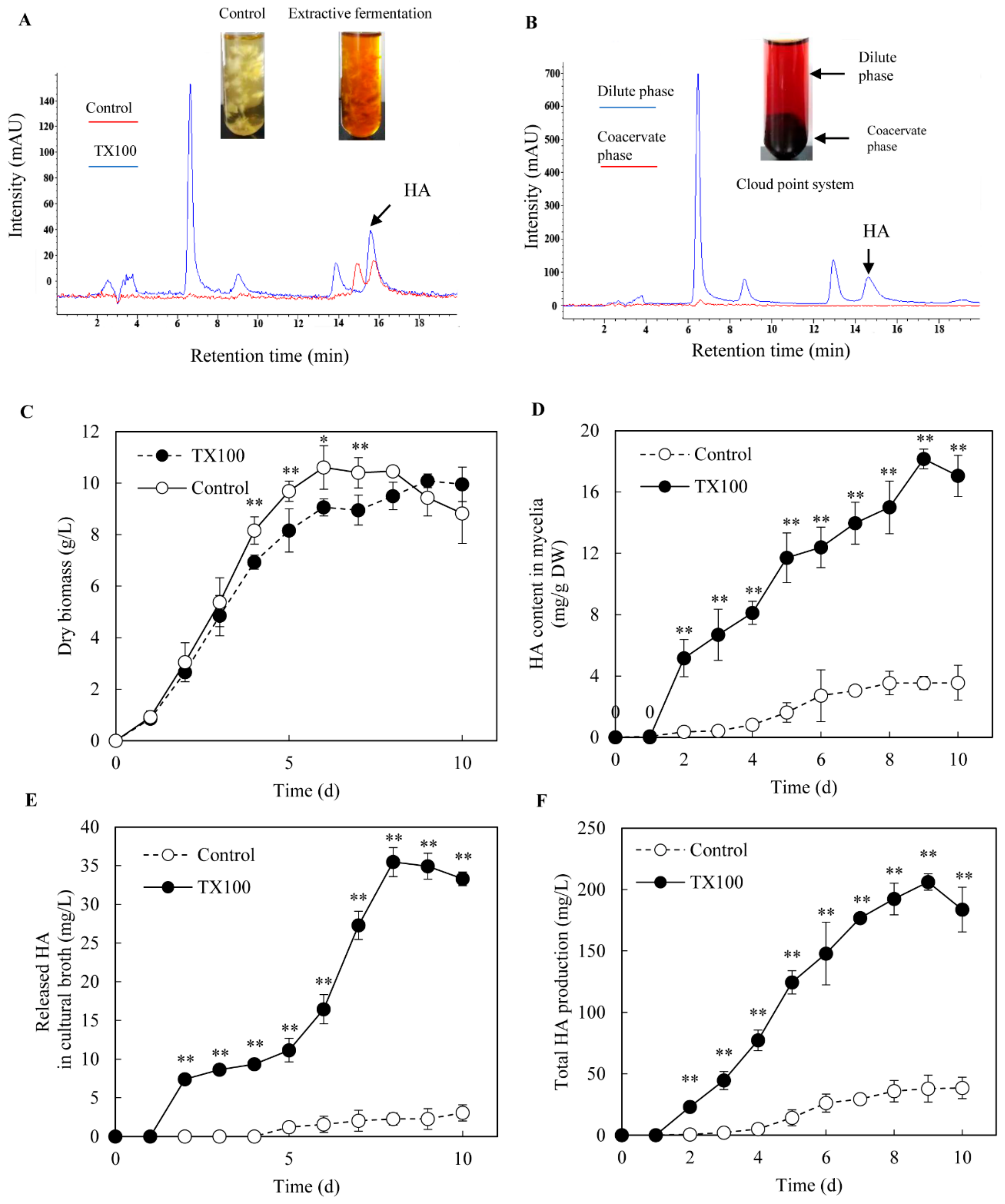
2.1. Extractive Fermentation in Micelle Aqueous Solution
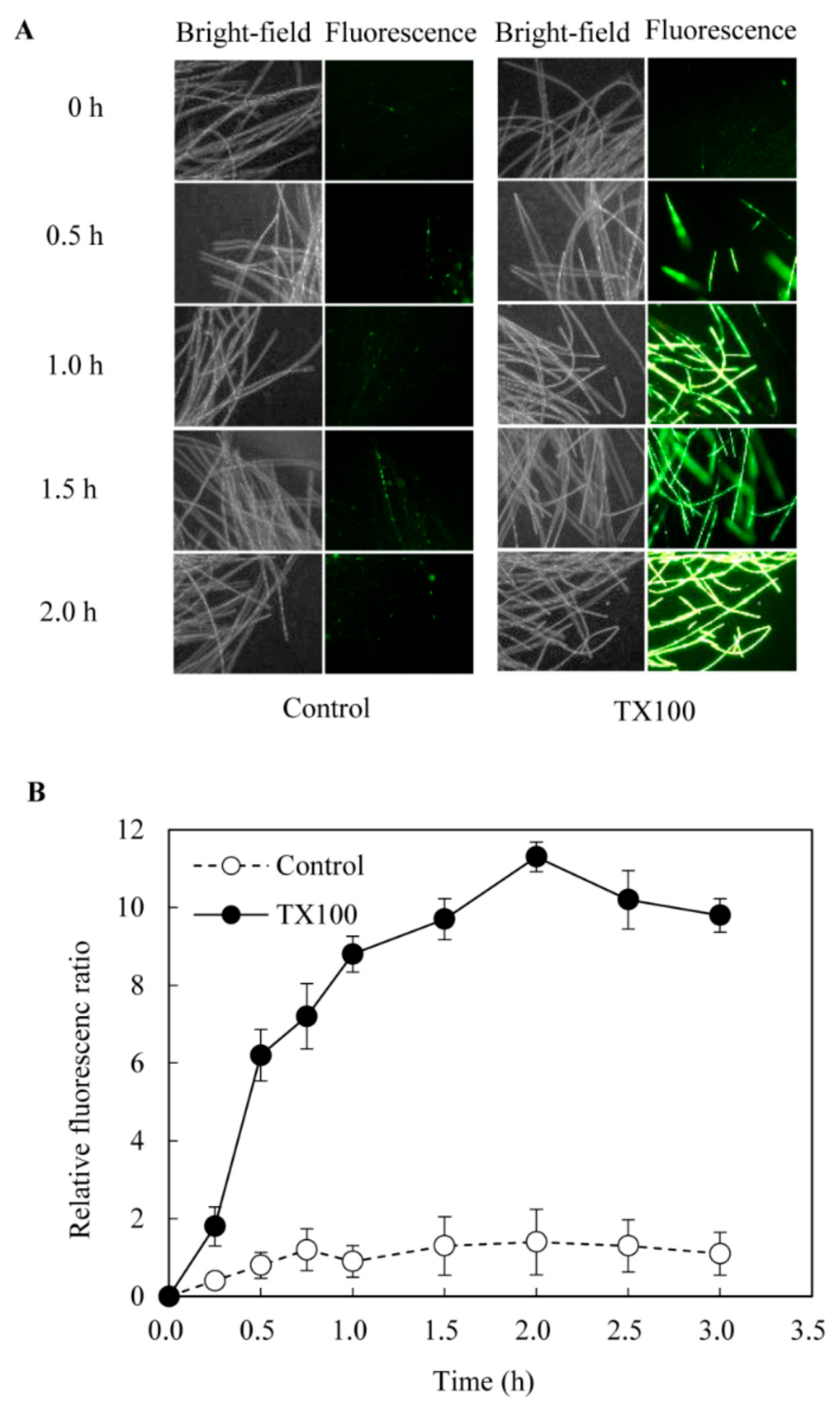
2.2. Effect of TX100 on Fungal Membrane Permeabilization
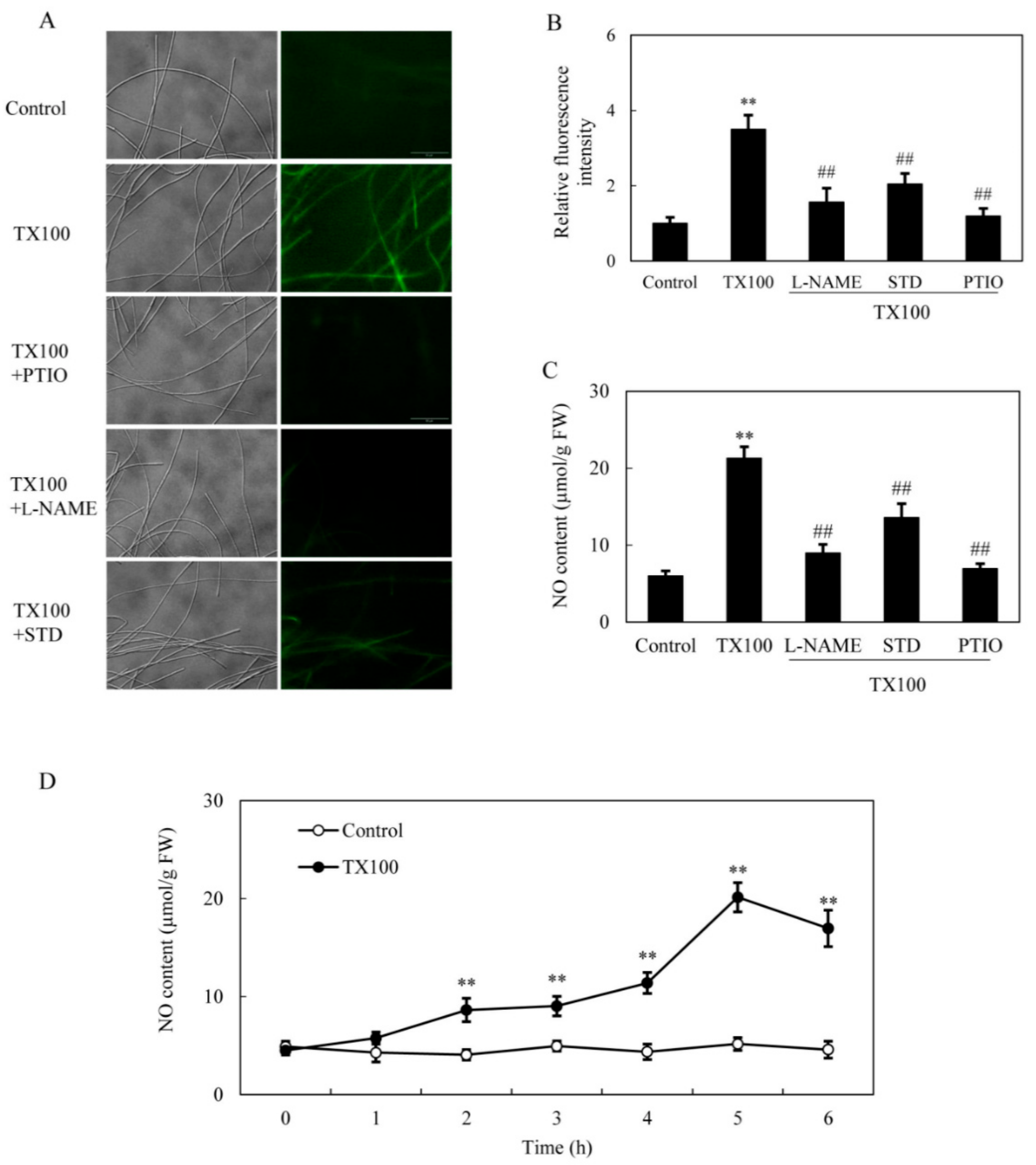
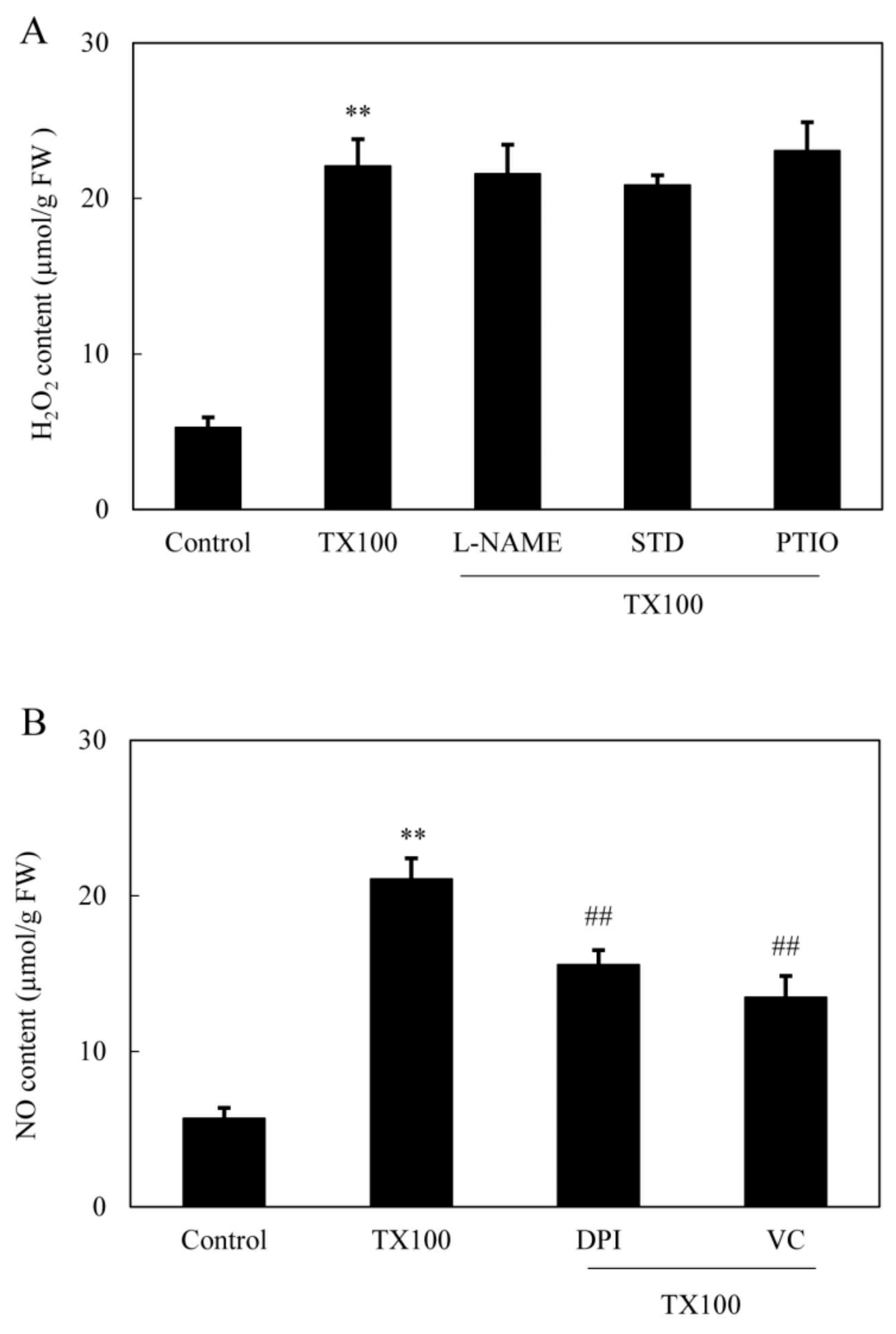
2.3. NO Generation in the Extractive Fermentation
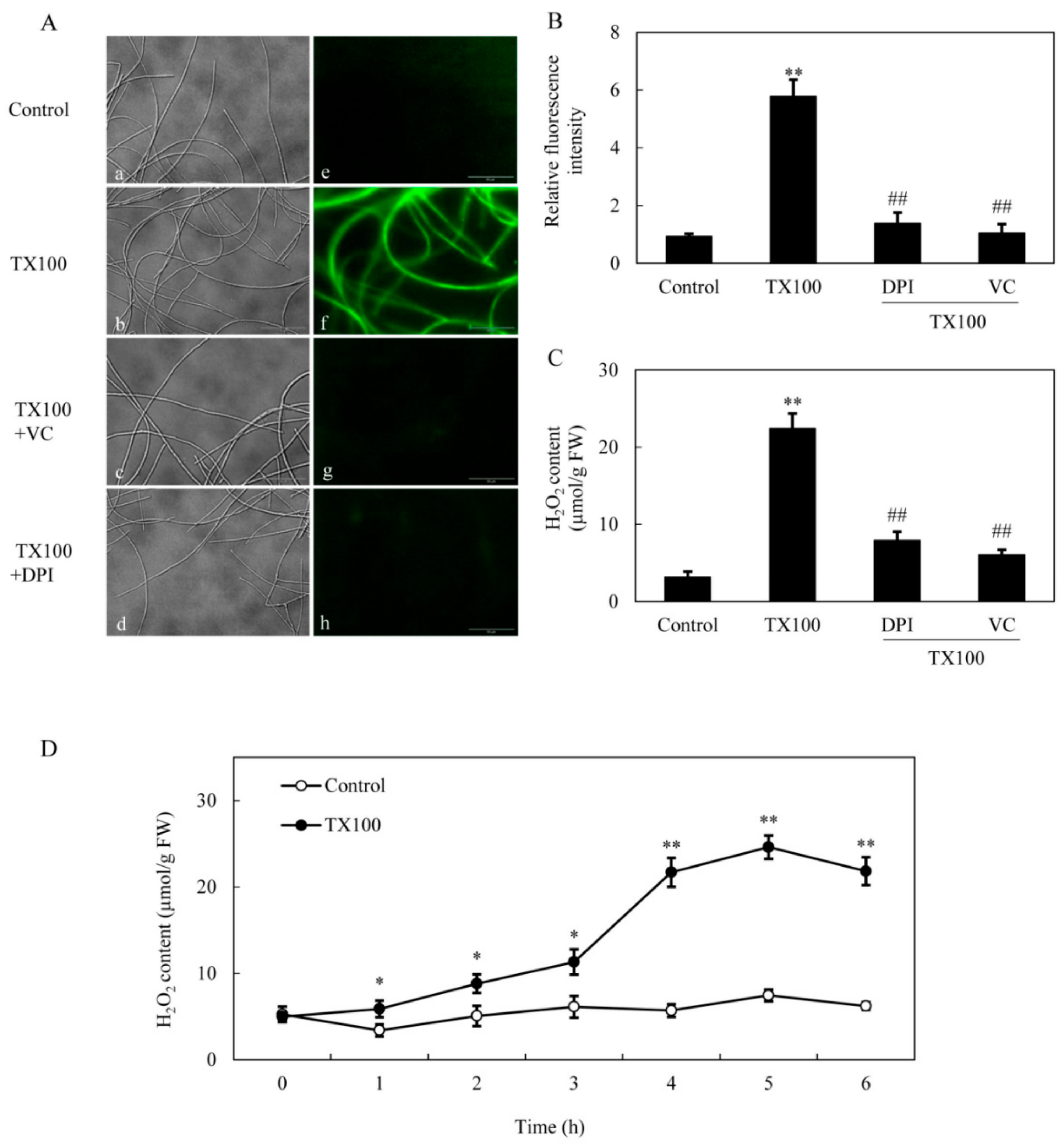
2.4. ROS Production in the Extractive Fermentation
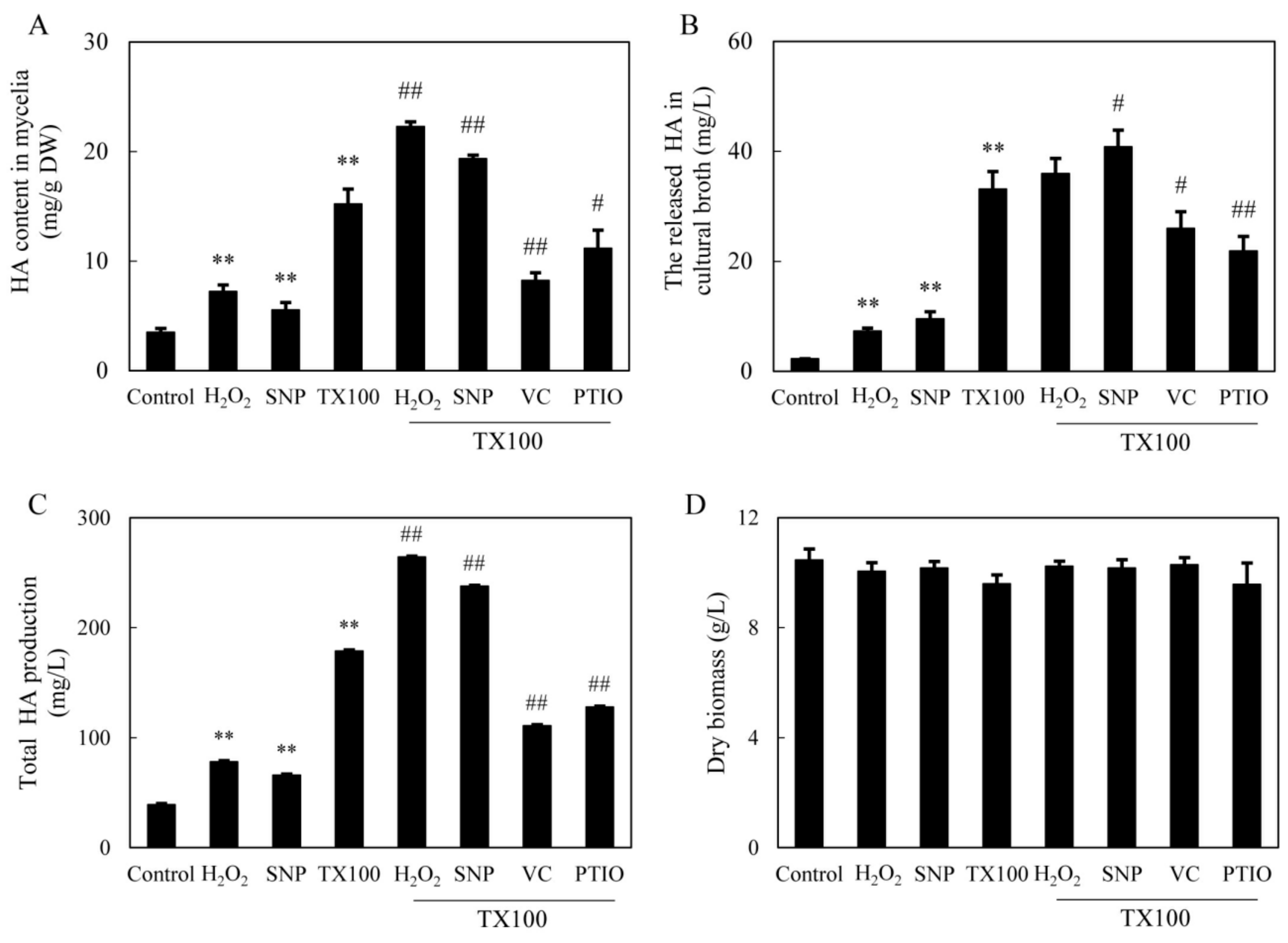
2.5. Effect of NO and ROS on HA Production
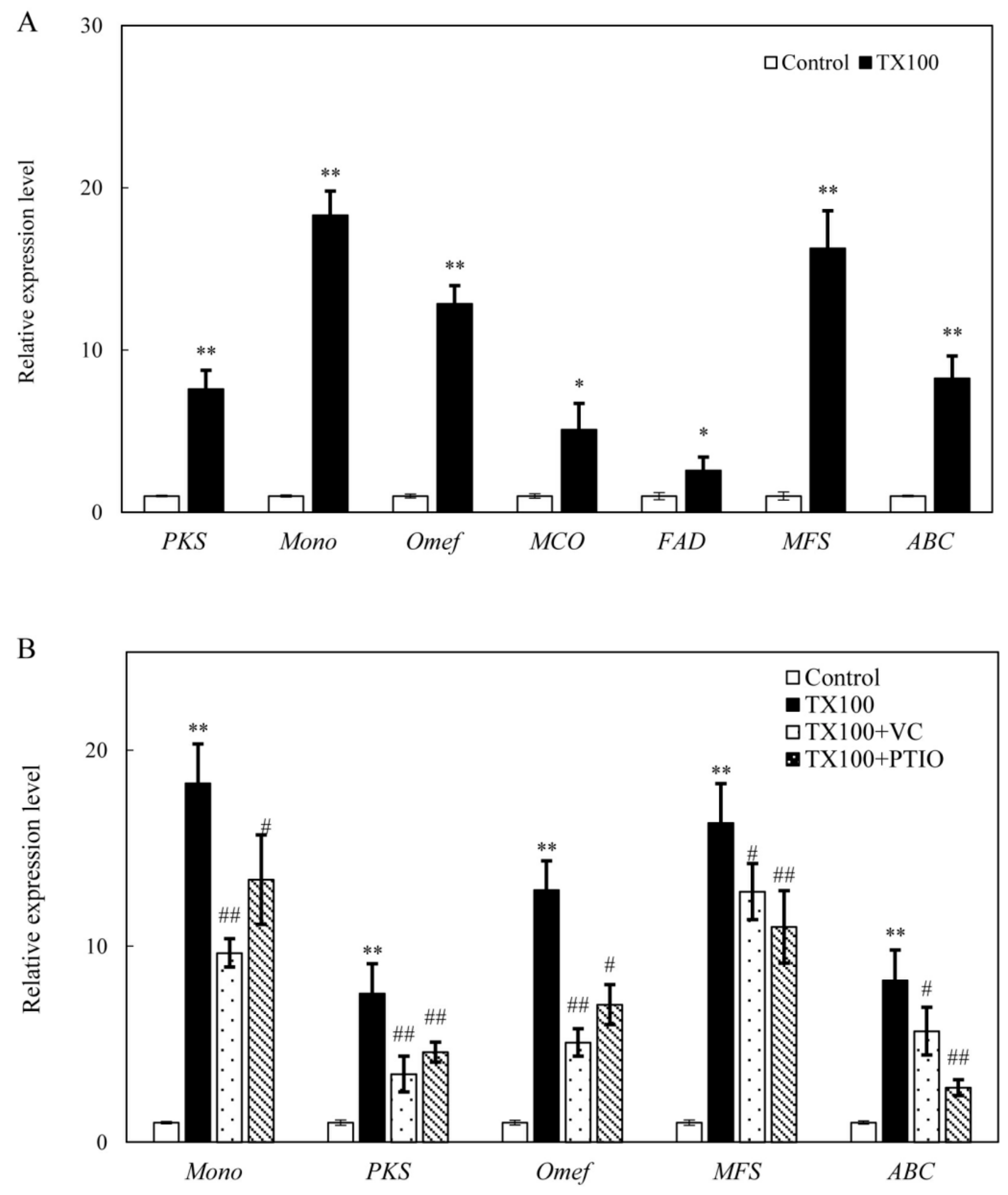
2.6. Effect of NO and ROS on Gene Expression for HA Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Membrane Permeabilization Assay
4.3. Detection of NO and ROS Generation
4.4. Inhibition Experiments on NO and ROS Effects
4.5. Extraction and Quantification of HA
4.6. Quantitative Real-time PCR
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| DAF-2 DA | 4, 5-diaminofluorescein diacetate |
| DCFH-DA | 2, 7-dichlorodihydro fluorescein diacetate |
| DPI | diphenylene iodonium |
| FAD | FAD/FMN-containing dehydrogenase |
| FW | fresh weight |
| H2O2 | hydrogen peroxide |
| HA | hypocrellin A |
| L-NAME | Nω-nitro-L-arginine methyl ester |
| MCO | multicopper oxidase |
| MFS | major facilitator superfamily |
| Mono | monooxygenase |
| NO | nitric oxide synthase |
| NOX | NADPH oxidase |
| NR | nitrate reductase |
| Omef | O-methyl-transferase |
| PDA | potato dextrose agar |
| PDT | photodynamic therapy |
| PKS | polyketide synthase |
| PTIO | 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide |
| ROS | reactive oxygen species |
| SDS | sodium dodecyl sulfate |
| SNP | sodium nitroprusside |
| STD | sodium tungstate dehydrate |
| TX100 | Triton X-100 |
| VC | vitamin C |
References
- Zhang, Z.Y.; Wang, N.H.; Wan, Q.; Li, M.F. EPR studies of singlet oxygen (1O2) and free radicals (O2−, OH, HB−) generated during photosensitization of hypocrellin B. Free Radic. Biol. Med. 1993, 14, 1–9. [Google Scholar] [CrossRef]
- Chio-Srichan, S.; Oudrhiri, N.; Bennaceur-Griscelli, A.; Turhan, A.G.; Dumas, P.; Refregiers, M. Toxicity and phototoxicity of hypocrellin A on malignant human cell lines, evidence of a synergistic action of photodynamic therapy with Imatinib mesylate. J. Photochem. Photobiol. B-Biol. 2010, 99, 100–104. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.M.; Morgan, B.J.; Mulrooney, C.A.; Carroll, P.J.; Kozlowski, M.C. Perylenequinone natural products: Total synthesis of hypocrellin A. J. Org. Chem. 2010, 75, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Z.W.; Mao, L.W.; Liang, H.L.; Zhang, Z.B.; Wang, Y.; Yan, R.M. Simultaneous determination of six perylenequinones in Shiraia sp. Slf14 by HPLC. J. Liq. Chromatogr. Relat. Technol. 2017, 40, 536–540. [Google Scholar] [CrossRef]
- Du, W.; Liang, Z.Q.; Zou, X.; Han, Y.F.; Liang, J.D.; Yu, J.P.; Chen, W.H.; Wang, Y.R.; Sun, C.L. Effects of microbial elicitor on production of hypocrellin by Shiraia bambusicola. Folia Microbiol. 2013, 58, 283–289. [Google Scholar] [CrossRef]
- Du, W.; Sun, C.L.; Wang, B.G.; Wang, Y.M.; Dong, B.; Liu, J.H.; Xia, J.B.; Xie, W.J.; Wang, J.; Sun, J.K.; et al. Response mechanism of hypocrellin colorants biosynthesis by Shiraia bambusicola to elicitor PB90. AMB Express 2019, 9, 146. [Google Scholar] [CrossRef]
- Sun, C.X.; Ma, Y.J.; Wang, J.W. Enhanced production of hypocrellin A by ultrasound stimulation in submerged cultures of Shiraia bambusicola. Ultrason. Sonochem. 2017, 38, 214–224. [Google Scholar] [CrossRef]
- Gao, R.J.; Xu, Z.C.; Deng, H.X.; Guan, Z.B.; Liao, X.R.; Zhao, Y.; Zheng, X.H.; Cai, Y.J. Influences of light on growth, reproduction and hypocrellin production by Shiraia sp. SUPER-H168. Arch. Microbiol. 2018, 200, 1217–1225. [Google Scholar] [CrossRef]
- Ma, Y.J.; Sun, C.X.; Wang, J.W. Enhanced production of Hypocrellin A in submerged cultures of Shiraia bambusicola by red light. Photochem. Photobiol. 2019, 95, 812–822. [Google Scholar] [CrossRef]
- Liu, X.Y.; Shen, X.Y.; Fan, L.; Gao, J.; Hou, C.L. High-efficiency biosynthesis of hypocrellin A in Shiraia sp. using gamma-ray mutagenesis. Appl. Microbiol. Biot. 2016, 100, 4875–4883. [Google Scholar] [CrossRef]
- Wang, Z.L.; Dai, Z.W. Extractive microbial fermentation in cloud point system. Enzym. Microb. Tech. 2010, 46, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Hinze, W.L.; Pramauro, E. A critical review of surfactant-mediated phase separations (cloud-point extractions): Theory and applications. Crit. Rev. Anal. Chem. 1993, 24, 133–177. [Google Scholar] [CrossRef]
- Hu, Z.Q.; Zhang, X.H.; Wu, Z.Q.; Qi, H.S.; Wang, Z.L. Export of intracellular Monascus pigments by two-stage microbial fermentation in nonionic surfactant micelle aqueous solution. J. Biotechnol. 2012, 162, 202–209. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Oliveira, J.; Sousa-Gallagher, M.; Méndez-Zavala, A.; Montañez, J.C. Perstraction of intracellular pigments through submerged fermentation of Talaromyces spp. in a surfactant rich media: A novel approach for enhanced pigment recovery. J. Fungi 2017, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.Y.; Qian, C.; Wang, Z.L.; Xu, J.H.; Yang, R.D.; Qi, H.S. Investigation of extractive microbial transformation in nonionic surfactant micelle aqueous solution using response surface methodology. Appl. Microbiol. Biotechnol. 2010, 85, 517–524. [Google Scholar] [CrossRef]
- Wang, Z.L.; Zhao, F.S.; Hao, X.Q.; Chen, D.J.; Li, D.T. Microbial transformation of hydrophobic compound in cloud point system. J. Mol. Catal. B-Enzym. 2004, 27, 147–153. [Google Scholar] [CrossRef]
- Cai, Y.J.; Liao, X.H.; Liang, X.R.; Ding, Y.R.; Sun, J.; Zhang, D.B. Induction of hypocrellin production by Triton X-100 under submerged fermentation with Shiraia sp. SUPER-H168. New Biotech. 2011, 28, 588–592. [Google Scholar] [CrossRef]
- Lei, X.Y.; Zhang, M.Y.; Ma, Y.J.; Wang, J.W. Transcriptomic responses involved in enhanced production of hypocrellin A by addition of Triton X-100 in submerged cultures of Shiraia bambusicola. J. Ind. Microbiol. Biotechnol. 2017, 44, 1415–1429. [Google Scholar] [CrossRef]
- Callow, N.V.; Ju, L.K. Promoting pellet growth of Trichoderma reesei Rut C30 by surfactants for easy separation and enhanced cellulase production. Enzym. Microb. Technol. 2012, 50, 311–317. [Google Scholar] [CrossRef]
- Hu, Z.Q.; Zhang, X.H.; Wu, Z.Q.; Qi, H.S.; Wang, Z.L. Perstraction of intracellular pigments by submerged cultivation of Monascus in nonionic surfactant micelle aqueous solution. Appl. Microbiol. Biotechnol. 2012, 94, 81–89. [Google Scholar] [CrossRef]
- Li, M.H. Effects of nonionic and ionic surfactants on survival, oxidative stress, and cholinesterase activity of planarian. Chemosphere 2008, 70, 1796–1803. [Google Scholar] [CrossRef] [PubMed]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Wendehenne, D.; Hancock, J.T. New frontiers in nitric oxide biology in plant. Plant Sci. 2011, 181, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Zheng, L.P.; Zhang, B.; Zou, T. Stimulation of artemisinin synthesis by combined cerebroside and nitric oxide elicitation in Artemisia annua hairy roots. Appl. Microbiol. Biotechnol. 2009, 85, 285–292. [Google Scholar] [CrossRef]
- Wang, J.W.; Zheng, L.P.; Wu, J.Y.; Tan, R.X. Involvement of nitric oxide in oxidative burst, phenylalanine ammonia-lyase activation and Taxol production induced by low-energy ultrasound in Taxus yunnanensis cell suspension cultures. Nitric Oxide 2006, 15, 351–358. [Google Scholar] [CrossRef]
- Wang, C.H.; Zheng, L.P.; Tian, H.; Wang, J.W. Synergistic effects of ultraviolet-B and methyl jasmonate on tanshinone biosynthesis in Salvia miltiorrhiza hairy roots. J. Photochem. Photobiol. B-Biol. 2016, 159, 93–100. [Google Scholar] [CrossRef]
- Ma, Y.J.; Zheng, L.P.; Wang, J.W. Bacteria associated with Shiraia fruiting bodies influence fungal production of hypocrellin A. Front. Microbiol. 2019, 10, 2023. [Google Scholar] [CrossRef]
- Zheng, W.F.; Miao, K.J.; Zhang, Y.X.; Pan, S.Y.; Zhang, M.M.; Jiang, H. Nitric oxide mediates the fungal-elicitor-enhanced biosynthesis of antioxidant polyphenols in submerged cultures of Inonotus obliquus. Microbiology 2009, 155, 3440–3448. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.X.; Xi, Q.; Xu, Q.; He, M.H.; Ding, J.N.; Dai, Y.C.; Keller, N.P.; Zheng, W.F. Correlation of nitric oxide produced by an inducible nitric oxide synthase-like protein with enhanced expression of the phenylpropanoid pathway in Inonotus obliquus cocultured with Phellinus morii. Appl. Microbiol. Biotechnol. 2015, 99, 4361–4372. [Google Scholar] [CrossRef]
- Gu, L.; Zhong, X.; Lian, D.H.; Zheng, Y.M.; Wang, H.Z.; Liu, X. Triterpenoid biosynthesis and the transcriptional response elicited by nitric oxide in submerged fermenting Ganoderma lucidum. Process Biochem. 2017, 60, 19–26. [Google Scholar] [CrossRef]
- Du, W.; Sun, C.L.; Wang, J.; Xie, W.J.; Wang, B.Q.; Liu, X.H.; Zhang, Y.M.; Fan, Y.H. Conditions and regulation of mixed culture to promote Shiraia bambusicola and Phoma sp. BZJ6 for laccase production. Sci. Rep. 2017, 7, 17801. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Liang, J.D.; Han, Y.F.; Yu, J.P.; Liang, Z.Q. Nitric oxide mediates hypocrellin accumulation induced by fungal elicitor in submerged cultures of Shiraia bambusicola. Biotechnol. Lett. 2015, 37, 153–159. [Google Scholar] [CrossRef]
- Ninnemann, H.; Maier, J. Indications for the occurrence of nitric oxide synthases in fungi and plants and the involvement in photoconidiation of Neurospora crassa. Photochem. Photobiol. 1996, 64, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Samalova, M.; Johnson, J.; IIIes, M.; Kelly, S.; Fricker, M.; Gurr, S. Nitric oxide generated by the rice blast fungus Magnaporthe oryzae drives plant infection. New Phytol. 2013, 197, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Marcos, A.T.; Ramos, M.S.; Marcos, J.F.; Carmona, L.; Strauss, J.; Cánovas, D. Nitric oxide synthesis by nitrate reductase is regulated during development in Aspergillus. Mol. Microbiol. 2016, 99, 15–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, N.; Gonugunta, V.K.; Puli, M.R.; Raghavendra, A.S. Nitric oxide production occurs downstream of reactive oxygen species in guard cells during stomatal closure induced by chitosan in abaxial epidermis of Pisum sativum. Planta 2009, 229, 757–765. [Google Scholar] [CrossRef]
- Sun, C.X.; Ma, Y.J.; Wang, J.W. Improved hypocrellin A production in Shiraia bambusicola by light-dark shift. J. Photochem. Photobiol. B-Biol. 2018, 182, 100–107. [Google Scholar] [CrossRef]
- Lu, C.S.; Ma, Y.J.; Wang, J.W. Lanthanum elicitation on hypocrellin A production in mycelium cultures of Shiraia bambusicola is mediated by ROS generation. J. Rare Earth. 2019, 37, 895–902. [Google Scholar] [CrossRef]
- Yang, H.L.; Wang, Y.; Zhang, Z.B.; Yan, R.M.; Zhu, D. Whole-genome shotgun assembly and analysis of the genome of Shiraia sp. strain Slf14, a novel endophytic fungus producing huperzine A and hypocrellin A. Genome Announc. 2014, 2, e00011–e00014. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Lin, X.; Qi, S.S.; Luo, Z.M.; Chen, S.L.; Yan, S.Z. De novo transcriptome assembly in Shiraia bambusicola to investigate putative genes involved in the biosynthesis of hypocrellin A. Int. J. Mol. Sci. 2016, 17, 311. [Google Scholar] [CrossRef]
- Deng, H.X.; Gao, R.J.; Liao, X.R.; Cai, Y.J. Characterization of a major facilitator superfamily transporter in Shiraia bambusicola. Res. Microbiol. 2017, 168, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Zheng, L.P.; Wang, J.W. Inducing perylenequinone production from a bambusicolous fungus Shiraia sp. S9 through co-culture with a fruiting body-associated bacterium Pseudomonas fulva SB1. Microb. Cell Fact. 2019, 18, 121. [Google Scholar] [CrossRef]
- Thevissen, K.; Terras, F.R.G.; Broekaert, W.F. Permeabilization of fungal membranes by plant defensins inhibits fungal growth. Appl. Environ. Microbiol. 1999, 65, 5451–5458. [Google Scholar] [CrossRef] [Green Version]
- Turrion-Gomez, J.L.; Benito, E.P. Flux of nitric oxide between the necrotrophic pathogen Botrytis cinerea and the host plant. Mol. Plant Pathol. 2011, 12, 606–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, S.N.; Gao, Z.J.; Wang, C.F.; Huang, L.L.; Kang, Z.S.; Zhang, H.C. Nitric oxide and reactive oxygen species coordinately regulate the germination of Puccinia striiformis f. sp. tritici urediniospores. Front. Microbiol. 2016, 7, 178. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.P.; Wang, Y.; Ma, Y.J.; Wang, J.W.; Zheng, L.P. Nitric Oxide and Hydrogen Peroxide Signaling in Extractive Shiraia Fermentation by Triton X-100 for Hypocrellin A Production. Int. J. Mol. Sci. 2020, 21, 882. https://doi.org/10.3390/ijms21030882
Li XP, Wang Y, Ma YJ, Wang JW, Zheng LP. Nitric Oxide and Hydrogen Peroxide Signaling in Extractive Shiraia Fermentation by Triton X-100 for Hypocrellin A Production. International Journal of Molecular Sciences. 2020; 21(3):882. https://doi.org/10.3390/ijms21030882
Chicago/Turabian StyleLi, Xin Ping, Yue Wang, Yan Jun Ma, Jian Wen Wang, and Li Ping Zheng. 2020. "Nitric Oxide and Hydrogen Peroxide Signaling in Extractive Shiraia Fermentation by Triton X-100 for Hypocrellin A Production" International Journal of Molecular Sciences 21, no. 3: 882. https://doi.org/10.3390/ijms21030882
APA StyleLi, X. P., Wang, Y., Ma, Y. J., Wang, J. W., & Zheng, L. P. (2020). Nitric Oxide and Hydrogen Peroxide Signaling in Extractive Shiraia Fermentation by Triton X-100 for Hypocrellin A Production. International Journal of Molecular Sciences, 21(3), 882. https://doi.org/10.3390/ijms21030882