Mechanisms and Regulation of Nonsense-Mediated mRNA Decay and Nonsense-Associated Altered Splicing in Lymphocytes


{kind=link}
{kind=link}
{kind=link}
Abstract
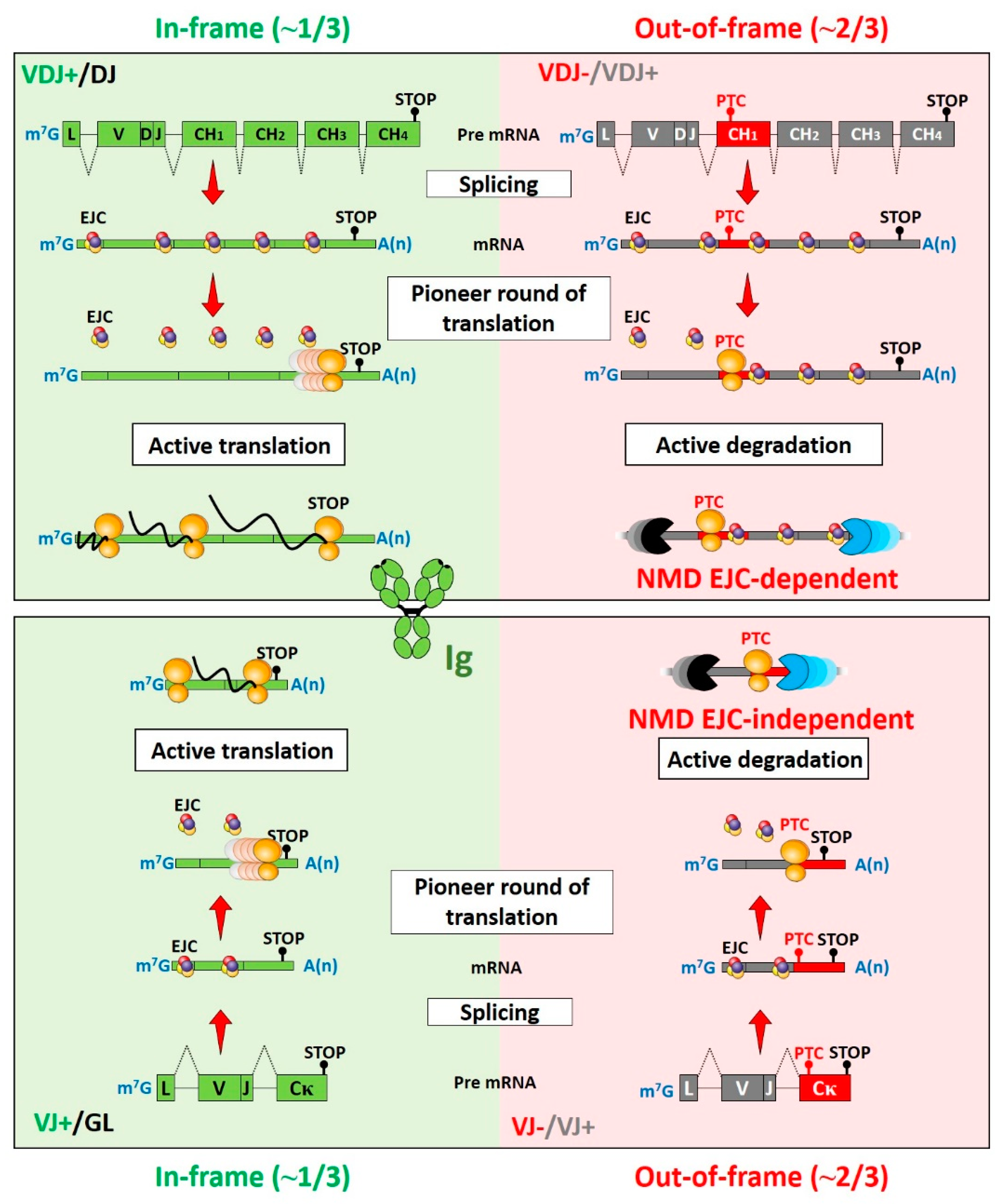
:1. Mechanisms of Nonsense-Mediated mRNA Decay (NMD)
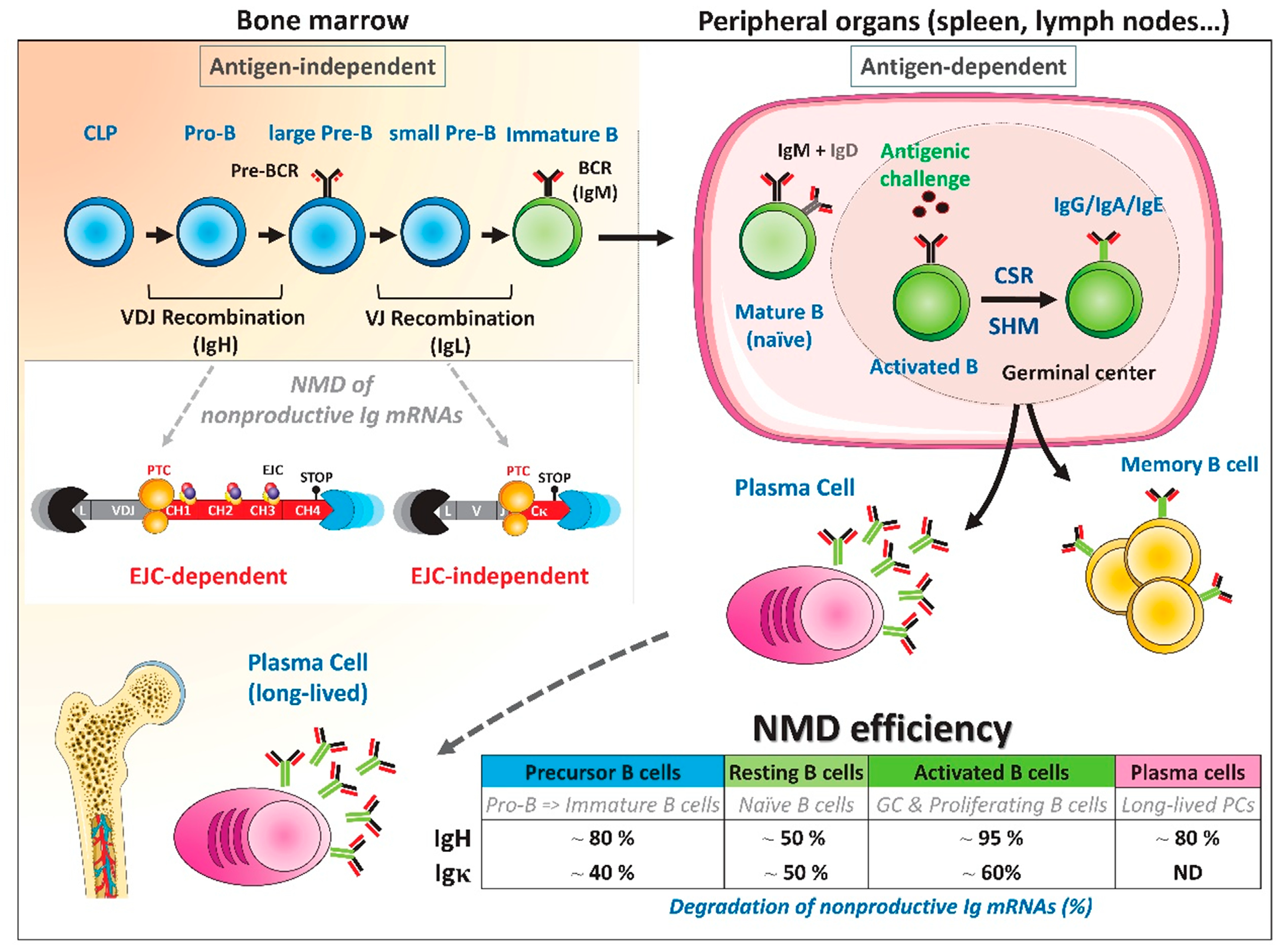
2. Fluctuations of NMD Efficiency during B-Cell Development
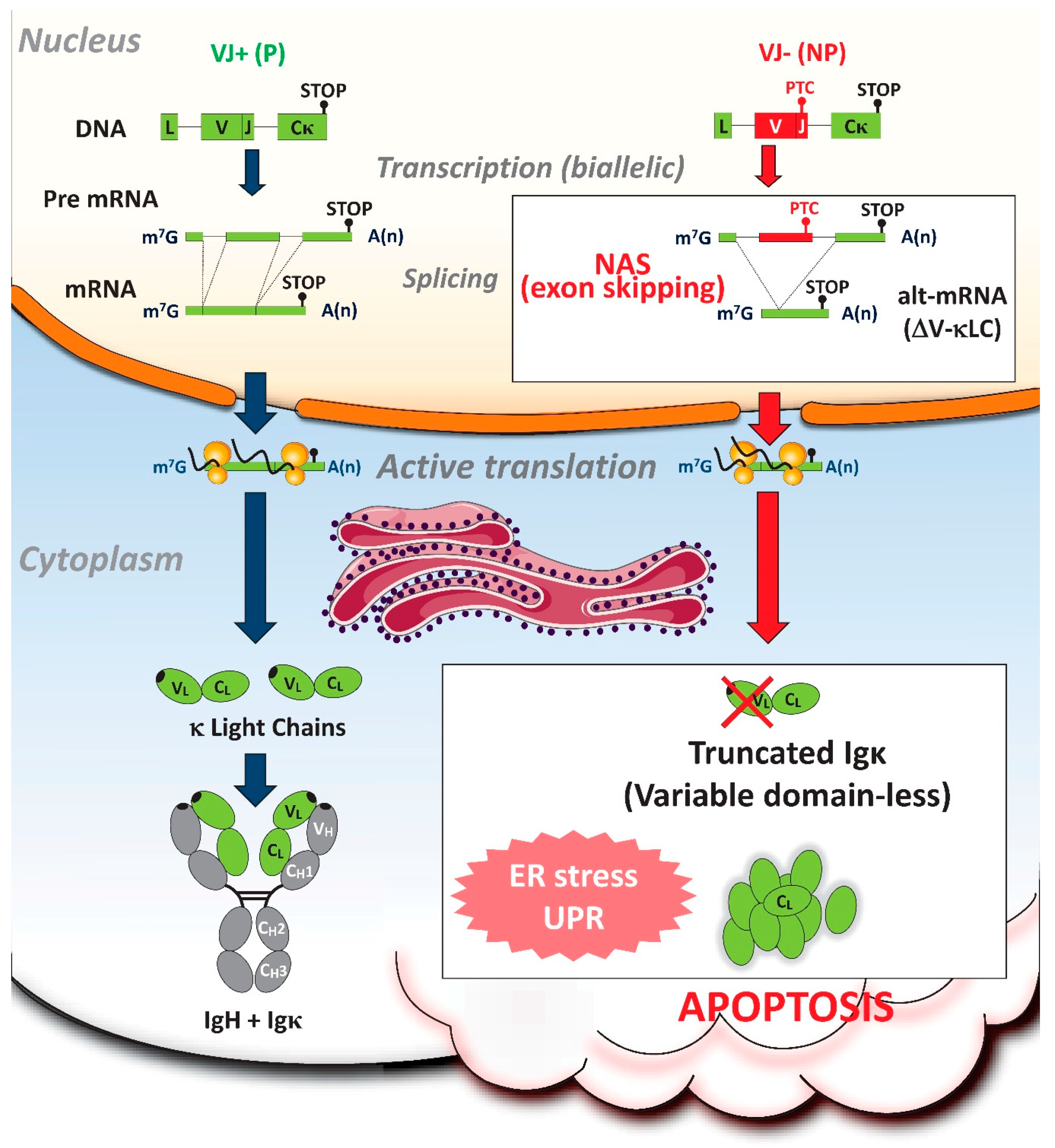
3. Nonsense-Associated Altered Splicing (NAS)
4. NAS of PTC-containing Ig RNAs during PC Differentiation
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kurosaki, T.; Maquat, L.E. Nonsense-Mediated mRNA decay in humans at a glance. J. Cell Sci. 2016, 129, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.C.; Temple, G.F.; Trecartin, R.F.; Kan, Y.W. Suppression of the nonsense mutation in homozygous beta 0 thalassaemia. Nature 1979, 281, 602–603. [Google Scholar] [CrossRef]
- Maquat, L.E.; Kinniburgh, A.J.; Rachmilewitz, E.A.; Ross, J. Unstable beta-Globin mRNA in mRNA-deficient beta o thalassemia. Cell 1981, 27, 543–553. [Google Scholar] [CrossRef]
- Mendell, J.T.; Sharifi, N.A.; Meyers, J.L.; Martinez-Murillo, F.; Dietz, H.C. Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nat. Genet. 2004, 36, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wilkinson, M.F. Nonsense surveillance in lymphocytes? Immunity 1998, 8, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Lykke-Andersen, S.; Jensen, T.H. Nonsense-Mediated mRNA decay: An intricate machinery that shapes transcriptomes. Nat. Rev. Mol. Cell Biol. 2015, 16, 665–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, P.; Yepiskoposyan, H.; Metze, S.; Zamudio Orozco, R.; Kleinschmidt, N.; Mühlemann, O. Nonsense-mediated mRNA decay in human cells: Mechanistic insights, functions beyond quality control and the double-Life of NMD factors. Cell. Mol. Life Sci. CMLS 2010, 67, 677–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebbapragada, I.; Lykke-Andersen, J. Execution of nonsense-Mediated mRNA decay: What defines a substrate? Curr. Opin. Cell Biol. 2009, 21, 394–402. [Google Scholar] [CrossRef]
- Rehwinkel, J.; Raes, J.; Izaurralde, E. Nonsense-Mediated mRNA decay: Target genes and functional diversification of effectors. Trends Biochem. Sci. 2006, 31, 639–646. [Google Scholar] [CrossRef]
- Schweingruber, C.; Rufener, S.C.; Zünd, D.; Yamashita, A.; Mühlemann, O. Nonsense-Mediated mRNA decay-Mechanisms of substrate mRNA recognition and degradation in mammalian cells. Biochim. Biophys. Acta 2013, 1829, 612–623. [Google Scholar] [CrossRef]
- Seyedali, A.; Berry, M.J. Nonsense-Mediated decay factors are involved in the regulation of selenoprotein mRNA levels during selenium deficiency. RNA 2014, 20, 1248–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frischmeyer, P.A.; Dietz, H.C. Nonsense-Mediated mRNA decay in health and disease. Hum. Mol. Genet. 1999, 8, 1893–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popp, M.W.; Maquat, L.E. Leveraging Rules of Nonsense-Mediated mRNA Decay for Genome Engineering and Personalized Medicine. Cell 2016, 165, 1319–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindeboom, R.G.H.; Supek, F.; Lehner, B. The rules and impact of nonsense-Mediated mRNA decay in human cancers. Nat. Genet. 2016, 48, 1112–1118. [Google Scholar] [CrossRef] [Green Version]
- Popp, M.W.; Maquat, L.E. Nonsense-Mediated mRNA Decay and Cancer. Curr. Opin. Genet. Dev. 2018, 48, 44–50. [Google Scholar] [CrossRef]
- Chamieh, H.; Ballut, L.; Bonneau, F.; Le Hir, H. NMD factors UPF2 and UPF3 bridge UPF1 to the exon junction complex and stimulate its RNA helicase activity. Nat. Struct. Mol. Biol. 2008, 15, 85–93. [Google Scholar] [CrossRef]
- Fiorini, F.; Boudvillain, M.; Le Hir, H. Tight intramolecular regulation of the human Upf1 helicase by its N- and C-Terminal domains. Nucleic Acids Res. 2013, 41, 2404–2415. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Li, Y.-R. Upf proteins: Highly conserved factors involved in nonsense mRNA mediated decay. Mol. Biol. Rep. 2018, 45, 39–55. [Google Scholar] [CrossRef]
- Shigeoka, T.; Kato, S.; Kawaichi, M.; Ishida, Y. Evidence that the Upf1-Related molecular motor scans the 3′-UTR to ensure mRNA integrity. Nucleic Acids Res. 2012, 40, 6887–6897. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, F.; Bagchi, D.; Le Hir, H.; Croquette, V. Human Upf1 is a highly processive RNA helicase and translocase with RNP remodelling activities. Nat. Commun. 2015, 6, 7581. [Google Scholar] [CrossRef] [Green Version]
- Franks, T.M.; Singh, G.; Lykke-Andersen, J. Upf1 ATPase-Dependent mRNP disassembly is required for completion of nonsense-Mediated mRNA decay. Cell 2010, 143, 938–950. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, P.V.; Gehring, N.H.; Kunz, J.B.; Hentze, M.W.; Kulozik, A.E. Interactions between UPF1, eRFs, PABP and the exon junction complex suggest an integrated model for mammalian NMD pathways. EMBO J. 2008, 27, 736–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashima, I.; Yamashita, A.; Izumi, N.; Kataoka, N.; Morishita, R.; Hoshino, S.; Ohno, M.; Dreyfuss, G.; Ohno, S. Binding of a novel SMG-1-Upf1-eRF1-eRF3 complex (SURF) to the exon junction complex triggers Upf1 phosphorylation and nonsense-mediated mRNA decay. Genes Dev. 2006, 20, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, A.; Izumi, N.; Kashima, I.; Ohnishi, T.; Saari, B.; Katsuhata, Y.; Muramatsu, R.; Morita, T.; Iwamatsu, A.; Hachiya, T.; et al. SMG-8 and SMG-9, two novel subunits of the SMG-1 complex, regulate remodeling of the mRNA surveillance complex during nonsense-mediated mRNA decay. Genes Dev. 2009, 23, 1091–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melero, R.; Uchiyama, A.; Castaño, R.; Kataoka, N.; Kurosawa, H.; Ohno, S.; Yamashita, A.; Llorca, O. Structures of SMG1-UPFs complexes: SMG1 contributes to regulate UPF2-dependent activation of UPF1 in NMD. Structure 2014, 22, 1105–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, N.; Cáceres, J.F. The RNA helicase DHX34 activates NMD by promoting a transition from the surveillance to the decay-Inducing complex. Cell Rep. 2014, 8, 1845–1856. [Google Scholar] [CrossRef] [Green Version]
- Melero, R.; Hug, N.; López-Perrote, A.; Yamashita, A.; Cáceres, J.F.; Llorca, O. The RNA helicase DHX34 functions as a scaffold for SMG1-mediated UPF1 phosphorylation. Nat. Commun. 2016, 7, 10585. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.B.F.; Ballut, L.; Johansen, J.S.; Chamieh, H.; Nielsen, K.H.; Oliveira, C.L.P.; Pedersen, J.S.; Séraphin, B.; Le Hir, H.; Andersen, G.R. Structure of the exon junction core complex with a trapped DEAD-box ATPase bound to RNA. Science 2006, 313, 1968–1972. [Google Scholar] [CrossRef]
- Le Hir, H.; Moore, M.J.; Maquat, L.E. Pre-mRNA splicing alters mRNP composition: Evidence for stable association of proteins at exon-Exon junctions. Genes Dev. 2000, 14, 1098–1108. [Google Scholar]
- Le Hir, H.; Izaurralde, E.; Maquat, L.E.; Moore, M.J. The spliceosome deposits multiple proteins 20–24 nucleotides upstream of mRNA exon-exon junctions. EMBO J. 2000, 19, 6860–6869. [Google Scholar] [CrossRef] [Green Version]
- Saulière, J.; Murigneux, V.; Wang, Z.; Marquenet, E.; Barbosa, I.; Le Tonquèze, O.; Audic, Y.; Paillard, L.; Roest Crollius, H.; Le Hir, H. CLIP-seq of eIF4AIII reveals transcriptome-Wide mapping of the human exon junction complex. Nat. Struct. Mol. Biol. 2012, 19, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kucukural, A.; Cenik, C.; Leszyk, J.D.; Shaffer, S.A.; Weng, Z.; Moore, M.J. The cellular EJC interactome reveals higher-Order mRNP structure and an EJC-SR protein nexus. Cell 2012, 151, 750–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, S.; Lykke-Andersen, J. Nonsense-Mediated mRNA decay occurs during eIF4F-Dependent translation in human cells. Nat. Struct. Mol. Biol. 2013, 20, 702–709. [Google Scholar] [CrossRef]
- Ishigaki, Y.; Li, X.; Serin, G.; Maquat, L.E. Evidence for a pioneer round of mRNA translation: mRNAs subject to nonsense-Mediated decay in mammalian cells are bound by CBP80 and CBP20. Cell 2001, 106, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Lejeune, F.; Ishigaki, Y.; Li, X.; Maquat, L.E. The exon junction complex is detected on CBP80-Bound but not eIF4E-Bound mRNA in mammalian cells: Dynamics of mRNP remodeling. EMBO J. 2002, 21, 3536–3545. [Google Scholar] [CrossRef] [Green Version]
- Rufener, S.C.; Mühlemann, O. eIF4E-Bound mRNPs are substrates for nonsense-Mediated mRNA decay in mammalian cells. Nat. Struct. Mol. Biol. 2013, 20, 710–717. [Google Scholar] [CrossRef]
- Le Hir, H.; Gatfield, D.; Izaurralde, E.; Moore, M.J. The exon-Exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-Mediated mRNA decay. EMBO J. 2001, 20, 4987–4997. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, H.; Saulière, J.; Wang, Z. The exon junction complex as a node of post-Transcriptional networks. Nat. Rev. Mol. Cell Biol. 2016, 17, 41–54. [Google Scholar] [CrossRef]
- Shum, E.Y.; Jones, S.H.; Shao, A.; Dumdie, J.; Krause, M.D.; Chan, W.-K.; Lou, C.-H.; Espinoza, J.L.; Song, H.-W.; Phan, M.H.; et al. The Antagonistic Gene Paralogs Upf3a and Upf3b Govern Nonsense-Mediated RNA Decay. Cell 2016, 165, 382–395. [Google Scholar] [CrossRef] [Green Version]
- Mühlemann, O.; Karousis, E.D. New functions in translation termination uncovered for NMD factor UPF3B. EMBO J. 2017, 36, 2928–2930. [Google Scholar] [CrossRef]
- Neu-Yilik, G.; Raimondeau, E.; Eliseev, B.; Yeramala, L.; Amthor, B.; Deniaud, A.; Huard, K.; Kerschgens, K.; Hentze, M.W.; Schaffitzel, C.; et al. Dual function of UPF3B in early and late translation termination. EMBO J. 2017, 36, 2968–2986. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Bonneau, F.; Schüssler, S.; Eppinger, E.; Conti, E. Phospho-Dependent and phospho-Independent interactions of the helicase UPF1 with the NMD factors SMG5-SMG7 and SMG6. Nucleic Acids Res. 2014, 42, 9447–9460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, S.; Franks, T.M.; Lykke-Andersen, J. Hyperphosphorylation amplifies UPF1 activity to resolve stalls in nonsense-Mediated mRNA decay. Nat. Commun. 2016, 7, 12434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, T.; Yamashita, A.; Kashima, I.; Schell, T.; Anders, K.R.; Grimson, A.; Hachiya, T.; Hentze, M.W.; Anderson, P.; Ohno, S. Phosphorylation of hUPF1 induces formation of mRNA surveillance complexes containing hSMG-5 and hSMG-7. Mol. Cell 2003, 12, 1187–1200. [Google Scholar] [CrossRef]
- Okada-Katsuhata, Y.; Yamashita, A.; Kutsuzawa, K.; Izumi, N.; Hirahara, F.; Ohno, S. N- and C-Terminal Upf1 phosphorylations create binding platforms for SMG-6 and SMG-5:SMG-7 during NMD. Nucleic Acids Res. 2012, 40, 1251–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberle, A.B.; Lykke-Andersen, S.; Mühlemann, O.; Jensen, T.H. SMG6 promotes endonucleolytic cleavage of nonsense mRNA in human cells. Nat. Struct. Mol. Biol. 2009, 16, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Glavan, F.; Behm-Ansmant, I.; Izaurralde, E.; Conti, E. Structures of the PIN domains of SMG6 and SMG5 reveal a nuclease within the mRNA surveillance complex. EMBO J. 2006, 25, 5117–5125. [Google Scholar] [CrossRef] [Green Version]
- Huntzinger, E.; Kashima, I.; Fauser, M.; Saulière, J.; Izaurralde, E. SMG6 is the catalytic endonuclease that cleaves mRNAs containing nonsense codons in metazoan. RNA 2008, 14, 2609–2617. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Kim, K.M.; Kim, Y.K. Human proline-Rich nuclear receptor coregulatory protein 2 mediates an interaction between mRNA surveillance machinery and decapping complex. Mol. Cell 2009, 33, 75–86. [Google Scholar] [CrossRef]
- Cho, H.; Han, S.; Choe, J.; Park, S.G.; Choi, S.S.; Kim, Y.K. SMG5-PNRC2 is functionally dominant compared with SMG5-SMG7 in mammalian nonsense-Mediated mRNA decay. Nucleic Acids Res. 2013, 41, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Loh, B.; Jonas, S.; Izaurralde, E. The SMG5-SMG7 heterodimer directly recruits the CCR4-NOT deadenylase complex to mRNAs containing nonsense codons via interaction with POP2. Genes Dev. 2013, 27, 2125–2138. [Google Scholar] [CrossRef] [Green Version]
- Lai, T.; Cho, H.; Liu, Z.; Bowler, M.W.; Piao, S.; Parker, R.; Kim, Y.K.; Song, H. Structural basis of the PNRC2-Mediated link between mrna surveillance and decapping. Struct. Lond. Engl. 1993 2012, 20, 2025–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, P.; Gkratsou, A.; Josi, C.; Colombo, M.; Mühlemann, O. Dissecting the functions of SMG5, SMG7, and PNRC2 in nonsense-mediated mRNA decay of human cells. RNA 2018, 24, 557–573. [Google Scholar] [PubMed] [Green Version]
- Gatfield, D.; Izaurralde, E. Nonsense-Mediated messenger RNA decay is initiated by endonucleolytic cleavage in Drosophila. Nature 2004, 429, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Lejeune, F.; Li, X.; Maquat, L.E. Nonsense-Mediated mRNA decay in mammalian cells involves decapping, deadenylating, and exonucleolytic activities. Mol. Cell 2003, 12, 675–687. [Google Scholar] [CrossRef]
- Casadio, A.; Longman, D.; Hug, N.; Delavaine, L.; Vallejos Baier, R.; Alonso, C.R.; Cáceres, J.F. Identification and characterization of novel factors that act in the nonsense-Mediated mRNA decay pathway in nematodes, flies and mammals. EMBO Rep. 2015, 16, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Balistreri, G.; Horvath, P.; Schweingruber, C.; Zünd, D.; McInerney, G.; Merits, A.; Mühlemann, O.; Azzalin, C.; Helenius, A. The host nonsense-Mediated mRNA decay pathway restricts Mammalian RNA virus replication. Cell Host Microbe 2014, 16, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Balistreri, G.; Bognanni, C.; Mühlemann, O. Virus Escape and Manipulation of Cellular Nonsense-Mediated mRNA Decay. Viruses 2017, 9, 24. [Google Scholar] [CrossRef]
- Garcia, D.; Garcia, S.; Voinnet, O. Nonsense-Mediated decay serves as a general viral restriction mechanism in plants. Cell Host Microbe 2014, 16, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Lykke-Andersen, J.; Bennett, E.J. Protecting the proteome: Eukaryotic cotranslational quality control pathways. J. Cell Biol. 2014, 204, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Mocquet, V.; Durand, S.; Jalinot, P. How Retroviruses Escape the Nonsense-Mediated mRNA Decay. AIDS Res. Hum. Retrovir. 2015, 31, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.L.; Wilusz, J. Cytoplasmic viruses: Rage against the (cellular RNA decay) machine. PLoS Pathog. 2013, 9, e1003762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasif, S.; Contu, L.; Mühlemann, O. Beyond quality control: The role of nonsense-mediated mRNA decay (NMD) in regulating gene expression. Semin. Cell Dev. Biol. 2018, 75, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Rigby, R.E.; Rehwinkel, J. RNA degradation in antiviral immunity and autoimmunity. Trends Immunol. 2015, 36, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, F.; Robin, J.-P.; Kanaan, J.; Borowiak, M.; Croquette, V.; Le Hir, H.; Jalinot, P.; Mocquet, V. HTLV-1 Tax plugs and freezes UPF1 helicase leading to nonsense-Mediated mRNA decay inhibition. Nat. Commun. 2018, 9, 431. [Google Scholar] [CrossRef] [PubMed]
- Mocquet, V.; Neusiedler, J.; Rende, F.; Cluet, D.; Robin, J.-P.; Terme, J.-M.; Duc Dodon, M.; Wittmann, J.; Morris, C.; Le Hir, H.; et al. The human T-Lymphotropic virus type 1 tax protein inhibits nonsense-mediated mRNA decay by interacting with INT6/EIF3E and UPF1. J. Virol. 2012, 86, 7530–7543. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, A.; Colognori, D.; Steitz, J.A. Human eIF4AIII interacts with an eIF4G-like partner, NOM1, revealing an evolutionarily conserved function outside the exon junction complex. Genes Dev. 2011, 25, 1078–1090. [Google Scholar] [CrossRef] [Green Version]
- Amrani, N.; Ganesan, R.; Kervestin, S.; Mangus, D.A.; Ghosh, S.; Jacobson, A. A faux 3′-UTR promotes aberrant termination and triggers nonsense-Mediated mRNA decay. Nature 2004, 432, 112–118. [Google Scholar] [CrossRef]
- Kervestin, S.; Jacobson, A. NMD: A multifaceted response to premature translational termination. Nat. Rev. Mol. Cell Biol. 2012, 13, 700–712. [Google Scholar] [CrossRef] [Green Version]
- Czaplinski, K.; Ruiz-Echevarria, M.J.; Paushkin, S.V.; Han, X.; Weng, Y.; Perlick, H.A.; Dietz, H.C.; Ter-Avanesyan, M.D.; Peltz, S.W. The surveillance complex interacts with the translation release factors to enhance termination and degrade aberrant mRNAs. Genes Dev. 1998, 12, 1665–1677. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.; Mikhailova, T.; Eliseev, B.; Yeramala, L.; Sokolova, E.; Susorov, D.; Shuvalov, A.; Schaffitzel, C.; Alkalaeva, E. PABP enhances release factor recruitment and stop codon recognition during translation termination. Nucleic Acids Res. 2016, 44, 7766–7776. [Google Scholar] [CrossRef] [PubMed]
- Amrani, N.; Dong, S.; He, F.; Ganesan, R.; Ghosh, S.; Kervestin, S.; Li, C.; Mangus, D.A.; Spatrick, P.; Jacobson, A. Aberrant termination triggers nonsense-Mediated mRNA decay. Biochem. Soc. Trans. 2006, 34, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Bühler, M.; Steiner, S.; Mohn, F.; Paillusson, A.; Mühlemann, O. EJC-Independent degradation of nonsense immunoglobulin-mu mRNA depends on 3′ UTR length. Nat. Struct. Mol. Biol. 2006, 13, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Metze, S.; Herzog, V.A.; Ruepp, M.-D.; Mühlemann, O. Comparison of EJC-Enhanced and EJC-independent NMD in human cells reveals two partially redundant degradation pathways. RNA 2013, 19, 1432–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatfield, D.; Unterholzner, L.; Ciccarelli, F.D.; Bork, P.; Izaurralde, E. Nonsense-Mediated mRNA decay in Drosophila: At the intersection of the yeast and mammalian pathways. EMBO J. 2003, 22, 3960–3970. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Maquat, L.E. Nonsense-Mediated mRNA decay (NMD) in animal embryogenesis: To die or not to die, that is the question. Curr. Opin. Genet. Dev. 2011, 21, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Brogna, S. Splicing-Dependent NMD does not require the EJC in Schizosaccharomyces pombe. EMBO J. 2010, 29, 1537–1551. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Yang, W.; Zhang, J.; Dang, H.; Lu, X.; Fu, C.; Miao, W. Nonsense-Mediated mRNA decay in Tetrahymena is EJC independent and requires a protozoa-specific nuclease. Nucleic Acids Res. 2017, 45, 6848–6863. [Google Scholar] [CrossRef]
- Gehring, N.H.; Kunz, J.B.; Neu-Yilik, G.; Breit, S.; Viegas, M.H.; Hentze, M.W.; Kulozik, A.E. Exon-Junction complex components specify distinct routes of nonsense-mediated mRNA decay with differential cofactor requirements. Mol. Cell 2005, 20, 65–75. [Google Scholar] [CrossRef]
- Viegas, M.H.; Gehring, N.H.; Breit, S.; Hentze, M.W.; Kulozik, A.E. The abundance of RNPS1, a protein component of the exon junction complex, can determine the variability in efficiency of the Nonsense Mediated Decay pathway. Nucleic Acids Res. 2007, 35, 4542–4551. [Google Scholar] [CrossRef] [Green Version]
- Peixeiro, I.; Inácio, Â.; Barbosa, C.; Silva, A.L.; Liebhaber, S.A.; Romão, L. Interaction of PABPC1 with the translation initiation complex is critical to the NMD resistance of AUG-proximal nonsense mutations. Nucleic Acids Res. 2012, 40, 1160–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, J.-M.; Srour, N.; Delpy, L. The Yin and Yang of RNA surveillance in B lymphocytes and antibody-Secreting plasma cells. BMB Rep. 2019. [Google Scholar] [CrossRef] [Green Version]
- Mostoslavsky, R.; Alt, F.W.; Rajewsky, K. The lingering enigma of the allelic exclusion mechanism. Cell 2004, 118, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Jung, D.; Giallourakis, C.; Mostoslavsky, R.; Alt, F.W. Mechanism and control of V(D)J recombination at the immunoglobulin heavy chain locus. Annu. Rev. Immunol. 2006, 24, 541–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belgrader, P.; Cheng, J.; Zhou, X.; Stephenson, L.S.; Maquat, L.E. Mammalian nonsense codons can be cis effectors of nuclear mRNA half-Life. Mol. Cell. Biol. 1994, 14, 8219–8228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, M.S.; Doskow, J.; Morris, P.; Li, S.; Nhim, R.P.; Sandstedt, S.; Wilkinson, M.F. A regulatory mechanism that detects premature nonsense codons in T-Cell receptor transcripts in vivo is reversed by protein synthesis inhibitors in vitro. J. Biol. Chem. 1995, 270, 28995–29003. [Google Scholar] [CrossRef] [Green Version]
- Gudikote, J.P.; Wilkinson, M.F. T-Cell receptor sequences that elicit strong down-Regulation of premature termination codon-Bearing transcripts. EMBO J. 2002, 21, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Gudikote, J.P.; Imam, J.S.; Garcia, R.F.; Wilkinson, M.F. RNA splicing promotes translation and RNA surveillance. Nat. Struct. Mol. Biol. 2005, 12, 801–809. [Google Scholar] [CrossRef]
- Wang, J.; Gudikote, J.P.; Olivas, O.R.; Wilkinson, M.F. Boundary-Independent polar nonsense-Mediated decay. EMBO Rep. 2002, 3, 274–279. [Google Scholar] [CrossRef] [Green Version]
- Frischmeyer-Guerrerio, P.A.; Montgomery, R.A.; Warren, D.S.; Cooke, S.K.; Lutz, J.; Sonnenday, C.J.; Guerrerio, A.L.; Dietz, H.C. Perturbation of thymocyte development in nonsense-mediated decay (NMD)-deficient mice. Proc. Natl. Acad. Sci. USA 2011, 108, 10638–10643. [Google Scholar] [CrossRef] [Green Version]
- Weischenfeldt, J.; Damgaard, I.; Bryder, D.; Theilgaard-Mönch, K.; Thoren, L.A.; Nielsen, F.C.; Jacobsen, S.E.; Nerlov, C.; Porse, B. NMD is essential for hematopoietic stem and progenitor cells and for eliminating by-products of programmed DNA rearrangements. Genes Dev. 2008, 22, 1381–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, B.; Potash, M.J.; Köhler, G. Consequences of frameshift mutations at the immunoglobulin heavy chain locus of the mouse. EMBO J. 1985, 4, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Bühler, M.; Paillusson, A.; Mühlemann, O. Efficient downregulation of immunoglobulin mu mRNA with premature translation-Termination codons requires the 5′-half of the VDJ exon. Nucleic Acids Res. 2004, 32, 3304–3315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chemin, G.; Tinguely, A.; Sirac, C.; Lechouane, F.; Duchez, S.; Cogné, M.; Delpy, L. Multiple RNA surveillance mechanisms cooperate to reduce the amount of nonfunctional Ig kappa transcripts. J. Immunol. 2010, 184, 5009–5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delpy, L.; Sirac, C.; Magnoux, E.; Duchez, S.; Cogné, M. RNA surveillance down-Regulates expression of nonfunctional kappa alleles and detects premature termination within the last kappa exon. Proc. Natl. Acad. Sci. USA 2004, 101, 7375–7380. [Google Scholar] [CrossRef] [Green Version]
- Jäck, H.M.; Berg, J.; Wabl, M. Translation affects immunoglobulin mRNA stability. Eur. J. Immunol. 1989, 19, 843–847. [Google Scholar] [CrossRef]
- Tinguely, A.; Chemin, G.; Péron, S.; Sirac, C.; Reynaud, S.; Cogné, M.; Delpy, L. Cross talk between immunoglobulin heavy-Chain transcription and RNA surveillance during B cell development. Mol. Cell. Biol. 2012, 32, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Casola, S.; Otipoby, K.L.; Alimzhanov, M.; Humme, S.; Uyttersprot, N.; Kutok, J.L.; Carroll, M.C.; Rajewsky, K. B cell receptor signal strength determines B cell fate. Nat. Immunol. 2004, 5, 317–327. [Google Scholar] [CrossRef]
- Hentze, M.W.; Kulozik, A.E. A perfect message: RNA surveillance and nonsense-mediated decay. Cell 1999, 96, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Mendell, J.T.; ap Rhys, C.M.J.; Dietz, H.C. Separable roles for rent1/hUpf1 in altered splicing and decay of nonsense transcripts. Science 2002, 298, 419–422. [Google Scholar] [CrossRef]
- Wang, J.; Hamilton, J.I.; Carter, M.S.; Li, S.; Wilkinson, M.F. Alternatively spliced TCR mRNA induced by disruption of reading frame. Science 2002, 297, 108–110. [Google Scholar] [CrossRef]
- Bühler, M.; Mohn, F.; Stalder, L.; Mühlemann, O. Transcriptional silencing of nonsense codon-containing immunoglobulin minigenes. Mol. Cell 2005, 18, 307–317. [Google Scholar] [CrossRef]
- Chang, Y.-F.; Chan, W.-K.; Imam, J.S.; Wilkinson, M.F. Alternatively spliced T-Cell receptor transcripts are up-Regulated in response to disruption of either splicing elements or reading frame. J. Biol. Chem. 2007, 282, 29738–29747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohn, F.; Bühler, M.; Mühlemann, O. Nonsense-Associated alternative splicing of T-Cell receptor beta genes: No evidence for frame dependence. RNA 2005, 11, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chang, Y.F.; Hamilton, J.I.; Wilkinson, M.F. Nonsense-Associated altered splicing: A frame-Dependent response distinct from nonsense-Mediated decay. Mol. Cell 2002, 10, 951–957. [Google Scholar] [CrossRef]
- Srour, N.; Chemin, G.; Tinguely, A.; Ashi, M.O.; Oruc, Z.; Péron, S.; Sirac, C.; Cogné, M.; Delpy, L. A plasma cell differentiation quality control ablates B cell clones with biallelic Ig rearrangements and truncated Ig production. J. Exp. Med. 2016, 213, 109–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maquat, L.E. NASty effects on fibrillin pre-mRNA splicing: Another case of ESE does it, but proposals for translation-dependent splice site choice live on. Genes Dev. 2002, 16, 1743–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, C.R. The association of nonsense codons with exon skipping. Mutat. Res. 1998, 411, 87–117. [Google Scholar] [CrossRef]
- Cartegni, L.; Krainer, A.R. Disruption of an SF2/ASF-Dependent exonic splicing enhancer in SMN2 causes spinal muscular atrophy in the absence of SMN1. Nat. Genet. 2002, 30, 377–384. [Google Scholar] [CrossRef]
- Cartegni, L.; Chew, S.L.; Krainer, A.R. Listening to silence and understanding nonsense: Exonic mutations that affect splicing. Nat. Rev. Genet. 2002, 3, 285–298. [Google Scholar] [CrossRef]
- Ashi, M.O.; Srour, N.; Lambert, J.-M.; Marchalot, A.; Martin, O.; Le Noir, S.; Pinaud, E.; Ayala, M.V.; Sirac, C.; Saulière, J.; et al. Physiological and druggable skipping of immunoglobulin variable exons in plasma cells. Cell. Mol. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nogués, G.; Kadener, S.; Cramer, P.; de la Mata, M.; Fededa, J.P.; Blaustein, M.; Srebrow, A.; Kornblihtt, A.R. Control of alternative pre-mRNA splicing by RNA Pol II elongation: Faster is not always better. IUBMB Life 2003, 55, 235–241. [Google Scholar] [PubMed]
- Shukla, S.; Oberdoerffer, S. Co-Transcriptional regulation of alternative pre-mRNA splicing. Biochim. Biophys. Acta 2012, 1819, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-K.; Xiang, Y.; Feng, X.; Garrard, W.T. Pronounced cohabitation of active immunoglobulin genes from three different chromosomes in transcription factories during maximal antibody synthesis. Genes Dev. 2014, 28, 1159–1164. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambert, J.-M.; Ashi, M.O.; Srour, N.; Delpy, L.; Saulière, J. Mechanisms and Regulation of Nonsense-Mediated mRNA Decay and Nonsense-Associated Altered Splicing in Lymphocytes. Int. J. Mol. Sci. 2020, 21, 1335. https://doi.org/10.3390/ijms21041335
Lambert J-M, Ashi MO, Srour N, Delpy L, Saulière J. Mechanisms and Regulation of Nonsense-Mediated mRNA Decay and Nonsense-Associated Altered Splicing in Lymphocytes. International Journal of Molecular Sciences. 2020; 21(4):1335. https://doi.org/10.3390/ijms21041335
Chicago/Turabian StyleLambert, Jean-Marie, Mohamad Omar Ashi, Nivine Srour, Laurent Delpy, and Jérôme Saulière. 2020. "Mechanisms and Regulation of Nonsense-Mediated mRNA Decay and Nonsense-Associated Altered Splicing in Lymphocytes" International Journal of Molecular Sciences 21, no. 4: 1335. https://doi.org/10.3390/ijms21041335
APA StyleLambert, J. -M., Ashi, M. O., Srour, N., Delpy, L., & Saulière, J. (2020). Mechanisms and Regulation of Nonsense-Mediated mRNA Decay and Nonsense-Associated Altered Splicing in Lymphocytes. International Journal of Molecular Sciences, 21(4), 1335. https://doi.org/10.3390/ijms21041335