Detection of LINE-1 hypomethylation in cfDNA of Esophageal Adenocarcinoma Patients

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
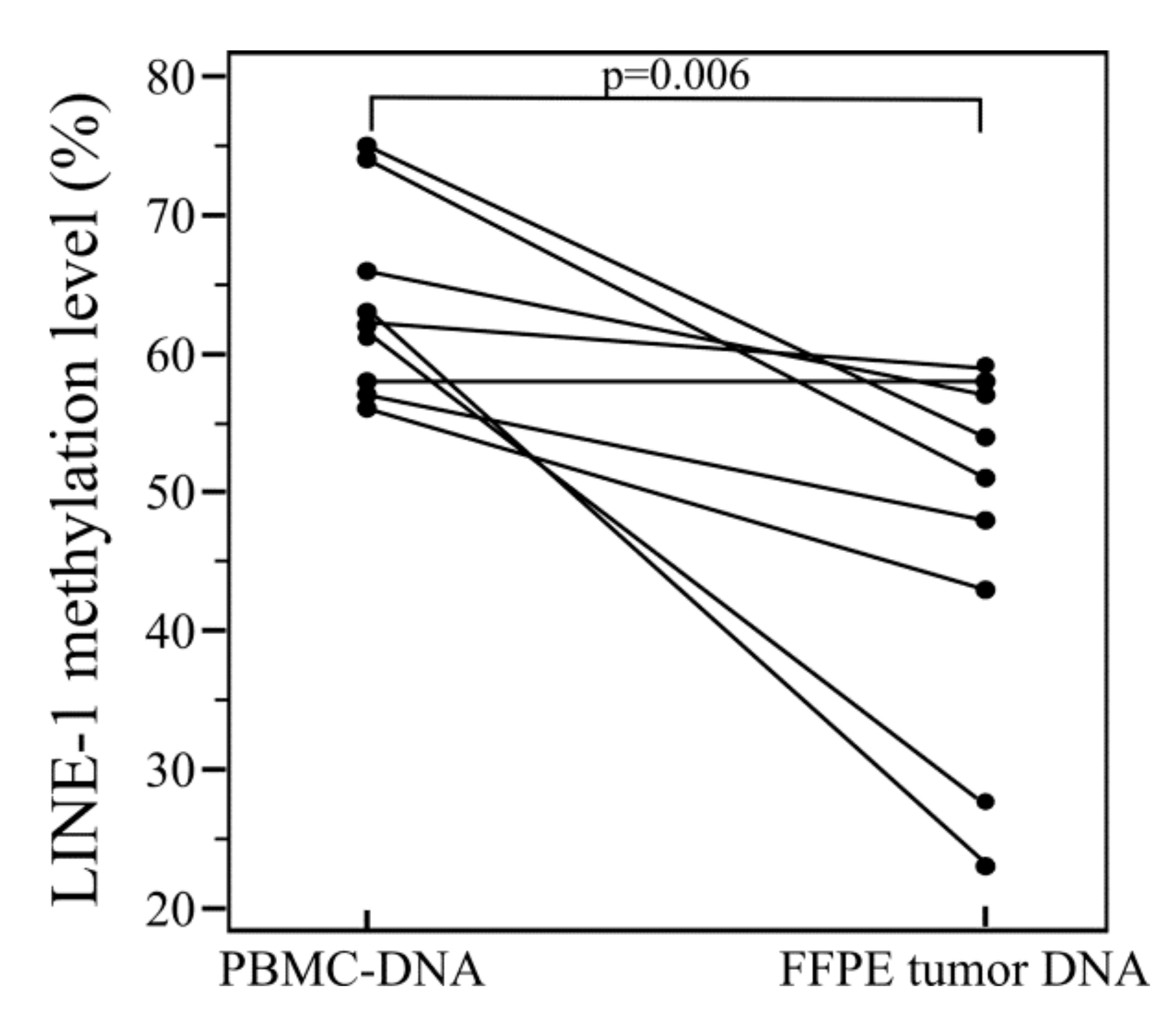
2.1. LINE-1 Methylation Analysis in Formalin Fixed Paraffin Embedded (FFPE)-DNA
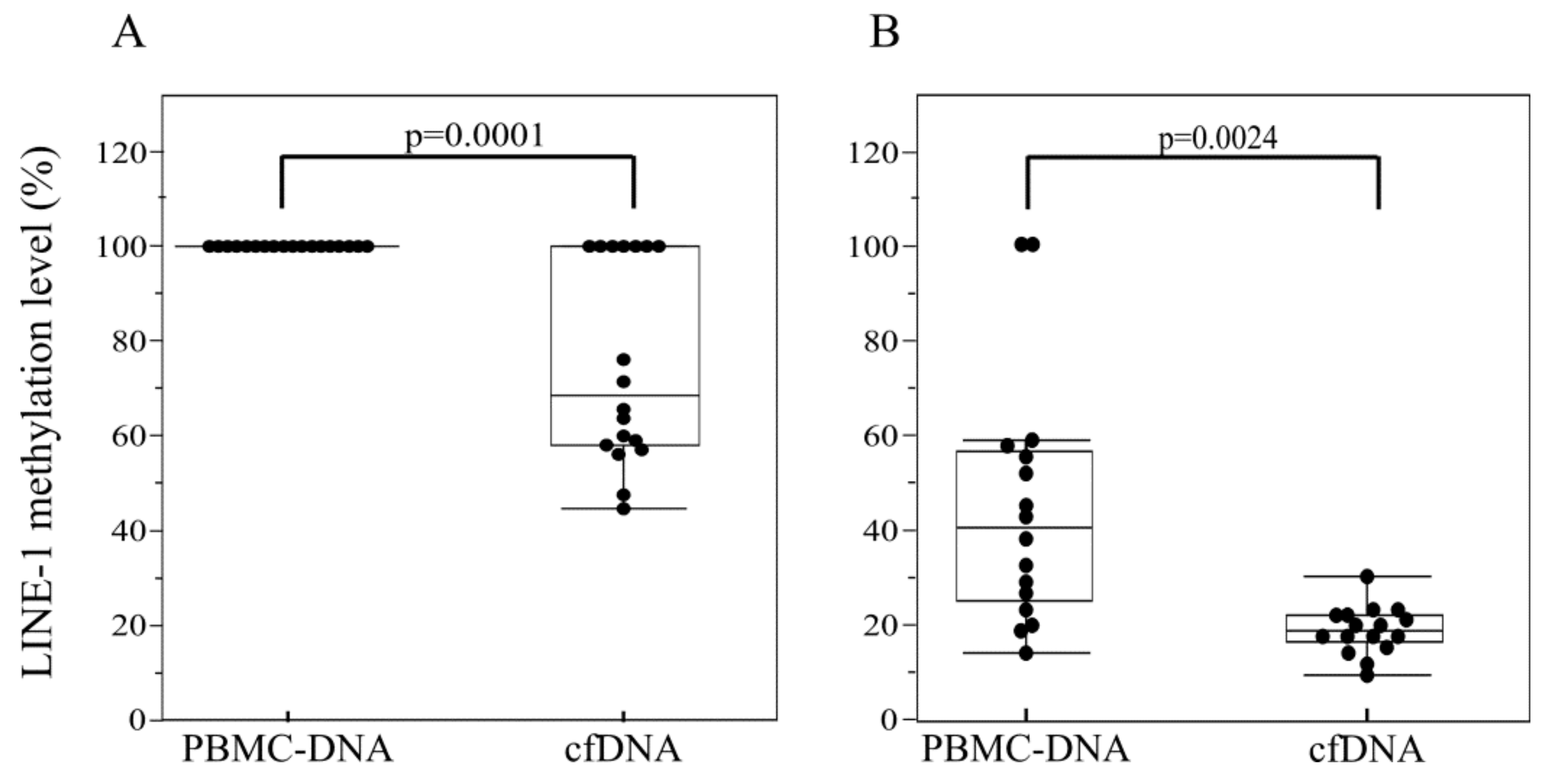
2.2. LINE-1 Methylation Analysis in cfDNA
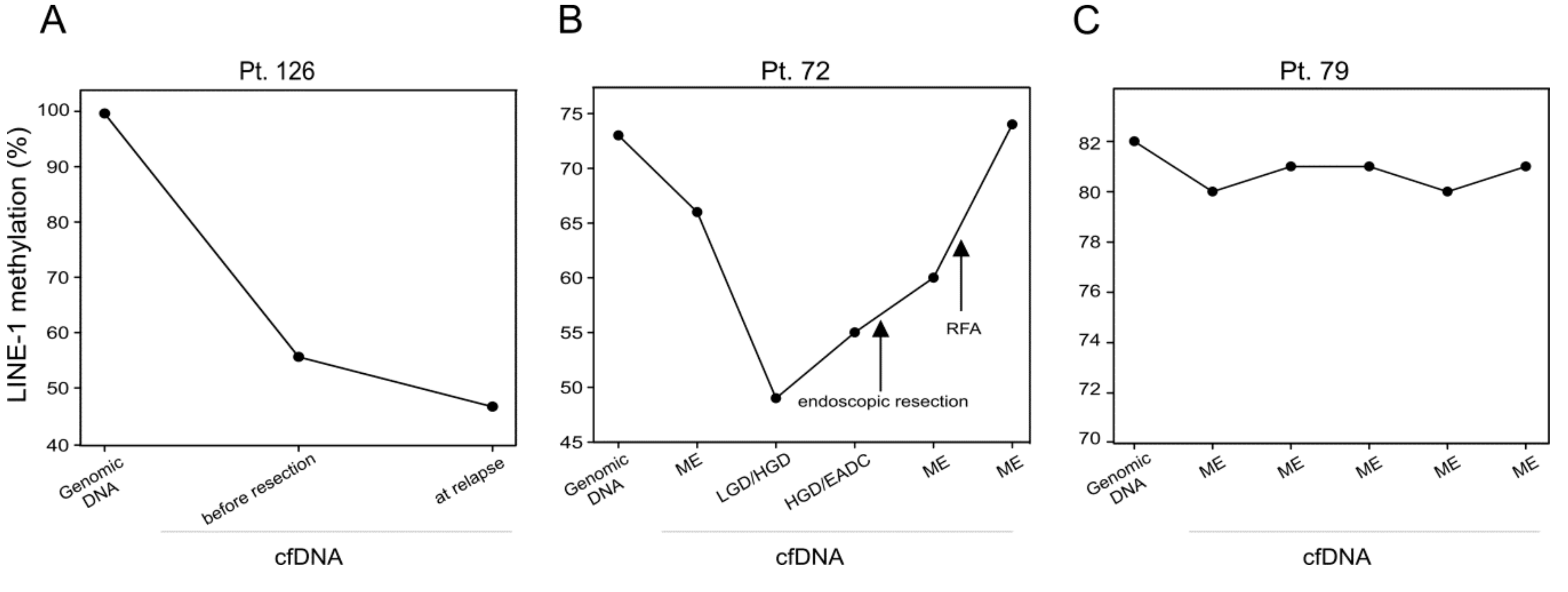
2.3. LINE-1 Methylation Analysis in cfDNA of Longitudinal Cases
3. Materials and Methods
3.1. Patients
3.2. DNA Extraction
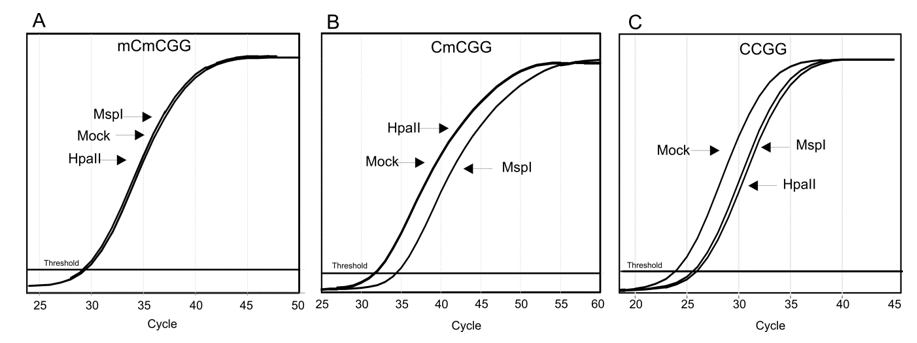
3.3. Methylation-Sensitive Restriction Enzyme Digestion and rtPCR
3.4. Statistics
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Deng, J.; Wang, C.; Xiang, M.; Liu, F.; Liu, Y.; Zhao, K. Meta-analysis of postoperative efficacy in patients receiving chemoradiotherapy followed by surgery for resectable esophageal carcinoma. Diagn. Pathol. 2014, 9, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronellenfitsch, U.; Schwarzbach, M.; Hofheinz, R.; Kienle, P.; Kieser, M.; Slanger, T.E.; Jensen, K.; Group, G.A.M. Perioperative chemo (radio) therapy versus primary surgery for resectable adenocarcinoma of the stomach, gastroesophageal junction, and lower esophagus. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hagen, P.; Hulshof, M.C.; van Lanschot, J.J.; Steyerberg, E.W.; van Berge Henegouwen, M.I.; Wijnhoven, B.P.; Richel, D.J.; Nieuwenhuijzen, G.A.; Hospers, G.A.; Bonenkamp, J.J.; et al. Preoperative chemoradiotherapy for esophageal or junctional cancer. N. Engl. J. Med. 2012, 366, 2074–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubecz, A.; Solymosi, N.; Stadlhuber, R.J.; Schweigert, M.; Stein, H.J.; Peters, J.H. Does the Incidence of Adenocarcinoma of the Esophagus and Gastric Cardia Continue to Rise in the Twenty-First Century?-a SEER Database Analysis. J. Gastrointest. Surg. 2013, 18, 124–129. [Google Scholar] [CrossRef]
- Edgren, G.; Adami, H.O.; Weiderpass, E.; Weiderpass Vainio, E.; Nyrén, O. A global assessment of the oesophageal adenocarcinoma epidemic. Gut 2013, 62, 1406–1414. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network; Analysis Working Group: Asan University; BC Cancer Agency; Brigham and Women’s Hospital; Broad Institute; Brown University; Dana-Farber Cancer Institute; Duke University; Harvard Medical School; Institute for Systems Biology; et al. Integrated genomic characterization of oesophageal carcinoma. Nature 2017, 541, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Polineni, R.; Hussein, Z.; Vigoda, I.; Bhagat, T.D.; Bhattacharyya, S.; Maitra, A.; Verma, A. Role of epigenetic alterations in the pathogenesis of Barrett’s esophagus and esophageal adenocarcinoma. Int. J. Clin. Exp. Pathol. 2012, 5, 382–396. [Google Scholar]
- Moinova, H.R.; LaFramboise, T.; Lutterbaugh, J.D.; Chandar, A.K.; Dumot, J.; Faulx, A.; Brock, W.; De la Cruz Cabrera, O.; Guda, K.; Barnholtz-Sloan, J.S.; et al. Identifying DNA methylation biomarkers for non-endoscopic detection of Barrett’s esophagus. Sci. Transl. Med. 2018, 10, eaao5848. [Google Scholar] [CrossRef] [Green Version]
- Kaz, A.M.; Grady, W.M. Epigenetic biomarkers in esophageal cancer. Cancer Lett. 2014, 342, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, M. DNA hypomethylation in cancer cells. Epigenomics 2009, 1, 239–259. [Google Scholar] [CrossRef] [Green Version]
- Van Tongelen, A.; Loriot, A.; De Smet, C. Oncogenic roles of DNA hypomethylation through the activation of cancer-germline genes. Cancer Lett. 2017, 396, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Xiao-Jie, L.; Hui-Ying, X.; Qi, X.; Jiang, X.; Shi-Jie, M. LINE-1 in cancer: Multifaceted functions and potential clinical implications. Genet. Med. 2016, 18, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordaux, R.; Batzer, M.A. The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 2009, 10, 691–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitkumthorn, N.; Mutirangura, A. Long interspersed nuclear element-1 hypomethylation in cancer: Biology and clinical applications. Clin. Epigenet. 2011, 2, 315–330. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Jamil, M.A.; Nuesgen, N.; Dauksa, A.; Gulbinas, A.; Schulz, W.A.; Oldenburg, J.; El-Maarri, O. Detailed methylation map of LINE-1 5’-promoter region reveals hypomethylated CpG hotspots associated with tumor tissue specificity. Mol. Genet. Genom. Med. 2019, 7, e601. [Google Scholar] [CrossRef] [Green Version]
- Levine, D.M.; Ek, W.E.; Zhang, R.; Liu, X.; Onstad, L.; Sather, C.; Lao-Sirieix, P.; Gammon, M.D.; Corley, D.A.; Shaheen, N.J.; et al. A genome-wide association study identifies new susceptibility loci for esophageal adenocarcinoma and Barrett’s esophagus. Nat. Genet. 2013, 45, 1487–1493. [Google Scholar] [CrossRef] [Green Version]
- Bronkhorst, A.J.; Aucamp, J.; Pretorius, P.J. Cell-free DNA: Preanalytical variables. Clin. Chim. Acta 2015, 450, 243–253. [Google Scholar] [CrossRef]
- Boldrin, E.; Rumiato, E.; Fassan, M.; Balsamo, L.; Realdon, S.; Battaglia, G.; Rugge, M.; Amadori, A.; Saggioro, D. Liquid biopsy as a novel tool to monitor the carcinogenesis of Barrett’s esophagus. Transl. Res. 2016, 176, 127–131. [Google Scholar] [CrossRef]
- Rumiato, E.; Boldrin, E.; Malacrida, S.; Realdon, S.; Fassan, M.; Morbin, T.; Battaglia, G.; Amadori, A.; Rugge, M.; Saggioro, D. Detection of genetic alterations in cfDNA as a possible strategy to monitor the neoplastic progression of Barrett’s esophagus. Transl. Res. 2017, 190, 16–24. [Google Scholar] [CrossRef]
- Eads, C.A.; Lord, R.V.; Wickramasinghe, K.; Long, T.I.; Kurumboor, S.K.; Bernstein, L.; Peters, J.H.; DeMeester, S.R.; DeMeester, T.R.; Skinner, K.A.; et al. Epigenetic patterns in the progression of esophageal adenocarcinoma. Cancer Res. 2001, 61, 3410–3418. [Google Scholar]
- Kawakami, K.; Brabender, J.; Lord, R.V.; Groshen, S.; Greenwald, B.D.; Krasna, M.J.; Yin, J.; Fleisher, A.S.; Abraham, J.M.; Beer, D.G.; et al. Hypermethylated APC DNA in plasma and prognosis of patients with esophageal adenocarcinoma. J. Natl. Cancer Inst. 2000, 92, 1805–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, M.V.; Gou, M.; Akiyama, Y.; Muller, A.; Wu, T.T.; Montgomery, E.; Deasel, M.; Germonpré, P.; Rubinson, L.; Heitmiller, R.F.; et al. Prognostic importance of promoter hypermethylation of multiple genes in esophageal adenocarcinoma. Clin. Cancer Res. 2003, 9, 2912–2919. [Google Scholar] [PubMed]
- Hamilton, J.P.; Sato, F.; Jin, Z.; Greenwald, B.D.; Ito, T.; Mori, Y.; Paun, B.C.; Kan, T.; Cheng, Y.; Wang, S.; et al. Reprimo methylation is a potential biomarker of Barrett’s-Associated esophageal neoplastic progression. Clin. Cancer Res. 2006, 12, 6637–6642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Z.; Mori, Y.; Yang, J.; Sato, F.; Ito, T.; Cheng, Y.; Paun, B.; Hamilton, J.P.; Kan, T.; Olaru, A.; et al. Hypermethylation of the nel-like 1 gene is a common and early event and is associated with poor prognosis in early-stage esophageal adenocarcinoma. Oncogene 2007, 26, 6332–6340. [Google Scholar] [CrossRef] [Green Version]
- Toh, Y.; Egashira, A.; Yamamoto, M. Epigenetic alterations and their clinical implications in esophageal squamous cell carcinoma. Gen. Thorac. Cardiovasc. Surg. 2013, 61, 262–269. [Google Scholar] [CrossRef]
- Iwagami, S.; Baba, Y.; Watanabe, M.; Shigaki, H.; Miyake, K.; Ida, S.; Nagai, Y.; Ishimoto, T.; Iwatsuki, M.; Sakamoto, Y.; et al. Pyrosequencing assay to measure LINE-1 methylation level in esophageal squamous cell carcinoma. Ann. Surg. Oncol. 2012, 19, 2726–2732. [Google Scholar] [CrossRef]
- Zhu, J.; Ling, Y.; Xu, Y.; Lu, M.Z.; Liu, Y.P.; Zhang, C.S. Elevated expression of MDR1 associated with Line-1 hypomethylation in esophageal squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 14392–14400. [Google Scholar]
- Baba, Y.; Watanabe, M.; Murata, A.; Shigaki, H.; Miyake, K.; Ishimoto, T.; Iwatsuki, M.; Iwagami, S.; Yoshida, N.; Oki, E.; et al. LINE-1 hypomethylation, DNA copy number alterations, and CDK6 amplification in esophageal squamous cell carcinoma. Clin. Cancer Res. 2014, 20, 1114–1124. [Google Scholar] [CrossRef] [Green Version]
- Iwagami, S.; Baba, Y.; Watanabe, M.; Shigaki, H.; Miyake, K.; Ishimoto, T.; Iwatsuki, M.; Sakamaki, K.; Ohashi, Y.; Baba, H. LINE-1 hypomethylation is associated with a poor prognosis among patients with curatively resected esophageal squamous cell carcinoma. Ann. Surg. 2013, 257, 449–455. [Google Scholar] [CrossRef]
- Doucet-O’Hare, T.T.; Rodić, N.; Sharma, R.; Darbari, I.; Abril, G.; Choi, J.A.; Young Ahn, J.; Cheng, Y.; Anders, R.A.; Burns, K.H.; et al. LINE-1 expression and retrotransposition in Barrett’s esophagus and esophageal carcinoma. Proc. Natl. Acad. Sci. USA 2015, 112, E4894–E4900. [Google Scholar] [CrossRef] [Green Version]
- Oakes, C.C.; La Salle, S.; Robaire, B.; Trasler, J.M. Evaluation of a quantitative DNA methylation analysis technique using methylation-sensitive/dependent restriction enzymes and real-time PCR. Epigenetics 2006, 1, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Shin, T.J.; Kim, W.H.; Cho, J.Y. Methylation of LINE-1 in cell-free DNA serves as a liquid biopsy biomarker for human breast cancers and dog mammary tumors. Sci. Rep. 2019, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Fulneček, J.; Kovařík, A. How to interpret methylation sensitive amplified polymorphism (MSAP) profiles? BMC Genet. 2014, 15, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerachian, M.A.; Kerachian, M. Long interspersed nucleotide element-1 (LINE-1) methylation in colorectal cancer. Clin. Chim. Acta 2019, 488, 209–214. [Google Scholar] [CrossRef]
- Furlan, C.; Polesel, J.; Barzan, L.; Franchin, G.; Sulfaro, S.; Romeo, S.; Colizzi, F.; Rizzo, A.; Baggio, V.; Giacomarra, V.; et al. Prognostic significance of LINE-1 hypomethylation in oropharyngeal squamous cell carcinoma. Clin. Epigenet. 2017, 9, 58. [Google Scholar] [CrossRef]
- Imperatori, A.; Sahnane, N.; Rotolo, N.; Franzi, F.; Nardecchia, E.; Libera, L.; Romualdi, C.; Cattoni, M.; Sessa, F.; Dominioni, L.; et al. LINE-1 hypomethylation is associated to specific clinico-pathological features in Stage I non-small cell lung cancer. Lung Cancer 2017, 108, 83–89. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boldrin, E.; Curtarello, M.; Dallan, M.; Alfieri, R.; Realdon, S.; Fassan, M.; Saggioro, D. Detection of LINE-1 hypomethylation in cfDNA of Esophageal Adenocarcinoma Patients. Int. J. Mol. Sci. 2020, 21, 1547. https://doi.org/10.3390/ijms21041547
Boldrin E, Curtarello M, Dallan M, Alfieri R, Realdon S, Fassan M, Saggioro D. Detection of LINE-1 hypomethylation in cfDNA of Esophageal Adenocarcinoma Patients. International Journal of Molecular Sciences. 2020; 21(4):1547. https://doi.org/10.3390/ijms21041547
Chicago/Turabian StyleBoldrin, Elisa, Matteo Curtarello, Marco Dallan, Rita Alfieri, Stefano Realdon, Matteo Fassan, and Daniela Saggioro. 2020. "Detection of LINE-1 hypomethylation in cfDNA of Esophageal Adenocarcinoma Patients" International Journal of Molecular Sciences 21, no. 4: 1547. https://doi.org/10.3390/ijms21041547
APA StyleBoldrin, E., Curtarello, M., Dallan, M., Alfieri, R., Realdon, S., Fassan, M., & Saggioro, D. (2020). Detection of LINE-1 hypomethylation in cfDNA of Esophageal Adenocarcinoma Patients. International Journal of Molecular Sciences, 21(4), 1547. https://doi.org/10.3390/ijms21041547