Oestrogenic Endocrine Disruptors in the Placenta and the Fetus




Abstract
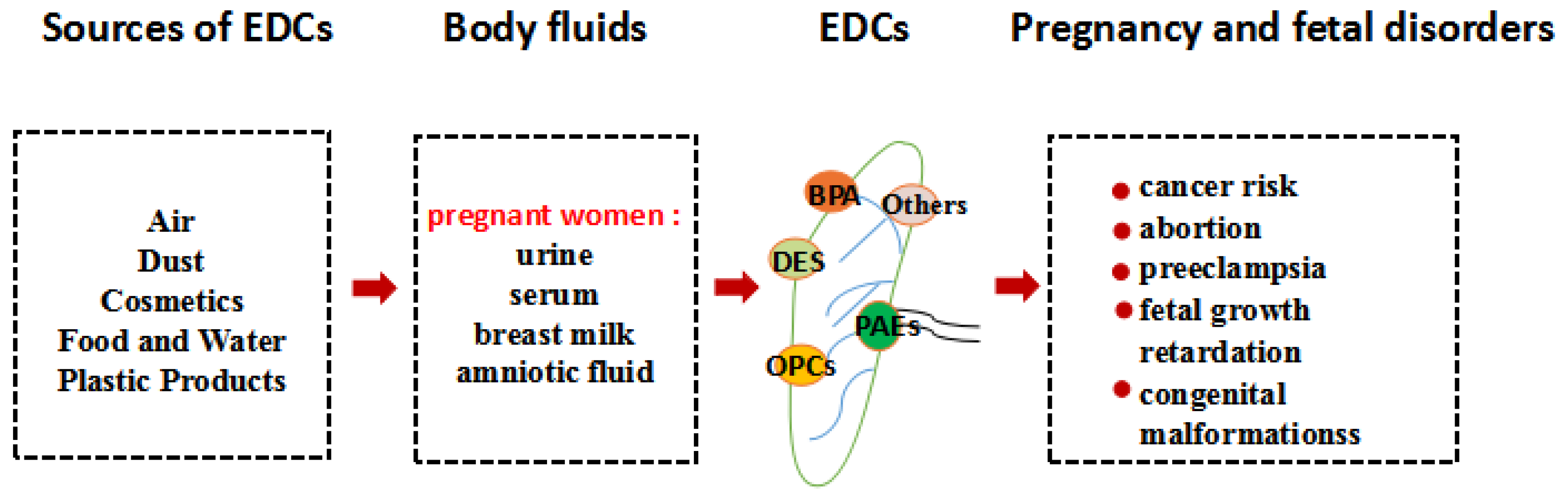
:1. Introduction
2. Detection Methods
3. Endocrine Disrupting Chemicals
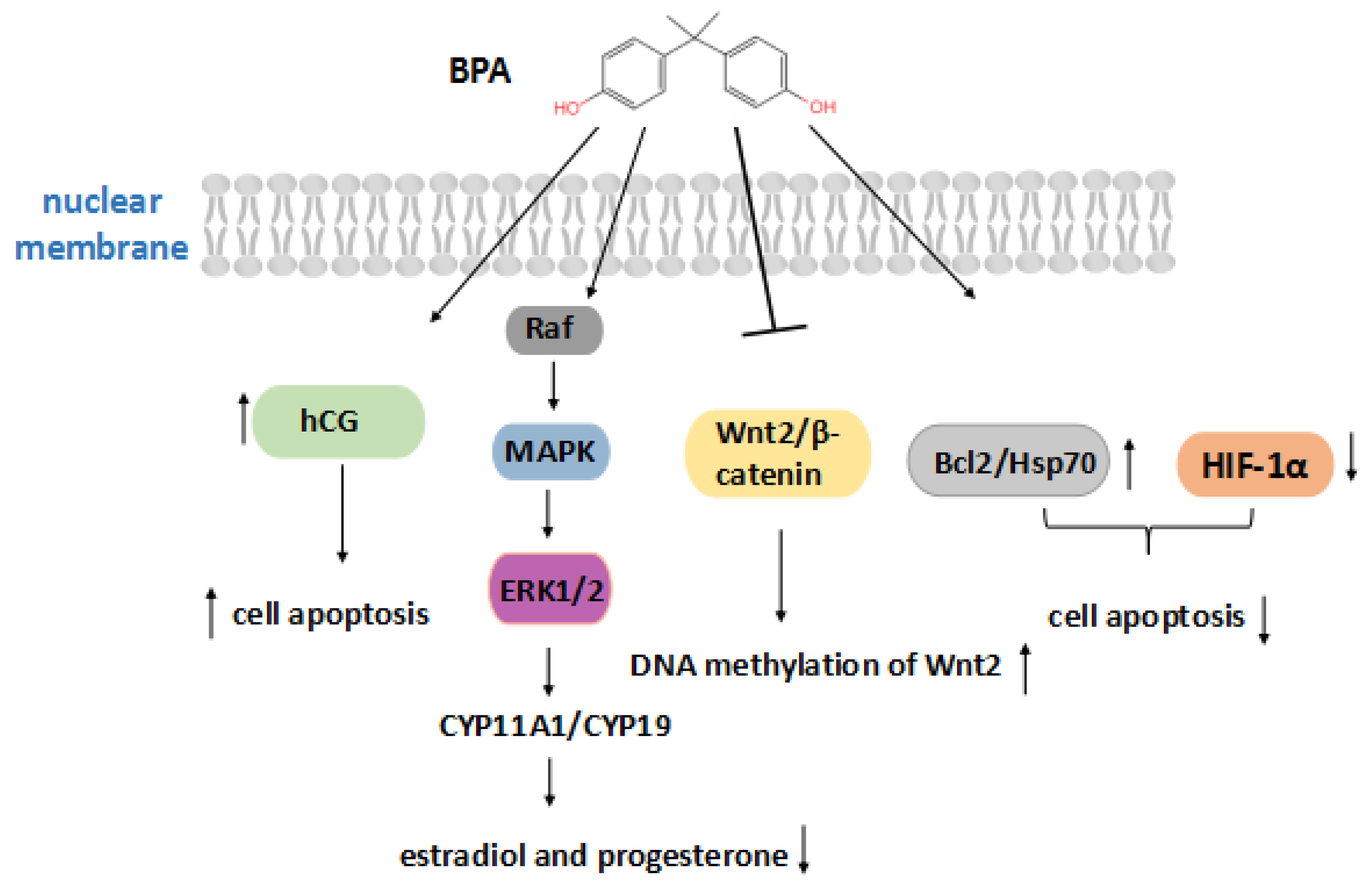
3.1. Bisphenol A
3.2. Organochlorine Pesticides
3.3. Diethylstilbestrol
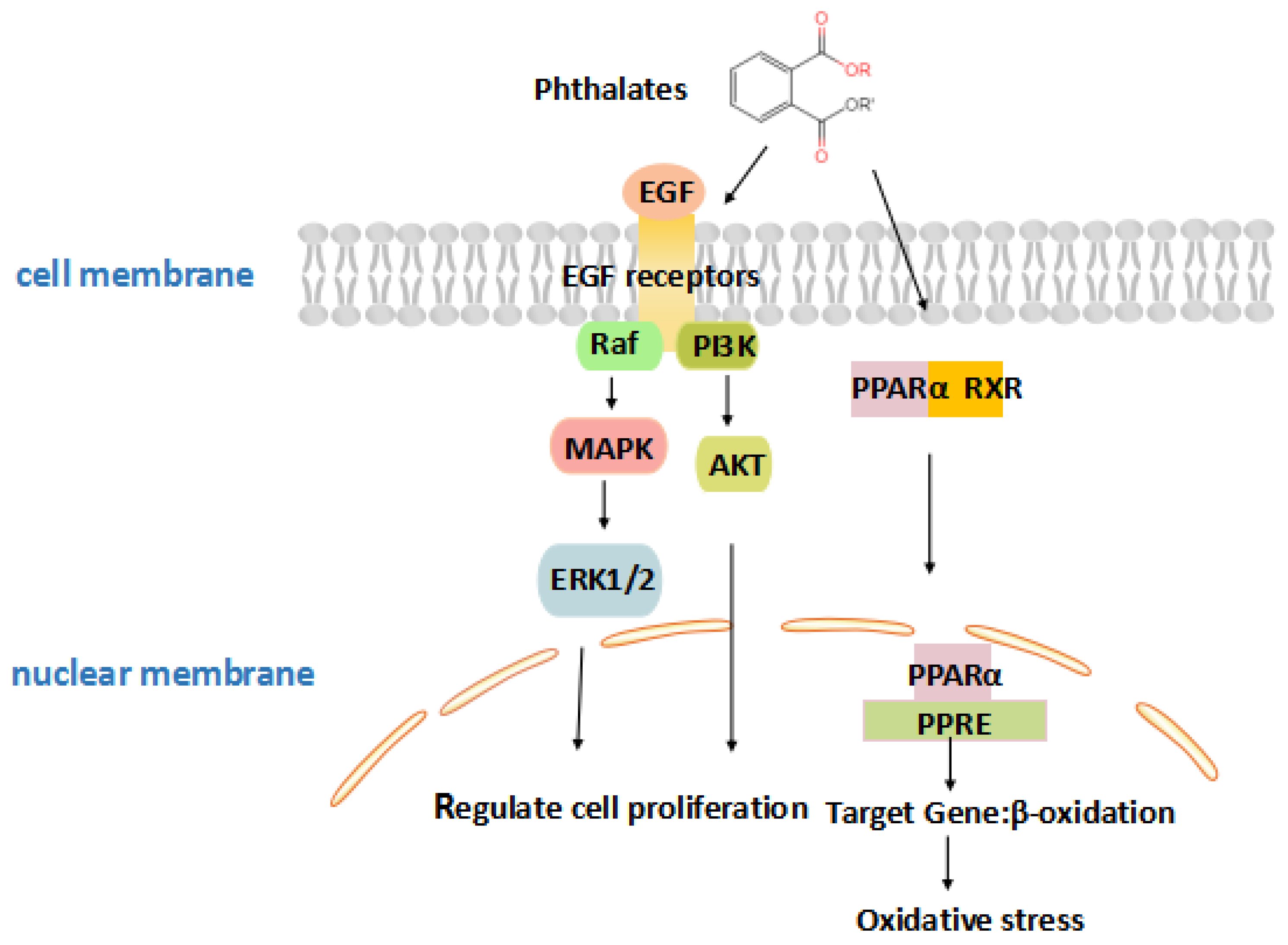
3.4. Phthalates
3.5. Phytoestrogens
4. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Kavlock, R.J.; Daston, G.P.; DeRosa, C.; Fenner-Crisp, P.; Gray, L.E.; Kaattari, S.; Lucier, G.; Luster, M.; Mac, M.J.; Maczka, C.; et al. Research needs for the risk assessment of health and environmental effects of endocrine disruptors: A report of the U.S. EPA-sponsored workshop. Environ. Health Perspect. 1996, 104, 715–740. [Google Scholar]
- Diamanti-Kandarakis, E.; Bourguignon, J.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Monneret, C. What is an endocrine disruptor? CR Biol. 2017, 340, 403–405. [Google Scholar] [CrossRef]
- Commission of the European Communities. Community Strategy for Endocrine Disrupters: A Range of Substances Suspected of Interfering with the Hormone Systems of Humans and Wildlife; COM (1999) 706 Final; Commission of the European Communities: Brussels, Belgium, 1999. [Google Scholar]
- Candidate List of Substances of Very High Concern for Authorisation; European Communities: Brussels, Belgium, 2017.
- Kabir, E.R.; Rahman, M.S.; Rahman, I. A review on endocrine disruptors and their possible impacts on human health. Environ. Toxicol. Pharmacol. 2015, 40, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Pollheimer, J.; Vondra, S.; Baltayeva, J.; Beristain, A.G.; Knofler, M. Regulation of Placental Extravillous Trophoblasts by the Maternal Uterine Environment. Front. Immunol. 2018, 9, 2597. [Google Scholar] [CrossRef] [PubMed]
- Saghian, R.; Bogle, G.; James, J.L.; Clark, A.R. Establishment of maternal blood supply to the placenta: Insights into plugging, unplugging and trophoblast behaviour from an agent-based model. Interface Focus 2019, 9, 20190019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, V.E.; Smith, R.; Giles, W.B.; Clifton, V.L. Endocrine regulation of human fetal growth: The role of the mother, placenta, and fetus. Endocr. Rev. 2006, 27, 141–169. [Google Scholar] [CrossRef]
- Lauritzen, H.B.; Larose, T.L.; Oien, T.; Sandanger, T.M.; Odland, J.O.; van de Bor, M.; Jacobsen, G.W. Maternal serum levels of perfluoroalkyl substances and organochlorines and indices of fetal growth: A Scandinavian case-cohort study. Pediatr. Res. 2017, 81, 33–42. [Google Scholar] [CrossRef]
- Geer, L.A.; Pycke, B.F.G.; Waxenbaum, J.; Sherer, D.M.; Abulafia, O.; Halden, R.U. Association of birth outcomes with fetal exposure to parabens, triclosan and triclocarban in an immigrant population in Brooklyn, New York. J. Hazard. Mater. 2017, 323, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Zhou, Q.; Feng, L.; Wu, J.; Xiong, Y.; Li, X. Maternal serum bisphenol A levels and risk of pre-eclampsia: A nested case-control study. Eur. J. Public Health 2017, 27, 1102–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.T.; Christie, P.; Luo, Y.M.; Teng, Y. Phthalate esters contamination in soil and plants on agricultural land near an electronic waste recycling site. Environ. Geochem. Health 2013, 35, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Michalowicz, J. Bisphenol A--sources, toxicity and biotransformation. Environ. Toxicol. Phar. 2014, 37, 738–758. [Google Scholar] [CrossRef] [PubMed]
- Maitre, L.; Robinson, O.; Martinez, D.; Toledano, M.B.; Ibarluzea, J.; Marina, L.S.; Sunyer, J.; Villanueva, C.M.; Keun, H.C.; Vrijheid, M.; et al. Urine Metabolic Signatures of Multiple Environmental Pollutants in Pregnant Women: An Exposome Approach. Environ. Sci. Technol. 2018, 52, 13469–13480. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, S.; Sood, S.; Showkat, S.; Lite, C.; Chandrasekhar, A.; Vairamani, M.; Barathi, S.; Santosh, W. Detection of phenolic endocrine disrupting chemicals (EDCs) from maternal blood plasma and amniotic fluid in Indian population. Gen. Comp. Endocr. 2017, 241, 100–107. [Google Scholar] [CrossRef]
- Susiarjo, M.; Sasson, I.; Mesaros, C.; Bartolomei, M.S. Bisphenol a exposure disrupts genomic imprinting in the mouse. PLoS Genet. 2013, 9, e1003401. [Google Scholar] [CrossRef] [Green Version]
- Skakkebaek, N.E.; Toppari, J.; Soder, O.; Gordon, C.M.; Divall, S.; Draznin, M. The exposure of fetuses and children to endocrine disrupting chemicals: A European Society for Paediatric Endocrinology (ESPE) and Pediatric Endocrine Society (PES) call to action statement. J. Clin. Endocrinol. Metab. 2011, 96, 3056–3058. [Google Scholar] [CrossRef]
- Perez-Albaladejo, E.; Fernandes, D.; Lacorte, S.; Porte, C. Comparative toxicity, oxidative stress and endocrine disruption potential of plasticizers in JEG-3 human placental cells. Toxicol. Vitr. 2017, 38, 41–48. [Google Scholar] [CrossRef]
- Li, L.; Chen, L.; Meng, X.; Chen, B.; Chen, S.; Zhao, Y.; Zhao, L.; Liang, Y.; Zhang, Y. Exposure levels of environmental endocrine disruptors in mother-newborn pairs in China and their placental transfer characteristics. PLoS ONE 2013, 8, e62526. [Google Scholar] [CrossRef] [Green Version]
- Roncati, L.; Piscioli, F.; Pusiol, T. The endocrine disruptors among the environmental risk factors for stillbirth. Sci. Total Environ. 2016, 563–564, 1086–1087. [Google Scholar] [CrossRef]
- Feeley, M.; Brouwer, A. Health risks to infants from exposure to PCBs, PCDDs and PCDFs. Food Addit. Contam. 2000, 17, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Dusza, H.M.; Janssen, E.; Kanda, R.; Legler, J. Method Development for Effect-Directed Analysis of Endocrine Disrupting Compounds in Human Amniotic Fluid. Environ. Sci. Technol. 2019, 53, 14649–14659. [Google Scholar] [CrossRef] [PubMed]
- Mannetje, A.T.; Coakley, J.; Mueller, J.F.; Harden, F.; Toms, L.; Douwes, J. Partitioning of persistent organic pollutants (POPs) between human serum and breast milk: A literature review. Chemosphere 2012, 89, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, H.; Kranich, S.K.; Jorgensen, N.; Taboureau, O.; Petersen, J.H.; Andersson, A. Temporal variability in urinary phthalate metabolite excretion based on spot, morning, and 24-h urine samples: Considerations for epidemiological studies. Environ. Sci. Technol. 2013, 47, 958–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krais, A.M.; Andersen, C.; Eriksson, A.C.; Johnsson, E.; Nielsen, J.; Pagels, J.; Gudmundsson, A.; Lindh, C.H.; Wierzbicka, A. Excretion of Urinary Metabolites of the Phthalate Esters DEP and DEHP in 16Volunteers after Inhalation and Dermal Exposure. Int. J. Environ. Res. Publ. Health 2018, 15, 2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrell, J.; Foster, W.G.; Kinniburgh, D.W. Phytoestrogens in human pregnancy. Obstet. Gynecol. Int. 2012, 2012, 850313. [Google Scholar] [CrossRef]
- Soucy, N.V.; Parkinson, H.D.; Sochaski, M.A.; Borghoff, S.J. Kinetics of genistein and its conjugated metabolites in pregnant Sprague-Dawleyrats following single and repeated genistein administration. Toxicol. Sci. Off. J. Soc. Toxicol. 2006, 90, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, B.; Thorstensen, E.B.; Ponnampalam, A.P.; Mitchell, M.D. Transplacental transfer and biotransformation of genistein in human placenta. Placenta 2010, 31, 506–511. [Google Scholar] [CrossRef]
- Sun, F.; Kang, L.; Xiang, X.; Li, H.; Luo, X.; Luo, R.; Lu, C.; Peng, X. Recent advances and progress in the detection of bisphenol A. Anal. Bioanal. Chem. 2016, 408, 6913–6927. [Google Scholar] [CrossRef]
- Schonfelder, G.; Wittfoht, W.; Hopp, H.; Talsness, C.E.; Paul, M.; Chahoud, I. Parent bisphenol A accumulation in the human maternal-fetal-placental unit. Environ. Health Perspect. 2002, 110, A703–A707. [Google Scholar] [CrossRef]
- Yamada, H.; Furuta, I.; Kato, E.H.; Kataoka, S.; Usuki, Y.; Kobashi, G.; Sata, F.; Kishi, R.; Fujimoto, S. Maternal serum and amniotic fluid bisphenol A concentrations in the early second trimester. Reprod. Toxicol. 2002, 16, 735–739. [Google Scholar] [CrossRef]
- Ikezuki, Y.; Tsutsumi, O.; Takai, Y.; Kamei, Y.; Taketani, Y. Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure. Hum. Reprod. (Oxf. Engl.) 2002, 17, 2839–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, N.; Surendranath, A.R.; Pathak, J.L.; Yu, S.; Chung, C.Y. Bisphenol A (BPA) the mighty and the mutagenic. Toxicol. Rep. 2018, 5, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Foster, W.; Chan, S.; Platt, L.; Hughes, C. Detection of endocrine disrupting chemicals in samples of second trimester human amniotic fluid. J. Clin. Endocrinol. Metab. 2000, 85, 2954–2957. [Google Scholar] [CrossRef]
- Koepke, R.; Warner, M.; Petreas, M.; Cabria, A.; Danis, R.; Hernandez-Avila, M.; Eskenazi, B. Serum DDT and DDE levels in pregnant women of Chiapas, Mexico. Arch. Environ. Health 2004, 59, 559–565. [Google Scholar] [CrossRef]
- Hines, E.P.; Calafat, A.M.; Silva, M.J.; Mendola, P.; Fenton, S.E. Concentrations of phthalate metabolites in milk, urine, saliva, and Serum of lactating North Carolina women. Environ. Health Perspect. 2009, 117, 86–92. [Google Scholar] [CrossRef]
- Arbuckle, T.E.; Davis, K.; Marro, L.; Fisher, M.; Legrand, M.; LeBlanc, A.; Gaudreau, E.; Foster, W.G.; Choeurng, V.; Fraser, W.D. Phthalate and bisphenol A exposure among pregnant women in Canada--results from the MIREC study. Environ. Int. 2014, 68, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Almeida, C.; Nogueira, J.M.F. Determination of steroid sex hormones in water and urine matrices by stir barsorptive extraction and liquid chromatography with diode array detection. J. Pharm. Biomed. 2006, 41, 1303–1311. [Google Scholar] [CrossRef]
- Foster, W.G.; Chan, S.; Platt, L.; Hughes, C.L.J. Detection of phytoestrogens in samples of second trimester human amniotic fluid. Toxicol. Lett. 2002, 129, 199–205. [Google Scholar] [CrossRef]
- Fleck, S.C.; Churchwell, M.I.; Doerge, D.R.; Teeguarden, J.G. Urine and serum biomonitoring of exposure to environmental estrogens II: Soyisoflavones and zearalenone in pregnant women. Food Chem. Toxicol. Int. J. Publ. Br. Biol. Res. Assoc. 2016, 95, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Geens, T.; Aerts, D.; Berthot, C.; Bourguignon, J.; Goeyens, L.; Lecomte, P.; Maghuin-Rogister, G.; Pironnet, A.; Pussemier, L.; Scippo, M.; et al. A review of dietary and non-dietary exposure to bisphenol-A. Food Chem. Toxicol. 2012, 50, 3725–3740. [Google Scholar] [CrossRef] [PubMed]
- Hartle, J.C.; Navas-Acien, A.; Lawrence, R.S. The consumption of canned food and beverages and urinary Bisphenol A concentrations in NHANES 2003-2008. Environ. Res. 2016, 150, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellahi, M.; Rashid, M.U. The Toxic Effects BPA on Fetuses, Infants, and Children. Intech Open 2017. [Google Scholar]
- Morck, T.J.; Sorda, G.; Bechi, N.; Rasmussen, B.S.; Nielsen, J.B.; Ietta, F.; Rytting, E.; Mathiesen, L.; Paulesu, L. Placental transport and in vitro effects of Bisphenol A. Reprod. Toxicol. 2010, 30, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Ponniah, M.; Billett, E.E.; De Girolamo, L.A. Bisphenol A increases BeWo trophoblast survival in stress-induced paradigms through regulation of oxidative stress and apoptosis. Chem. Res. Toxicol. 2015, 28, 1693–1703. [Google Scholar] [CrossRef]
- Chu, P.; Yang, Z.; Huang, H.; Chang, A.; Cheng, Y.; Wu, G.; Lan, H. Low-dose bisphenol A activates the ERK signaling pathway and attenuates steroidogenic gene expression in human placental cells. Biol. Reprod. 2018, 98, 250–258. [Google Scholar] [CrossRef]
- Ye, Y.; Tang, Y.; Xiong, Y.; Feng, L.; Li, X. Bisphenol A exposure alters placentation and causes preeclampsia-like features in pregnant mice involved in reprogramming of DNA methylation of WNT2. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 2732–2742. [Google Scholar] [CrossRef] [Green Version]
- Groff, T. Bisphenol A: Invisible pollution. Curr. Opin. Pediatr. 2010, 22, 524–529. [Google Scholar] [CrossRef]
- Benachour, N.; Aris, A. Toxic effects of low doses of Bisphenol-A on human placental cells. Toxicol. Appl. Pharmacol. 2009, 241, 322–328. [Google Scholar] [CrossRef]
- Narciso, L.; Ietta, F.; Romagnoli, R.; Paulesu, L.; Mantovani, A.; Tait, S. Effects of Bisphenol A on endogenous retroviral envelopes expression and trophoblast fusion in BeWo cells. Reprod. Toxicol. 2019, 89, 35–44. [Google Scholar] [CrossRef]
- Paulesu, L.; Rao, C.V.; Ietta, F.; Pietropolli, A.; Ticconi, C. hCG and Its Disruption by Environmental Contaminants during Human Pregnancy. Int. J. Mol. Sci. 2018, 19, 914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srakovsky, R.S.; Schantz, S.L. Impacts of bisphenol A (BPA) and exposures on epigenetic outcomes in the human placenta. Environ. Epigenet. 2018, 4, dvy022. [Google Scholar]
- Montrose, L.; Padmanabhan, V.; Goodrich, J.M.; Domino, S.E.; Treadwell, M.C.; Meeker, J.D.; Watkins, D.J.; Dolinoy, D.C. Maternal levels of endocrine disrupting chemicals in the first trimester ofpregnancy are associated with infant cord blood DNA methylation. Epigenetics 2018, 13, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, J.E.; Meyer, N.; Santamaria, C.G.; Schumacher, A.; Luque, E.H.; Zenclussen, M.L.; Rodriguez, H.A.; Zenclussen, A.C. Bisphenol A exposure during early pregnancy impairs uterine spiral artery remodeling and provokes intrauterine growth restriction in mice. Sci. Rep. 2018, 8, 9196. [Google Scholar] [CrossRef] [PubMed]
- Timms, B.G.; Howdeshell, K.L.; Barton, L.; Bradley, S. Estrogenic chemicals in plastic and oral contraceptives disrupt development of the fetal mouse prostate and urethra. Biol. Reprod. 2005, 102, 7014–7019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arase, S.; Ishii, K.; Igarashi, K.; Aisaki, K.; Yoshio, Y.; Matsushima, A.; Shimohigashi, Y.; Arima, K.; Kanno, J.; Sugimura, Y. Endocrine disrupter bisphenol A increases in situ estrogen production in the mouse urogenital sinus. Biol. Reprod. 2011, 84, 734–742. [Google Scholar] [CrossRef]
- vom Saal, F.S.; Timms, B.G.; Montano, M.M.; Palanza, P.; Thayer, K.A.; Nagel, S.C.; Dhar, M.D.; Ganjam, V.K.; Parmigiani, S.; Welshons, W.V. Prostate enlargement in mice due to fetal exposure to low doses of estradiol or diethylstilbestrol and opposite effects at high doses. Proc. Natl. Acad. Sci. USA 1997, 94, 2056–2061. [Google Scholar] [CrossRef] [Green Version]
- Shelnutt, S.; Kind, J.; Allaben, W. Bisphenol A: Update on newly developed data and how they address NTP’s 2008finding of “Some Concern”. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 57, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Kolla, S.; Morcos, M.; Martin, B.; Vandenberg, L.N. Low dose bisphenol S or ethinyl estradiol exposures during the perinatal period alter female mouse mammary gland development. Reprod. Toxicol. 2018, 78, 50–59. [Google Scholar] [CrossRef]
- Menone, M.L.; Aizpún de Moreno, J.E.; Moreno, V.J.; Lanfranchi, A.L.; Metcalfe, T.L.; Metcalfe, C.D. Organochlorine Pesticides and PCBs in a Southern Atlantic Coastal Lagoon Watershed, Argentina. Arch. Environ. Con. Tox. 2001, 40, 355–362. [Google Scholar]
- Fair, P.A.; White, N.D.; Wolf, B.; Arnott, S.A.; Kannan, K.; Karthikraj, R.; Vena, J.E. Persistent organic pollutants in fish from Charleston Harbor and tributaries, South Carolina, United States: A risk assessment. Environ. Res. 2018, 167, 598–613. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, R.; Megha, P.; Sreedev, P. Organochlorine pesticides, their toxic effects on living organisms and their fatein the environment. Interdiscip. Toxicol. 2016, 9, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandiera, S.M.; Torok, S.M.; Letcher, R.J.; Norstrom, R.J. Immunoquantitation of cytochromes P450 1A and P450 2B and comparison with chlorinated hydrocarbon levels in archived polar bear liver samples. Chemosphere 1997, 34, 1469–1479. [Google Scholar] [CrossRef]
- Docea, A.O.; Vassilopoulou, L.; Fragou, D.; Arsene, A.L.; Fenga, C.; Kovatsi, L.; Petrakis, D.; Rakitskii, V.N.; Nosyrev, A.E.; Izotov, B.N.; et al. CYP polymorphisms and pathological conditions related to chronic exposure to organochlorine pesticides. Toxicol. Rep. 2017, 4, 335–341. [Google Scholar] [CrossRef]
- Li, Y.; Millikan, R.C.; Bell, D.A.; Cui, L.; Tse, C.J.; Newman, B.; Conway, K. Polychlorinated biphenyls, cytochrome P450 1A1 (CYP1A1) polymorphisms, and breast cancer risk among African American women and white women in North Carolina: A population-based case-control study. Breast Cancer Res. BCR 2005, 7, R12–R18. [Google Scholar] [CrossRef] [Green Version]
- Mrema, E.J.; Rubino, F.M.; Brambilla, G.; Moretto, A.; Tsatsakis, A.M.; Colosio, C. Persistent organochlorinated pesticides and mechanisms of their toxicity. Toxicology 2013, 307, 74–88. [Google Scholar] [CrossRef]
- Lopez-Espinosa, M.J.; Granada, A.; Carreno, J. Organochlorine Pesticides in Placentas from Southern Spain and Some Related Factors. Placenta 2007, 28, 631–638. [Google Scholar] [CrossRef]
- Kumar, S. Occupational exposure associated with reproductive dysfunction. J. Occup. Health 2004, 46, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Anand, M.; Agarwal, P.; Singh, L. Persistent organochlorine pesticides and oxidant/antioxidant status in the placental tissue of the women with full-term and pre-term deliveries. Toxicol. Res. 2015, 4, 326–332. [Google Scholar] [CrossRef]
- Anand, M.; Singh, L.; Agarwal, P.; Saroj, R.; Taneja, A. Pesticides exposure through environment and risk of pre-term birth: A study from Agra city. Drug Chem. Toxicol. 2019, 42, 471–477. [Google Scholar] [CrossRef]
- Pathak, R.; Mustafa, M.; Ahmed, R.S.; Tripathi, A.K.; Guleria, K.; Banerjee, B.D. Association between recurrent miscarriages and organochlorine pesticide levels. Clin. Biochem. 2010, 43, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.H.B.; Polder, A.; Brynildsrud, O.B.; Gronnestad, R.; Karimi, M.; Lie, E.; Manyilizu, W.B.; Mdegela, R.H.; Mokiti, F.; Murtadha, M.; et al. Prenatal exposure to persistent organic pollutants in Northern Tanzania and their distribution between breast milk, maternal blood, placenta and cord blood. Environ. Res. 2019, 170, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Cho, Y.H.; Lee, I.; Kim, W.; Won, S.; Ku, J.; Moon, H.; Park, J.; Kim, S.; Choi, G.; et al. Prenatal exposure to persistent organic pollutants and methylation of LINE-1 and imprinted genes in placenta: A CHECK cohort study. Environ. Int. 2018, 119, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Dewan, P.; Jain, V.; Gupta, P.; Banerjeeb, B.D. Organochlorine pesticide residues in maternal blood, cord blood, placenta, and breastmilk and their relation to birth size. Chemosphere 2013, 90, 1704–1710. [Google Scholar] [CrossRef]
- Freire, C.; Lopez-Espinosa, M.; Fernández, M. Prenatal exposure to organochlorine pesticides and TSH status in newborns from Southern Spain. Sci. Total Environ. 2011, 409, 3281–3287. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Cheng, Y.; Tang, Q.; Lin, S.; Li, Y.; Hu, X.; Nian, J.; Gu, H.; Lu, Y.; Tang, H.; et al. The association between prenatal exposure to organochlorine pesticides and thyroid hormone levels in newborns in Yancheng, China. Environ. Res. 2014, 129, 47–51. [Google Scholar] [CrossRef]
- Freire, C.; Koifman, R.J.; Sarcinelli, P.N.; Simoes Rosa, A.C.; Clapauch, R.; Koifman, S. Long-term exposure to organochlorine pesticides and thyroid status in adults in a heavily contaminated area in Brazil. Environ. Res. 2013, 127, 7–15. [Google Scholar] [CrossRef]
- Bonde, J.P.; Flachs, E.M.; Rimborg, S.; Glazer, C.H.; Giwercman, A.; Ramlau-Hansen, C.H.; Hougaard, K.S.; Hoyer, B.B.; Haervig, K.K.; Petersen, S.B.; et al. The epidemiologic evidence linking prenatal and postnatal exposure to endocrine disrupting chemicals with male reproductive disorders: A systematic review and meta-analysis. Hum. Reprod. Update 2016, 23, 104–125. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Yadav, C.S.; Singh, S.; Goel, S.; Ahmed, R.S.; Gupta, S.; Grover, R.K.; Banerjee, B.D. CYP 1A1 polymorphism and organochlorine pesticides levels in the etiology of prostate cancer. Chemosphere 2010, 81, 464–468. [Google Scholar] [CrossRef]
- Korach, K.S.; Metzler, M.; McLachlan, J.A. Estrogenic activity in vivo and in vitro of some diethylstilbestrol metabolitesand analogs. Proc. Natl. Acad. Sci. USA 1978, 75, 468–471. [Google Scholar] [CrossRef] [Green Version]
- Herbst, A.L.; Ulfelder, H.; Poskanzer, D.C.; Longo, L.D. Adenocarcinoma of the vagina. Association of maternal stilbestrol therapy with tumor appearance in young women. Am. J. Obstet. Gynecol. 1999, 181, 1574–1575. [Google Scholar] [CrossRef]
- Veurink, M.; Koster, M.; Berg, L.T.W.D. The history of DES, lessons to be learned. Pharm. World Sci. 2005, 27, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Colton, T.; Greenberg, E.R.; Noller, K.; Resseguie, L.; Van Bennekom, C.; Heeren, T.; Zhang, Y. Breast cancer in mothers prescribed diethylstilbestrol in pregnancy. Further follow-up. JAMA 1993, 269, 2096–2100. [Google Scholar] [CrossRef] [PubMed]
- Bibbo, M.; Haenszel, W.M.; Wied, G.L.; Hubby, M.; Herbst, A.L. A twenty-five-year follow-up study of women exposed to diethylstilbestrol during pregnancy. N. Engl. J. Med. 1978, 298, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, E.R.; Barnes, A.B.; Resseguie, L.; Barrett, J.A.; Burnside, S.; Lanza, L.L.; Neff, R.K.; Stevens, M.; Young, R.H.; Colton, T. Breast cancer in mothers given diethylstilbestrol in pregnancy. N. Engl. J. Med. 1984, 311, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Titus-Ernstoff, L.; Hatch, E.E.; Hoover, R.N.; Palmer, J.; Greenberg, E.R.; Ricker, W.; Kaufman, R.; Noller, K.; Herbst, A.L.; Colton, T.; et al. Long-term cancer risk in women given diethylstilbestrol (DES) during pregnancy. Br. J. Cancer 2001, 84, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Wang, R.; Li, M.; Sun, T.; Zhou, Y.; Li, Y.; Sun, L.; Zhang, B.; Lian, Z.; Xue, S.; et al. Melatonin Reduces Androgen Production and Upregulates Heme Oxygenase-1 Expression in Granulosa Cells from PCOS Patients with Hypoestrogenia and Hyperandrogenia. Oxid. Med. Cell. Longev. 2019, 2019, 13. [Google Scholar] [CrossRef]
- Newbold, R.R.; Hanson, R.B.; Jefferson, W.N.; Bullock, B.C.; Haseman, J.; McLachlan, J.A. Increased tumors but uncompromised fertility in the female descendants of mice exposed developmentally to diethylstilbestrol. Carcinogenesis 1998, 19, 1655–1663. [Google Scholar] [CrossRef] [Green Version]
- Newbold, R.R.; Padilla-Banks, E.; Jefferson, W.N. Adverse effects of the model environmental estrogen diethylstilbestrol are transmitted to subsequent generations. Endocrinology 2006, 147, S11–S17. [Google Scholar] [CrossRef]
- Wilcox, A.J.; Baird, D.D.; Weinberg, C.R.; Hornsby, P.P.; Herbst, A.L. Fertility in men exposed prenatally to diethylstilbestrol. N. Engl. J. Med. 1995, 332, 1411–1416. [Google Scholar] [CrossRef]
- Al Jishi, T.; Sergi, C. Current perspective of diethylstilbestrol (DES) exposure in mothers and offspring. Reprod. Toxicol. 2017, 71, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Newbold, R.R. Prenatal exposure to diethylstilbestrol (DES). Fertil. Steril. 2008, 89, e55–e56. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S. Expression of estrogen-regulated genes during development in the mouse uterus exposed to diethylstilbestrol neonatally. Curr. Pharm. Des. 2006, 12, 1505–1520. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Hussain, I.; Ansari, K.I.; Bobzean, S.A.M.; Perrotti, L.I.; Mandal, S.S. Histone methyltransferase EZH2 is transcriptionally induced by estradiol as well as estrogenic endocrine disruptors bisphenol-A and diethylstilbestrol. J. Mol. Biol. 2014, 426, 3426–3441. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; Abbas, I.K.; Menard, M.; Singh, U.P.; Zhang, J.; Nagarkatti, P.; Nagarkatti, M. Exposure to diethylstilbestrol during pregnancy modulates microRNA expression profile in mothers and fetuses reflecting oncogenic and immunological changes. Mol. Pharmacol. 2015, 87, 842–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikula, P.; Svobodova, Z.; Smutna, M. Phthalates: Toxicology and Food Safety—a Review. Czech J. Food Sci. 2005, 23, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Ito, R.; Seshimo, F.; Miura, N.; Kawaguchi, M.; Saito, K.; Nakazawa, H. Effect of sterilization process on the formation of mono(2-ethylhexyl) phthalate from di(2-ethylhexyl) phthalate. J. Pharm. Biomed. Anal. 2006, 41, 455–460. [Google Scholar] [CrossRef]
- Zhao, Y.; Ao, H.; Chen, L.; Sottas, C.M.; Ge, R.; Li, L.; Zhang, Y. Mono-(2-ethylhexyl) phthalate affects the steroidogenesis in rat Leydig cells through provoking ROS perturbation. Toxicol. Int. J. Publ. Assoc. BIBRA 2012, 26, 950–955. [Google Scholar] [CrossRef]
- Shoaito, H.; Petit, J.; Chissey, A.; Auzeil, N.; Guibourdenche, J.; Gil, S.; Laprevote, O.; Fournier, T.; Degrelle, S.A. The Role of Peroxisome Proliferator-Activated Receptor Gamma (PPAR gamma) in Mono (2-ethylhexyl) Phthalate (MEHP)-Mediated Cytotrophoblast Differentiation. Environ. Health Perspect. 2019, 127, 27003. [Google Scholar] [CrossRef]
- Valinluck, V.; Tsai, H.; Rogstad, D.K.; Burdzy, A.; Bird, A.; Sowers, L.C. Oxidative damage to methyl-CpG sequences inhibits the binding of the methyl-CpG binding domain (MBD) of methyl-CpG binding protein 2 (MeCP2). Nucleic Acids Res. 2004, 32, 4100–4108. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Hu, W.; Li, Y.; Shen, H.; Hu, J. Mono-2-ethylhexyl phthalate inhibits human extravillous trophoblast invasion via the PPARgamma pathway. Toxicol. Appl. Pharm. 2017, 327, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, K.E.; Morrish, D.; Roth, I.; Bhardwaj, D.; Taylor, R.; Zhou, Y.; Fisher, S.J. Human cytotrophoblast invasion is up-regulated by epidermal growth factor: Evidence that paracrine factors modify this process. Dev. Biol. 1994, 164, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Grindler, N.M.; Vanderlinden, L.; Karthikraj, R.; Kannan, K.; Teal, S.; Polotsky, A.J.; Powell, T.L.; Yang, I.V.; Jansson, T. Exposure to Phthalate, an Endocrine Disrupting Chemical, Alters the First Trimester Placental Methylome and Transcriptome in Women. Sci. Rep. 2018, 8, 6086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, L.; Cao, Y.; Chen, B.; Zheng, L.; Ge, R. Phthalate levels and low birth weight: A nested case-control study of Chinesenewborns. J. Pediatr. 2009, 155, 500–504. [Google Scholar] [CrossRef]
- Buck Louis, G.M.; Sundaram, R.; Sweeney, A.M.; Schisterman, E.F.; Maisog, J.; Kannan, K. Urinary bisphenol A, phthalates, and couple fecundity: The Longitudinal Investigation of Fertility and the Environment (LIFE) Study. Fertil. Steril. 2014, 101, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Toft, G.; Jonsson, B.A.G.; Lindh, C.H.; Jensen, T.K.; Hjollund, N.H.; Vested, A.; Bonde, J.P. Association between pregnancy loss and urinary phthalate levels around the time of conception. Environ. Health Perspect. 2012, 120, 458–463. [Google Scholar] [CrossRef] [Green Version]
- Jukic, A.M.; Calafat, A.M.; McConnaughey, D.R.; Longnecker, M.P.; Hoppin, J.A.; Weinberg, C.R.; Wilcox, A.J.; Baird, D.D. Urinary Concentrations of Phthalate Metabolites and Bisphenol A and Associations with Follicular-Phase Length, Luteal-Phase Length, Fecundability, and Early Pregnancy Loss. Environ. Health Perspect. 2016, 124, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Petit, J.; Wakx, A.; Gil, S.; Fournier, T.; Auzeil, N.; Rat, P.; Laprevote, O. Lipidome-wide disturbances of human placental JEG-3cells by the presence of MEHP. Biochimie 2018, 149, 1–8. [Google Scholar] [CrossRef]
- Philippat, C.; Heude, B.; Botton, J.; Alfaidy, N.; Calafat, A.M.; Slama, R. Prenatal Exposure to Select Phthalates and Phenols and Associations with Fetal and Placental Weight among Male Births in the EDEN Cohort (France). Environ. Health Perspect. 2019, 127, 17002. [Google Scholar] [CrossRef] [Green Version]
- Kilcoyne, K.R.; Mitchell, R.T. Effect of environmental and pharmaceutical exposures on fetal testis development and function: A systematic review of human experimental data. Hum. Reprod. Update 2019, 25, 397–421. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.S.; Anand-Ivell, R.; Norgaard-Pedersen, B.; Jonsson, B.A.G.; Bonde, J.P.; Hougaard, D.M.; Cohen, A.; Lindh, C.H.; Ivell, R.; Toft, G. Amniotic fluid phthalate levels and male fetal gonad function. Epidemiology (Camb. Mass.) 2015, 26, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Kuhnle, G.G.C.; Dell’Aquila, C.; Aspinall, S.M.; Runswick, S.A.; Mulligan, A.A.; Bingham, S.A. Phytoestrogen content of beverages, nuts, seeds, and oils. J. Agric. Food Chem. 2008, 56, 7311–7315. [Google Scholar] [CrossRef] [PubMed]
- Tucak, M.; Horvat, D.; Cupic, T.; Krizmanic, G.; Tomas, V.; Ravlic, M.; Popovic, S. Forage Legumes as Sources of Bioactive Phytoestrogens for Use in Pharmaceutics: A Review. Curr. Pharm. Biotechnol. 2018, 19, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Bukovsky, A.; Caudle, M.R.; Cekanova, M.; Fernando, R.I.; Wimalasena, J.; Foster, J.S.; Henley, D.C.; Elder, R.F. Placental expression of estrogen receptor beta and its hormone binding variant—comparison with estrogen receptor alpha and a role for estrogen receptors in asymmetric division and differentiation of estrogen-dependent cells. Reprod. Biol. Endocrinol. 2003, 1, 36. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Zhang, R.; Lian, Z.; Deng, S.; Yu, K. Estrogen-Receptor Expression and Function in Female Reproductive Disease. Cells 2019, 8, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, M.; Dahlman-Wright, K.; Gustafsson, J. Estrogen receptor alpha and beta in health and disease. Best Pract. Research. Clin. Endocrinol. Metab. 2015, 29, 557–568. [Google Scholar] [CrossRef]
- Awobajo, F.; Abdul, M.; Aminu, B.; Medobi, E. Alteration to PGF and IGF-1, signalled the adverse growth of the foetus and placenta in a genistein exposed pregnancy in experimental rats. Endocr. Abstr. 2018, 59, 192. [Google Scholar] [CrossRef]
- Jeschke, U.; Briese, V.; Richter, D.; Bruer, G.; Plessow, D.; Waldschlager, J.; Mylonas, I.; Friese, K. Effects of phytoestrogens genistein and daidzein on production of human chorionicgonadotropin in term trophoblast cells in vitro. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2005, 21, 180–184. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| EDCs Exposure | Compound | Area | Measurement Method | Body Fluid | Mean ± SD or Range | Reference |
|---|---|---|---|---|---|---|
| Bisphenol A | BPA | Germany | Chemical derivatization-GC-MS | Maternal plasma | 0.3–18.9 ng/mL | [31] |
| BPA | Germany | GC-MS | Fetal plasma | 0.2–9.2 ng/mL | [31] | |
| BPA | Japan | Enzyme-linked immunosorbent assay | Amniotic fluids | 0–8.38 ng/mL | [32,33] | |
| BPA | Germany | GC-MS | The placental tissue | 1–104.4 ng/g | [31,34] | |
| Organochlorine pesticides | α-hexachlorocyclohexane (α-HCH) | The Los Angeles area | GC-MS | Amniotic fluids | 0.15 ± 0.06 ng/mL | [35] |
| p,p′-Dichlorodiphenyldichloroethylene (p,p’-DDE) | The Los Angeles area | GC-MS | Amniotic fluids | 0.21 ± 0.18 ng/mL | [35] | |
| p,p’-Dichloro-diphenyl-trichloroethane (p,p′-DDT) | Mexico | Dual column gas chromatograph-electron capture detector | Serum | 676 ng/g-lipid | [36] | |
| p,p′-DDE | Mexico | Dual column gas chromatograph-electron capture detector | Serum | 4,843 ng/g-lipid | [36] | |
| Phthalates | Methylerythritol cyclodiphosphate (MECPP) | America | Isotope-dilution HPLC/MS/MS | Breast milk | 0.1–0.4 μg/L | [37] |
| MEHHP | America | HPLC/MS/MS | Breast milk | 0.2–0.3 μg/L | [37] | |
| Mono-ethyl phthalate (MEP) | Canada | LC-MS/MS | Urine | 32.02 μg/L | [38] | |
| Mono-n-butyl phthalate (MnBP) | Canada | LC-MS / MS | Urine | 11.59 μg/L | [38] | |
| Diethylstilbestrol | DES | China | HPLC with diode array detection | Urine | 1–200 μg/L | [39] |
| Phytoestrogens | Daidzein | America | GC/MS | Amniotic fluid | 1.44–5.52 ng/mL | [40] |
| Genistein | America | GC/MS | Amniotic fluid | 1.69–6.54 ng/mL | [40] | |
| Daidzein | America | Ultra-performance liquid chromatography-MS/MS (UPLC-MS/MS) | Urine | 0.11 μg/L | [41] | |
| Genistein | America | UPLC-MS/MS | Urine | 0.59 μg/L | [41] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Z.-R.; Xu, X.-L.; Deng, S.-L.; Lian, Z.-X.; Yu, K. Oestrogenic Endocrine Disruptors in the Placenta and the Fetus. Int. J. Mol. Sci. 2020, 21, 1519. https://doi.org/10.3390/ijms21041519
Tang Z-R, Xu X-L, Deng S-L, Lian Z-X, Yu K. Oestrogenic Endocrine Disruptors in the Placenta and the Fetus. International Journal of Molecular Sciences. 2020; 21(4):1519. https://doi.org/10.3390/ijms21041519
Chicago/Turabian StyleTang, Zi-Run, Xue-Ling Xu, Shou-Long Deng, Zheng-Xing Lian, and Kun Yu. 2020. "Oestrogenic Endocrine Disruptors in the Placenta and the Fetus" International Journal of Molecular Sciences 21, no. 4: 1519. https://doi.org/10.3390/ijms21041519
APA StyleTang, Z. -R., Xu, X. -L., Deng, S. -L., Lian, Z. -X., & Yu, K. (2020). Oestrogenic Endocrine Disruptors in the Placenta and the Fetus. International Journal of Molecular Sciences, 21(4), 1519. https://doi.org/10.3390/ijms21041519