Absence of Dx2 at Glu-D1 Locus Weakens Gluten Quality Potentially Regulated by Expression of Nitrogen Metabolism Enzymes and Glutenin-Related Genes in Wheat


Abstract
:1. Introduction
2. Results
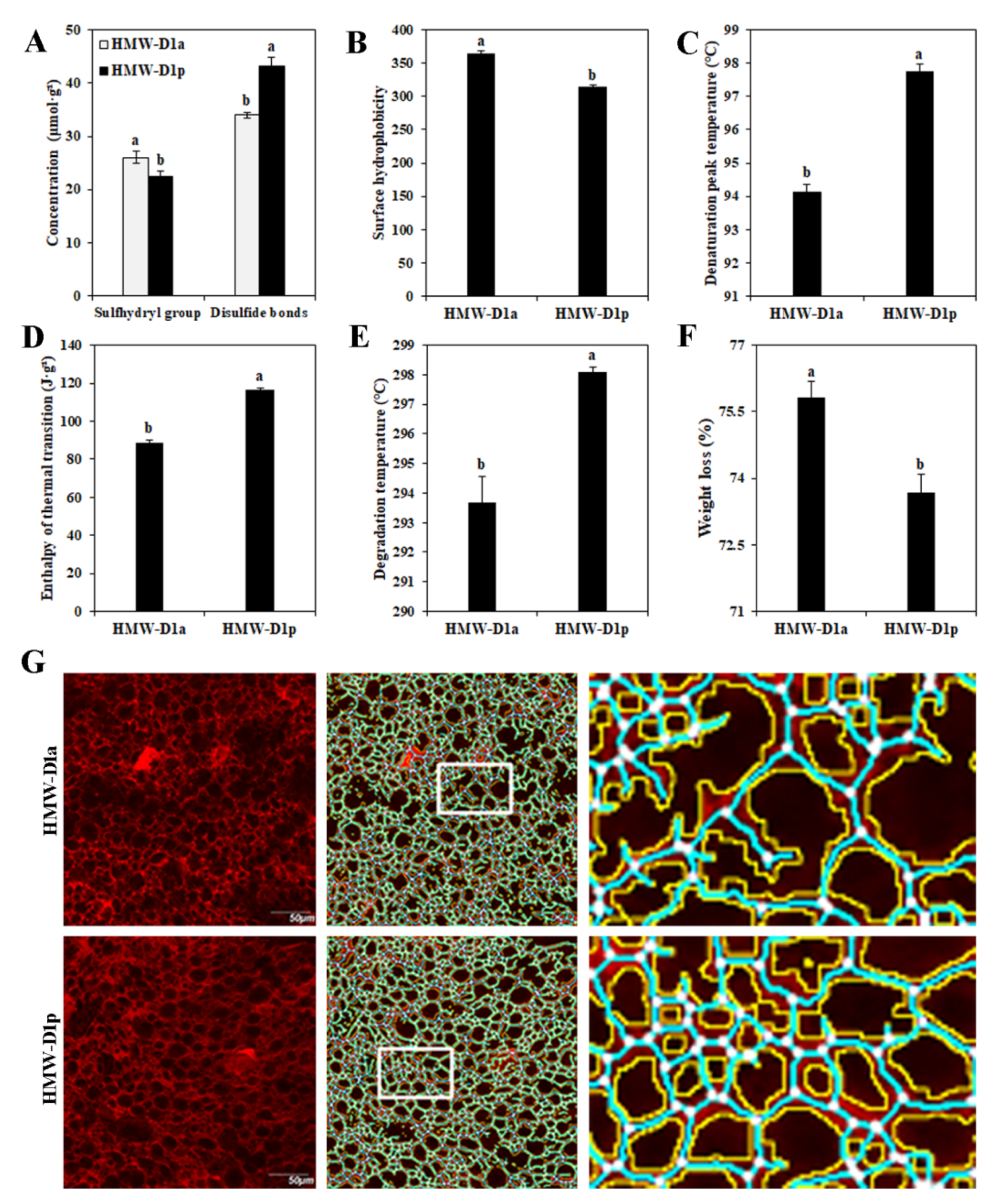
2.1. Analysis of Structural‒Thermal Properties of Gluten
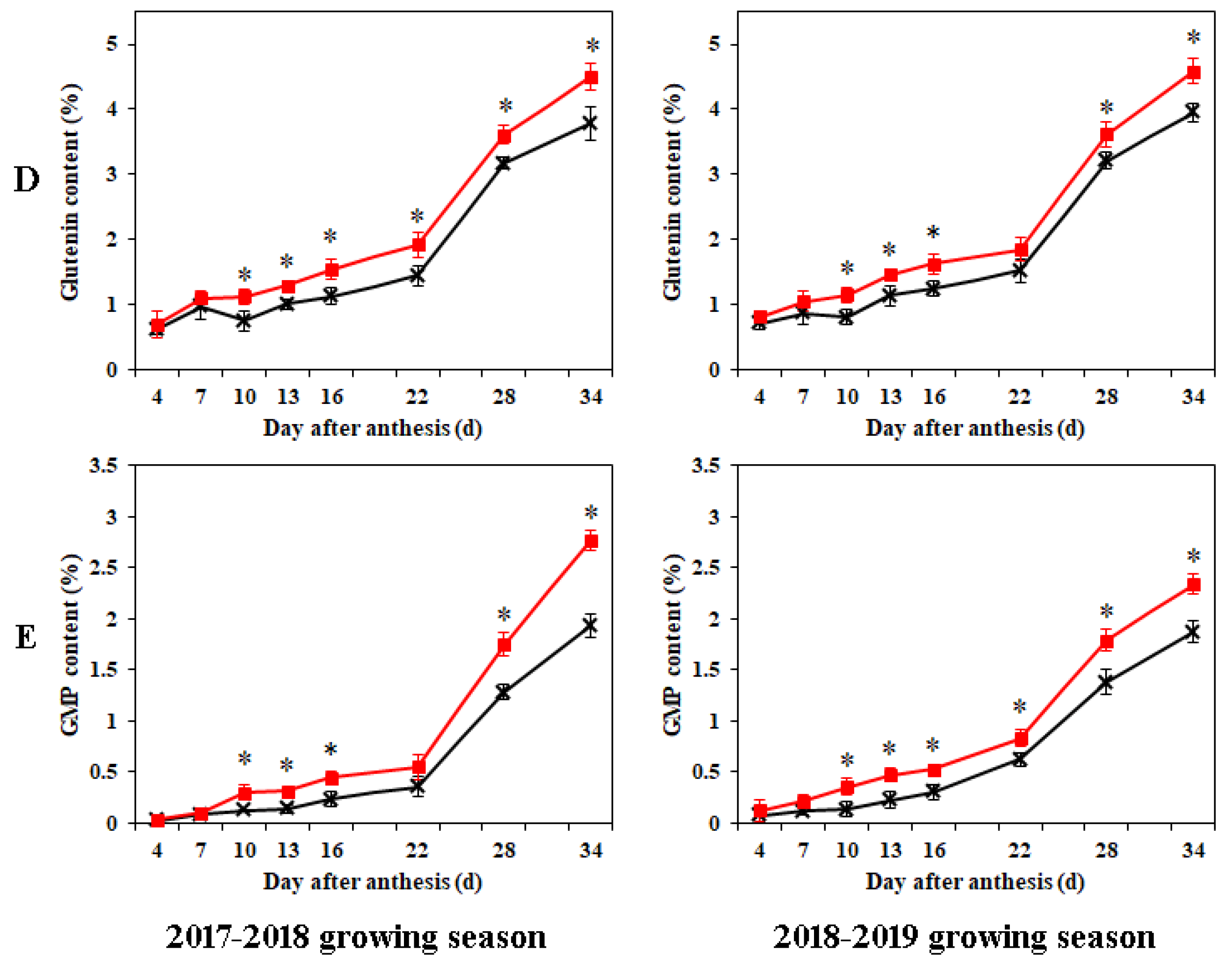
2.2. Dynamic Accumulations of Protein Fractions in Grains during Grain Development
2.3. Accumulation of PBs in Endosperm during Grain Development
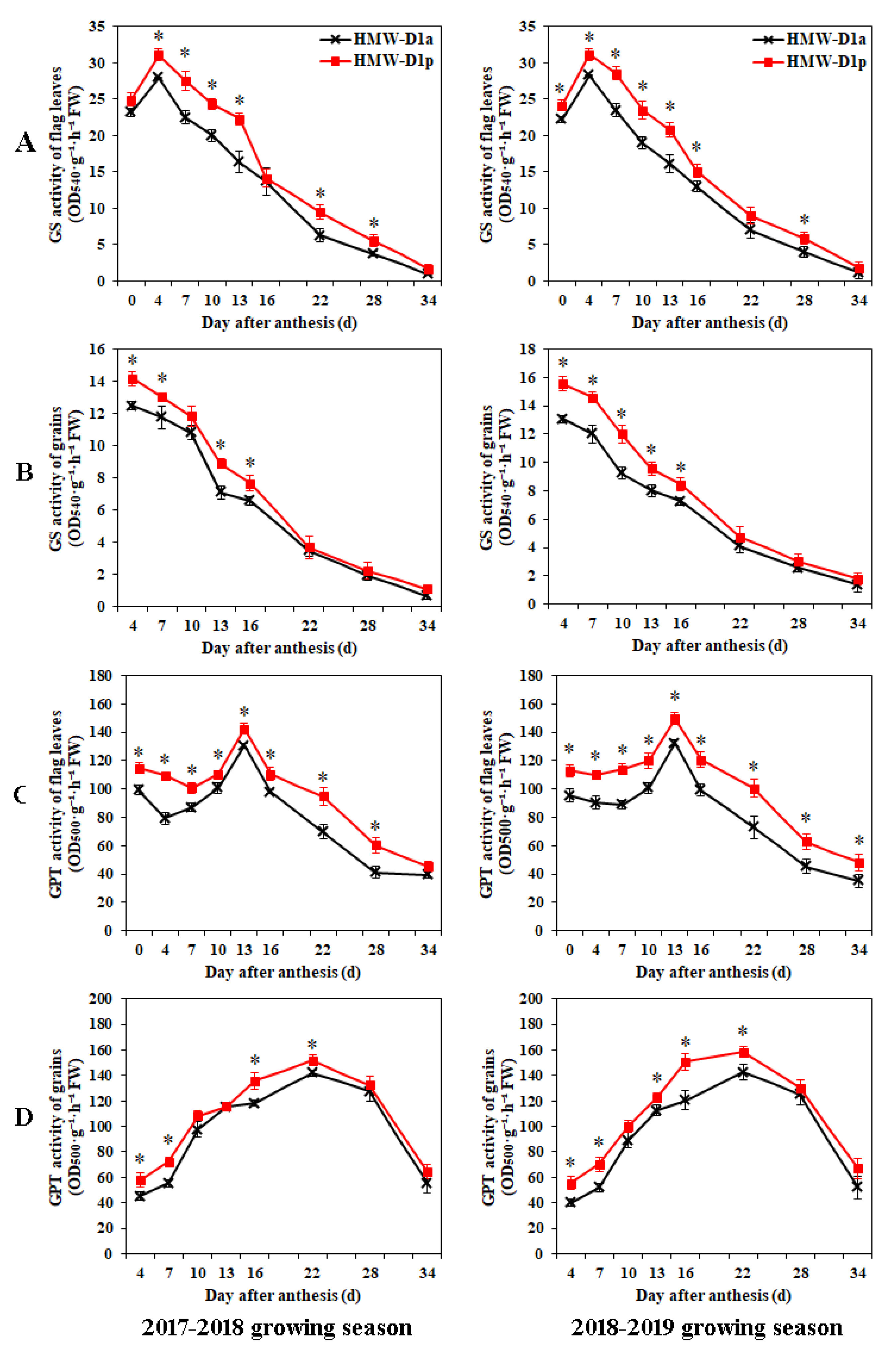
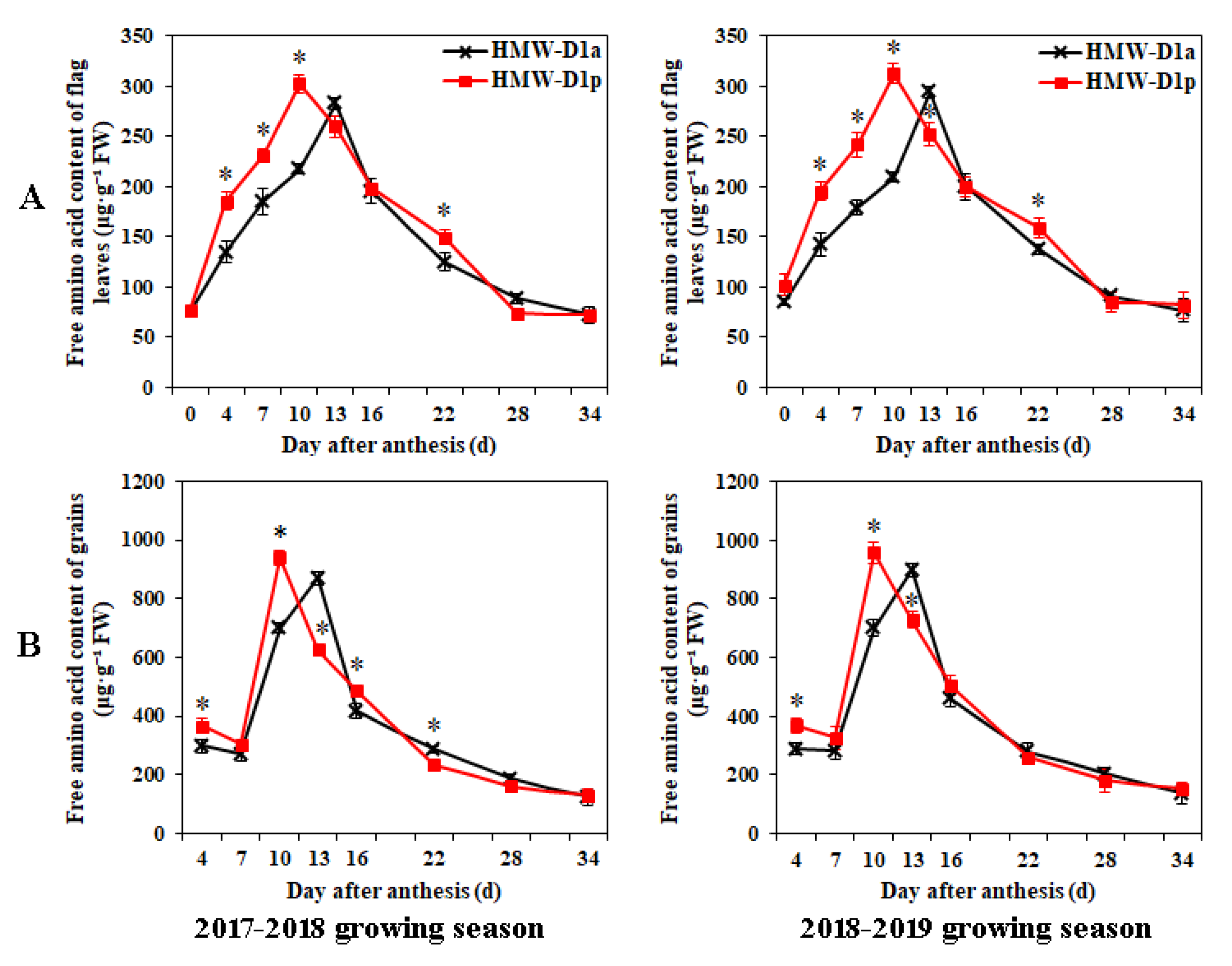
2.4. Changes in GS and GPT Activities and Levels of Amino Acids in Flag Leaves and Grains during Grain Development
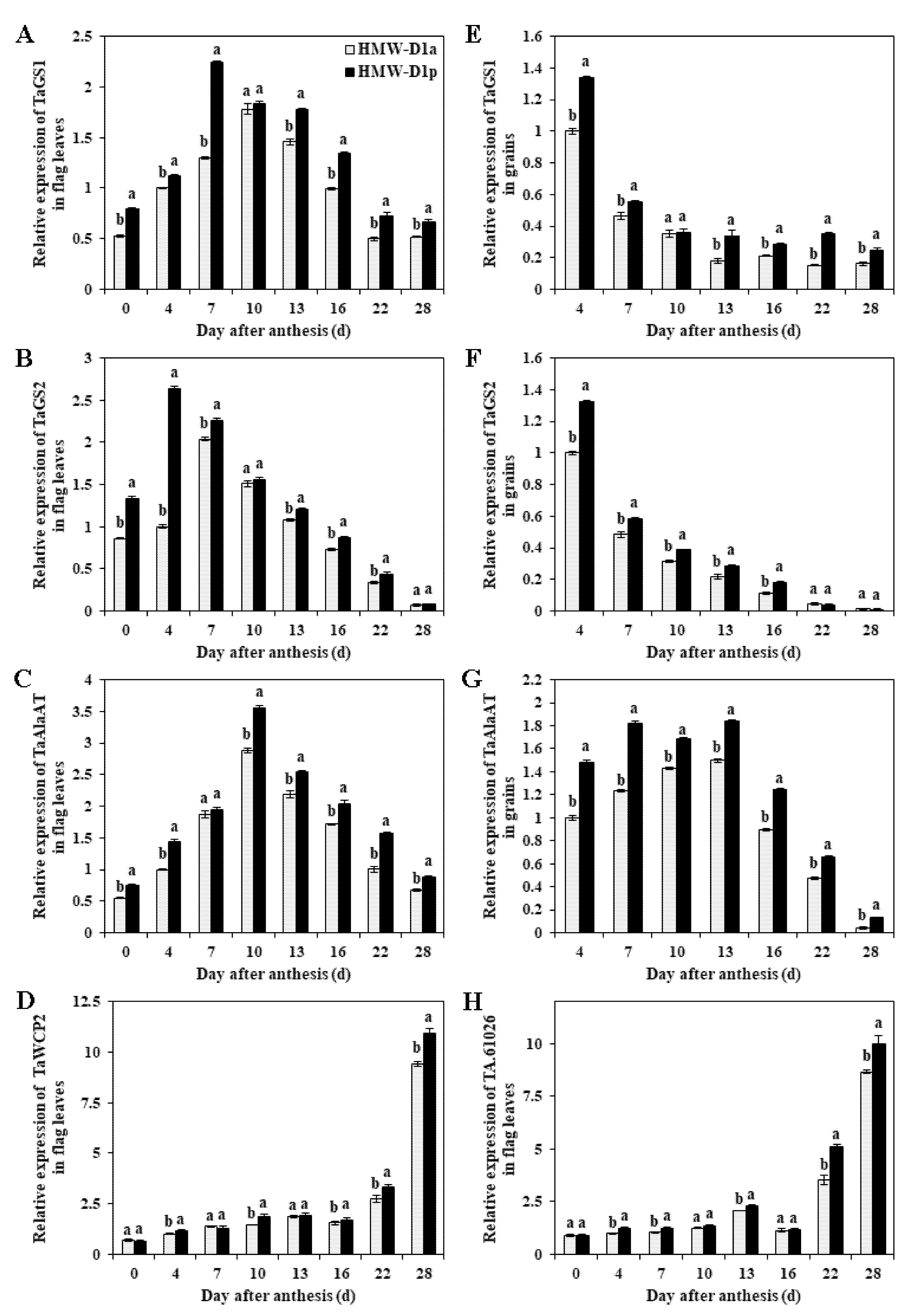
2.5. Expression of Genes Related to Nitrogen Metabolism during Grain Development
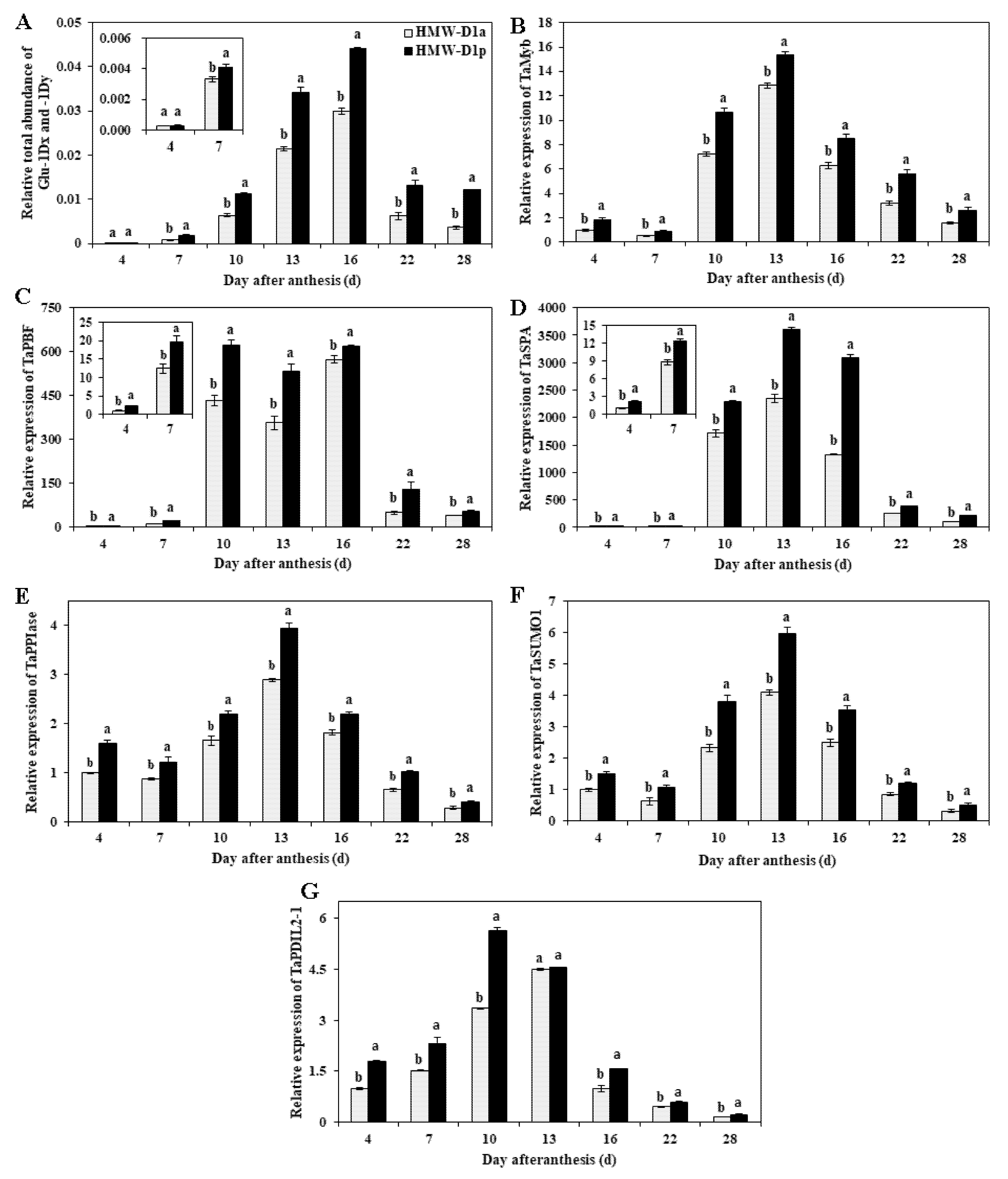
2.6. Transcription Levels of Genes Related to Glutenin Synthesis and Polymerization during Grain Development
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Determination of Structural‒Thermal Properties of Gluten
4.3. Determination of Protein Compositions of Grains during Grain Development
4.4. Cytological Analysis of Endosperm
4.5. Activities of Enzymes Related to Protein Synthesis and Amino Acid Levels in Grains and Flag Leaves during Grain Development
4.6. Gene Expression Analysis during Grain Development
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AlaAT | Alanine aminotransferase |
| DAA | Days after anthesis |
| FW | Fresh weight |
| GAMyb | Gibberellin-response myeloblastosis |
| GMP | Glutenin macropolymer |
| GPT | Glutamate pyruvate transaminase |
| GS | Glutamine synthetase |
| HMW-GS | High-molecular-weight glutenin subunit |
| LMW-GS | Low molecular weight subunit |
| NIL | Near-isogenic line |
| PBF | Prolamin-box binding factor |
| PB | Protein body |
| PDI | Protein disulfide isomerase |
| PDIL2-1 | Protein disulfide isomerase-like 2-1 |
| PPIase | Peptidyl-prolyl cis-trans isomerase |
| SPA | Storage protein activator |
| SUMO1 | Small ubiquitin-related modifier 1 |
| TF | Transcription factor |
References
- Ma, C.Y.; Zhou, J.W.; Chen, G.X.; Bian, Y.W.; Lv, D.W.; Li, X.H.; Wang, Z.M.; Yan, Y.M. iTRAQ-based quantitative proteome and phosphoprotein characterization reveals the central metabolism changes involved in wheat grain development. BMC Genom. 2014, 15, 1029–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goesaert, H.; Brijs, K.; Veraverbeke, W.S.; Courtin, C.M.; Gebruers, K.; Delcour, J.A. Wheat flour constituents: How they impact bread quality, and how to impact their functionality. Trends Food Sci. Technol. 2005, 16, 12–30. [Google Scholar] [CrossRef]
- Plessis, A.; Ravel, C.; Bordes, J.; Balfourier, F.; Martre, P. Association study of wheat grain protein composition reveals that gliadin and glutenin composition are trans-regulated by different chromosome regions. J. Exp. Bot. 2013, 64, 3627–3644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Halford, N.G.; Tatham, A.S.; Popineau, Y.; Belton, P.S. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv. Food Nutr. Res. 2003, 45, 219–302. [Google Scholar] [PubMed]
- Galili, G.; Feldman, M. Genetic control of endosperm proteins in wheat. Theor. Appl. Genet. 1985, 69, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Halford, N.G.; Lafiandra, D. Genetics of wheat gluten proteins. Adv. Genet. 2003, 49, 111–184. [Google Scholar] [PubMed]
- Gao, X.; Liu, T.H.; Ding, M.Y.; Wang, J.; Li, C.L.; Wang, Z.H.; Li, X.J. Effects of HMW-GS Ax1 or Dx2 absence on the glutenin polymerization and gluten micro structure of wheat (Triticum aestivum L.). Food Chem. 2018, 240, 626–633. [Google Scholar] [CrossRef]
- Li, X.J.; Liu, T.H.; Song, L.J.; Zhang, H.; Li, L.Q.; Gao, X. Influence of high-molecular-weight glutenin subunit composition at Glu-A1 and Glu-D1 loci on secondary and micro structures of gluten in wheat (Triticum aestivum L.). Food Chem. 2016, 213, 728–734. [Google Scholar] [CrossRef]
- Guo, W.W.; Yang, H.; Liu, Y.Q.; Gao, Y.J.; Ni, Z.F.; Peng, H.R.; Xin, M.M.; Hu, Z.R.; Sun, Q.X.; Yao, Y.Y. The wheat transcription factor TaGAMyb recruits histone acetyltransferase and activates the expression of a high-molecular-weight glutenin subunit gene. Plant J. 2015, 84, 347–359. [Google Scholar] [CrossRef]
- Zhong, Y.X.; Xu, D.C.; Hebelstrup, K.H.; Yang, D.L.; Cai, J.; Wang, X.; Zhou, Q.; Cao, W.X.; Dai, T.B.; Jiang, D. Nitrogen topdressing timing modifies free amino acids profiles and storage protein gene expression in wheat grain. BMC Plant Biol. 2018, 18, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Ravel, C.; Fiquet, S.; Boudet, J.; Dardevet, M.; Vincent, J.; Merlino, M.; Michard, R.; Martre, P. Conserved cis-regulatory modules in promoters of genes encoding wheat high-molecular-weight glutenin subunits. Front. Plant Sci. 2014, 5, 621–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.S.; Liu, X.Y.; Wei, Q.H.; Liu, J.N.; Yang, T.X.; Jia, L.Y.; Wang, Y.S.; Yang, G.X.; He, G.Y. Functional Characterization of TaFUSCA3, a B3-Superfamily Transcription Factor Gene in the Wheat. Front. Plant Sci. 2017, 8, 1133–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirel, B.; Gouis, J.L.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhao, W.; Yang, J.; Oosterhuis, D.M.; Loka, D.A.; Zhou, Z. Relationship between potassium fertilization and nitrogen metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll during the boll development stage. Plant Physiol. Biochem. 2016, 101, 113–123. [Google Scholar] [CrossRef]
- Shrawat, A.K.; Carroll, R.T.; Depauw, M.; Taylor, G.J.; Good, A.G. Genetic engineering of improved nitrogen use efficiency in rice by the tissue-specific expression of alanine aminotransferase. Plant Biotechnol. J. 2008, 6, 722–732. [Google Scholar] [CrossRef]
- Weber, H.; Sreenivasulu, N.; Weschke, W. Molecular physiology of seed maturation and seed storage protein biosynthesis. In Plant Developmental Biology—Biotechnological Perspectives; Springer: Heidelberg, Germany, 2010; pp. 83–104. [Google Scholar]
- Levanony, H.; Rubin, R.; Altschuler, Y.; Galili, G. Evidence for a novel route of wheat storage proteins to vacuoles. J. Cell Biol. 1992, 119, 1117–1128. [Google Scholar] [CrossRef]
- Baneyx, F.; Mujacic, M. Recombinant protein folding and misfolding in Escherichia coli. Nat. Biotechnol. 2004, 22, 1399–1408. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, W.Y.; Park, H.C.; Lee, S.Y.; Bohnert, H.J.; Yun, D.J. SUMO and SUMOylation in plants. Mol. Cells 2011, 32, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.T.; Islam, S.; She, M.Y.; Diepeveen, D.; Zhang, Y.J.; Tang, G.X.; Zhang, J.J.; Juhasz, A.; Yang, R.C.; Ma, W.J. Wheat grain protein accumulation and polymerization mechanisms driven by nitrogen fertilization. Plant J. 2018, 96, 1160–1177. [Google Scholar] [CrossRef]
- Kimura, S.; Higashino, Y.; Kitao, Y.; Masuda, T.; Urade, R. Expression and characterization of protein disulfide isomerase family proteins in bread wheat. BMC Plant Biol. 2015, 15, 73–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.X.; Yang, M.T.; Cai, J.; Wang, X.; Zhou, Q.; Cao, W.X.; Dai, T.B.; Jiang, D. Nitrogen topdressing timing influences the spatial distribution patterns of protein components and quality traits of flours from different pearling fractions of wheat (Triticum aestivum L.) grains. Field Crops Res. 2018, 216, 120–128. [Google Scholar] [CrossRef]
- Liu, T.H.; Gao, X.; Li, L.Q.; Lv, Q.; Du, D.F.; Cheng, X.; Zhao, Y.; Liu, Y.; Li, X.J. Effects of HMW-GS at Glu-B1 locus on the polymerization of glutenin during grain development and on the secondary and micro-structures of gluten in wheat (Triticum aestivum L.). J. Cereal Sci. 2016, 72, 101–107. [Google Scholar] [CrossRef]
- Zhu, J.; Li, L.Q.; Zhao, L.Y.; Song, L.J.; Li, X.J. Effects of freeze–thaw cycles on the structural and thermal properties of wheat gluten with variations in the high molecular weight glutenin subunit at the Glu-B1 locus. J. Cereal Sci. 2019, 87, 266–272. [Google Scholar] [CrossRef]
- Yu, X.R.; Chen, X.Y.; Wang, L.L.; Yang, Y.; Zhu, X.W.; Shao, S.S.; Cui, W.X.; Xiong, F. Novel insights into the effect of nitrogen on storage protein biosynthesis and protein body development in wheat caryopsis. J. Exp. Bot. 2017, 68, 2259–2275. [Google Scholar] [CrossRef] [PubMed]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.W.; Ma, D.Y.; Ma, G.; Wang, C.Y.; Xie, X.D.; Kang, G.A. Responses of glutamine synthetase activity and gene expression to nitrogen levels in winter wheat cultivars with different grain protein content. J. Cereal Sci. 2017, 74, 187–193. [Google Scholar] [CrossRef]
- Habash, D.Z.; Swarbreck, S.; Schondelmaier, J.; Weyen, J.; Quarrie, S. The genetics of nitrogen use in hexaploid wheat: N utilisation, development and yield. Theor. Appl. Genet. 2007, 114, 403–419. [Google Scholar] [CrossRef]
- Gadaleta, A.; Nigro, D.; Giancaspro, A.; Blanco, A. The glutamine synthetase (GS2) genes in relation to grain protein content of durum wheat. Funct. Integr. Genomic. 2011, 11, 665–670. [Google Scholar] [CrossRef]
- Tosi, P. Trafficking and deposition of prolamins in wheat. J. Cereal Sci. 2002, 56, 81–90. [Google Scholar] [CrossRef]
- Grimwade, B.; Tatham, A.S.; Freedman, R.B.; Shewry, P.R.; Napier, J.A. Comparison of the expression patterns of genes coding for wheat gluten proteins and proteins involved in the secretory pathway in developing caryopses of wheat. Plant Mol. Biol. 1996, 30, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Jalal, D.; Chalissery, J.; Hassan, A.H. Genome maintenance in Saccharomyces cerevisiae: The role of SUMO and SUMO-targeted ubiquitin ligases. Nucleic. Acids. Res. 2017, 45, 2242–2261. [Google Scholar] [PubMed] [Green Version]
- Morel, M.H.; Redl, A.; Guilbert, S. Mechanism of heat and shear mediated aggregation of wheat gluten protein upon mixing. Biomacromolecules 2002, 3, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Amagliani, L.; O’Regan, J.; Kelly, A.L.; O’Mahony, J.A. Composition and protein profile analysis of rice protein ingredients. J. Food Compos. Anal. 2017, 59, 18–26. [Google Scholar] [CrossRef]
- Weegels, P.L.; Hamer, R.J.; Schofield, J.D. Depolymerisation and repolymerisation of wheat glutenin during dough processing. II. Changes in composition. J. Cereal Sci. 1997, 25, 155–163. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Protein Area (× 104 μm2) | Protein Junctions (× 102) | Junction Density (×10−3) | Total Protein Length (× 103 µm) | Endpoints (× 102) | Lacunarity (×10−2) | Branching Rate (×10−3) | Endpoint Rate (×10−3) |
|---|---|---|---|---|---|---|---|---|
| HMW-D1a | 11.63 ± 0.15b | 11.74 ± 0.15b | 4.50 ± 0.05b | 21.88 ± 0.25b | 3.60 ± 0.21a | 4.81 ± 0.07b | 10.09 ± 0.04b | 3.09 ± 0.22a |
| HMW-D1p | 12.79 ± 0.05a | 13.64 ± 0.19a | 5.07 ± 0.04a | 23.77 ± 0.24a | 2.64 ± 0.19b | 5.76 ± 0.06a | 10.66 ± 0.15a | 2.06 ± 0.16b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, L.; Li, L.; Zhao, L.; Liu, Z.; Xie, T.; Li, X. Absence of Dx2 at Glu-D1 Locus Weakens Gluten Quality Potentially Regulated by Expression of Nitrogen Metabolism Enzymes and Glutenin-Related Genes in Wheat. Int. J. Mol. Sci. 2020, 21, 1383. https://doi.org/10.3390/ijms21041383
Song L, Li L, Zhao L, Liu Z, Xie T, Li X. Absence of Dx2 at Glu-D1 Locus Weakens Gluten Quality Potentially Regulated by Expression of Nitrogen Metabolism Enzymes and Glutenin-Related Genes in Wheat. International Journal of Molecular Sciences. 2020; 21(4):1383. https://doi.org/10.3390/ijms21041383
Chicago/Turabian StyleSong, Lijun, Liqun Li, Liye Zhao, Zhenzhen Liu, Tingting Xie, and Xuejun Li. 2020. "Absence of Dx2 at Glu-D1 Locus Weakens Gluten Quality Potentially Regulated by Expression of Nitrogen Metabolism Enzymes and Glutenin-Related Genes in Wheat" International Journal of Molecular Sciences 21, no. 4: 1383. https://doi.org/10.3390/ijms21041383
APA StyleSong, L., Li, L., Zhao, L., Liu, Z., Xie, T., & Li, X. (2020). Absence of Dx2 at Glu-D1 Locus Weakens Gluten Quality Potentially Regulated by Expression of Nitrogen Metabolism Enzymes and Glutenin-Related Genes in Wheat. International Journal of Molecular Sciences, 21(4), 1383. https://doi.org/10.3390/ijms21041383