Secreted Factors and EV-miRNAs Orchestrate the Healing Capacity of Adipose Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
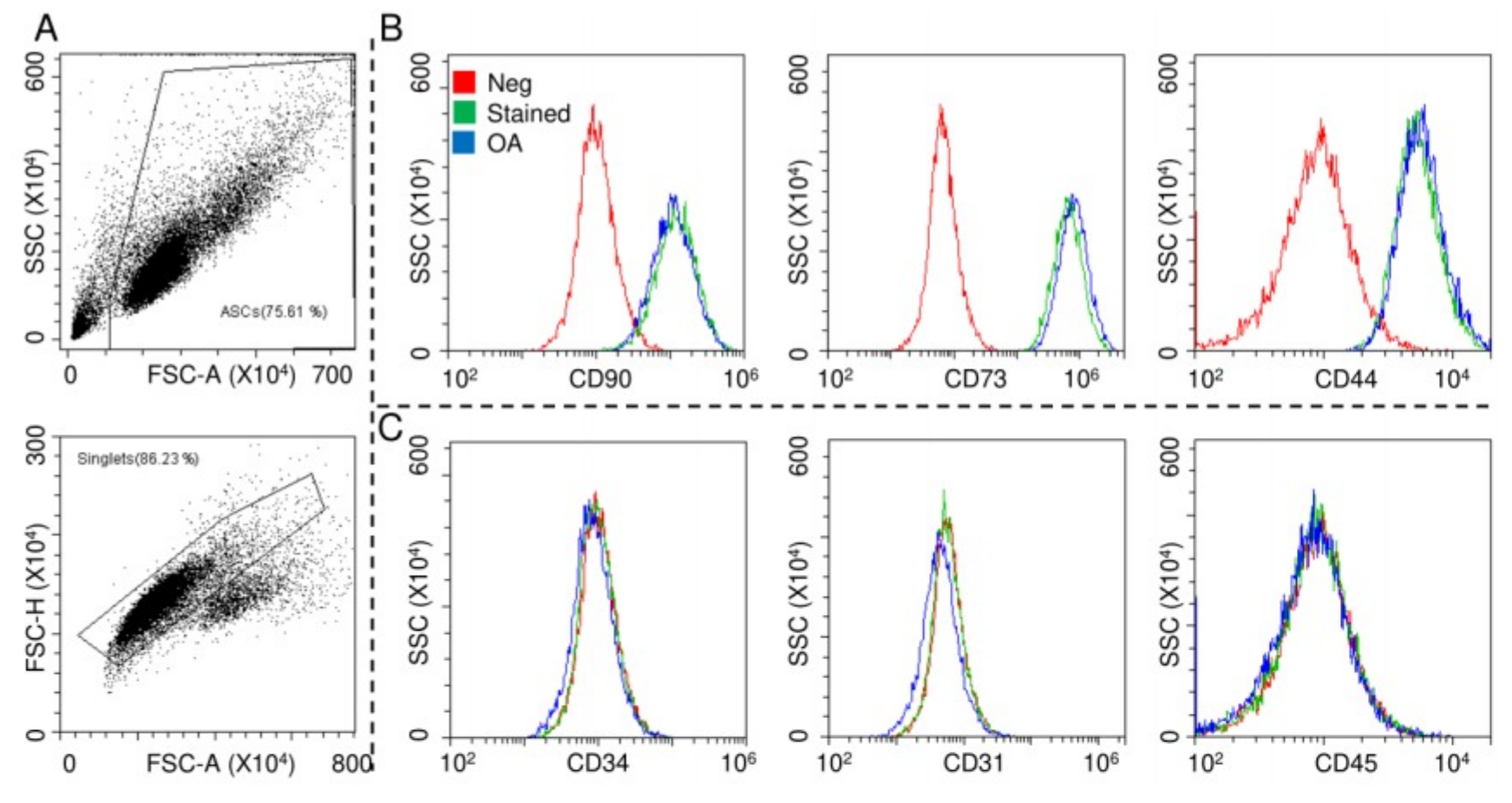
2.1. ASCs Characterization
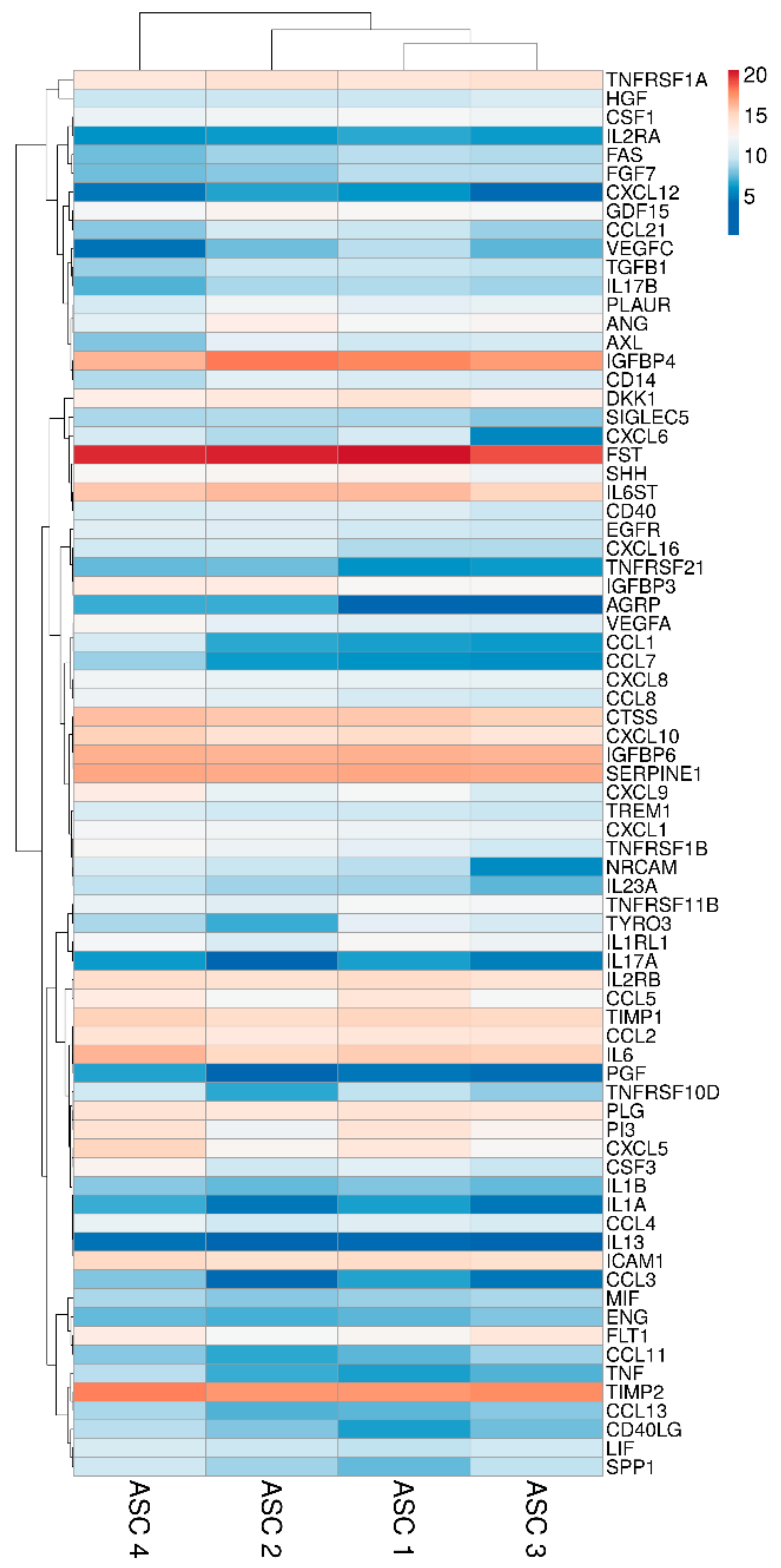
2.2. ASCs Secreted Factors
2.3. Characterization of ASC-Derived Extracellular Vesicles
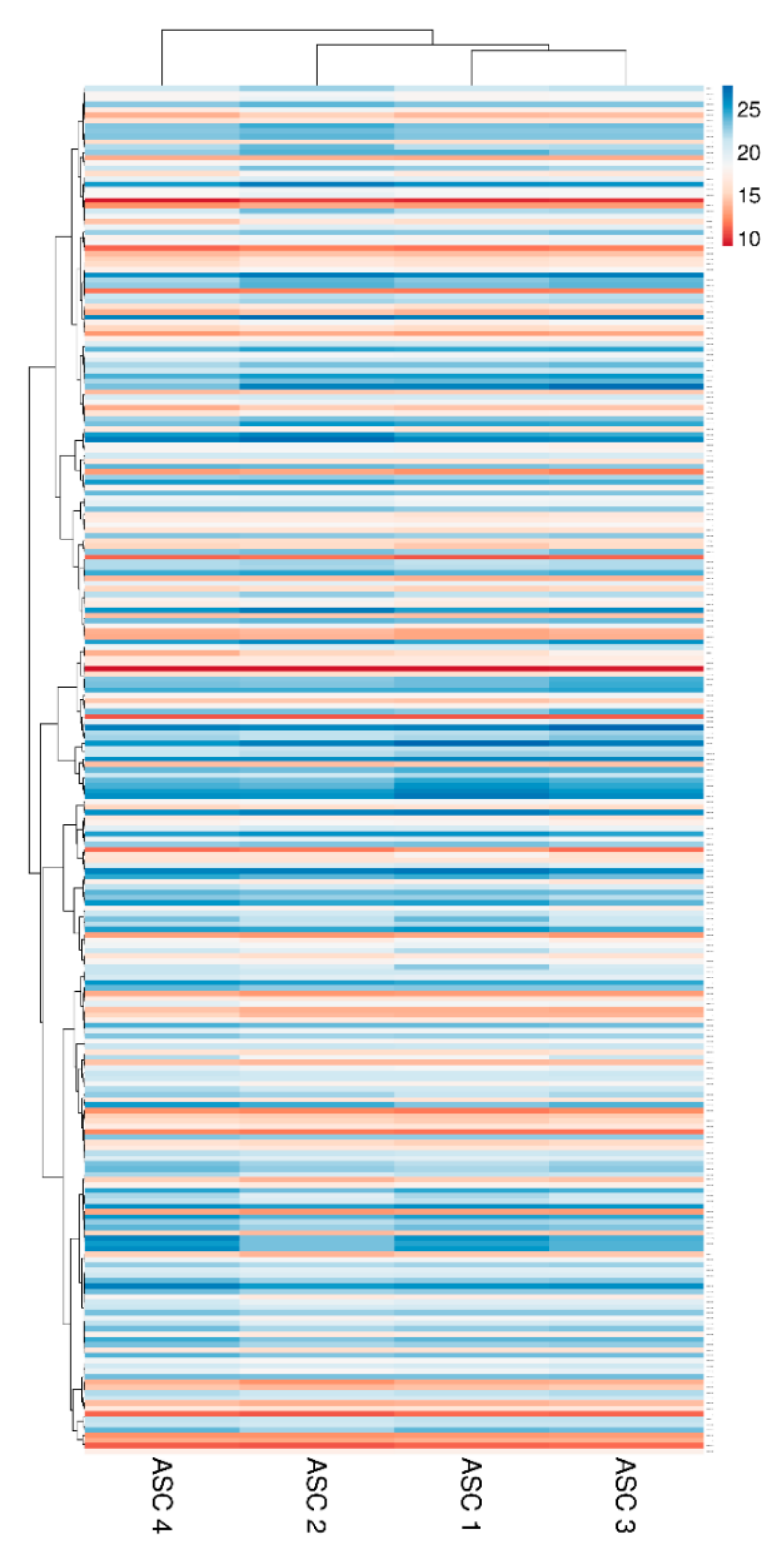
2.4. EV-Associated miRNAs
2.5. Target and Effect Prediction of EV-miRNAs on OA-Cartilage
2.6. Target and Effect Prediction of EV-miRNAs on Synovial Macrophages
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Adipose-Derived Mesenchymal Stem Cell (ASCs) Isolation and Expansion
4.3. ASCs Flow Cytometry Characterization
4.4. ELISA Assays
4.5. EVs Isolation and Characterization
- (i)
- Flow cytometry: Before ultracentrifugation, secretomes were additioned with 10 µM CFSE (Sigma-Aldrich) and incubated for 1 h at 37 °C. After ultracentrifugation, as previously described, labelled EVs were suspended in 50 µl PBS per 5 mL of processed medium. Particles were 1:10,000 diluted in PBS and 100 µl stained with anti-5 μL CD9-APC clone HI9a, anti-CD63-APC clone H5C6 and CD81-APC clone 5A6 (Biolegend, San Diego, CA, USA) for 30 min at 4 °C in the dark. Antibodies were used individually. Events collection was performed with a CytoFLEX flow cytometer collecting a minimum of 30,000 events. A reference bead mix (Biocytex, Marseille, France) composed of a FITC fluorescent mixture of spheres (100 nm, 300 nm, 500 nm and 900 nm) was used to set up the flow cytometer. Gains were: FSC = 106, SSC = 61, FITC = 272, PE = 116 and PC7 = 371. FITC threshold was set at 500 to include 100 nm beads and some smaller debris in the FITC channel.
- (ii)
- Transmission electron microscopy: Five microliters of PBS-suspended EVs were absorbed on Formvar carbon-coated grids for 10 min. Filter paper was used to blot drops. Negative stain was performed with 2% uranyl acetate aqueous suspension for 10 min, and excess was removed by filter paper before drying the grid at RT. Samples were examined with a TALOS L120C transmission electron microscope (Thermo Fisher Scientific, Waltham, MA, USA) at 120 kV.
- (iii)
- Nanoparticle tracking analysis (NTA): Purified EVs in PBS (1:100 diluted) were visualized by Nanosight LM10-HS system (NanoSight Ltd., Amesbury, UK). Three recordings of 30 s were performed for each sample. NTA software was used to analyze the data and provided both the concentration measurements and the high-resolution particle size distribution profiles.
4.6. Screening of EV-Embedded miRNA Expression
4.7. Pathway Analysis
- (i)
- Protein: Secretome-identified factors were subjected to functional enrichment analysis to provide insight into the functional associations of these protein subsets. The analysis was performed using Gorilla tool (http://cbl-gorilla.cs.technion.ac.il/) with two unranked lists of genes (target and background lists) running mode and 200 factors of the ELISA array as background [109,110]. The list of proteins was also submitted to the PANTHER web interface (http://www.pantherdb.org/) to identify proteins belonging to the same functional classifications, following default settings [111].
- (ii)
- miRNA: The predicted miRNA targets were annotated into functional BP using the microRNA Target Filter tool in ingenuity pathway analysis (IPA; Ingenuity® Systems, http://www.ingenuity.com). Filters were: confidence “experimentally observed”, with all sources available in the IPA database (TarBase, miRecords, TargetScan Human and Ingenuity Expert Findings). To obtain the list of all experimentally validated mRNA targets, no disease filter was set.
4.8. Hierarchical Clustering
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| OA | Osteoarthritis |
| MSC | Mesenchymal Stem Cells |
| EV | Extracellular Vesicles |
| miRNA | MicroRNA |
| HA | Hyaluronic Acid |
| ECM | Extracellular Matrix |
| GO | Gene Ontology |
| BP | Biological Process |
Appendix A
References
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef] [Green Version]
- Fu, K.; Robbins, S.R.; McDougall, J.J. Osteoarthritis: The genesis of pain. Rheumatology 2018, 57, iv43–iv50. [Google Scholar] [CrossRef] [Green Version]
- Loeser, R.F.; Collins, J.A.; Diekman, B.O. Ageing and the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 412–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, A.E.; Allen, K.D.; Golightly, Y.M.; Goode, A.P.; Jordan, J.M. A systematic review of recommendations and guidelines for the management of osteoarthritis: The chronic osteoarthritis management initiative of the US bone and joint initiative. Semin. Arthritis Rheum. 2014, 43, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Ferket, B.S.; Feldman, Z.; Zhou, J.; Oei, E.H.; Bierma-Zeinstra, S.M.; Mazumdar, M. Impact of total knee replacement practice: Cost effectiveness analysis of data from the Osteoarthritis Initiative. BMJ 2017, 356, j1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruyère, O.; Honvo, G.; Veronese, N.; Arden, N.K.; Branco, J.; Curtis, E.M.; Al-Daghri, N.M.; Herrero-Beaumont, G.; Martel-Pelletier, J.; Pelletier, J.P.; et al. An updated algorithm recommendation for the management of knee osteoarthritis from the European Society for Clinical and Economic Aspects of Osteoporosis, Osteoarthritis and Musculoskeletal Diseases (ESCEO). Semin. Arthritis Rheum. 2019, 49, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Skou, S.T.; Roos, E.M.; Laursen, M.B.; Rathleff, M.S.; Arendt-Nielsen, L.; Simonsen, O.; Rasmussen, S. A Randomized, Controlled Trial of Total Knee Replacement. N. Engl. J. Med. 2015, 373, 1597–1606. [Google Scholar] [CrossRef]
- Wylde, V.; Hewlett, S.; Learmonth, I.D.; Dieppe, P. Persistent pain after joint replacement: Prevalence, sensory qualities, and postoperative determinants. Pain 2011, 152, 566–572. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Manferdini, C.; Paolella, F.; Gabusi, E.; Gambari, L.; Piacentini, A.; Filardo, G.; Fleury-Cappellesso, S.; Barbero, A.; Murphy, M.; Lisignoli, G. Adipose stromal cells mediated switching of the pro-inflammatory profile of M1-like macrophages is facilitated by PGE2: In vitro evaluation. Osteoarthr. Cartil. 2017, 25, 1161–1171. [Google Scholar] [CrossRef] [Green Version]
- Ryan, A.; Murphy, M.; Barry, F. Mesenchymal stem/stromal cell therapy. In The Biology and Therapeutic Application of Mesenchymal Cells; Atkinson, K., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 426–440. [Google Scholar]
- Meisel, R.; Zibert, A.; Laryea, M.; Göbel, U.; Däubener, W.; Dilloo, D. Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood 2004, 103, 4619–4621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, W.; Jiang, J.; Arp, J.; Liu, W.; Garcia, B.; Wang, H. Regulatory T-cell generation and kidney allograft tolerance induced by mesenchymal stem cells associated with indoleamine 2,3-dioxygenase expression. Transplantation 2010, 90, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Khatab, S.; van Osch, G.J.; Kops, N.; Bastiaansen-Jenniskens, Y.M.; Bos, P.K.; Verhaar, J.A.; Bernsen, M.R.; van Buul, G.M. Mesenchymal stem cell secretome reduces pain and prevents cartilage damage in a murine osteoarthritis model. Eur. Cells Mater. 2018, 36, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chang, Y.W.; Tan, K.P.; Shen, Y.S.; Wang, Y.H.; Chang, C.H. Can mesenchymal stem cells and their conditioned medium assist inflammatory chondrocytes recovery? PLoS ONE 2018, 13, e0205563. [Google Scholar] [CrossRef] [PubMed]
- Van Buul, G.M.; Villafuertes, E.; Bos, P.K.; Waarsing, J.H.; Kops, N.; Narcisi, R.; Weinans, H.; Verhaar, J.A.; Bernsen, M.R.; Van Osch, G.J. Mesenchymal stem cells secrete factors that inhibit inflammatory processes in short-term osteoarthritic synovium and cartilage explant culture. Osteoarthr. Cartil. 2012, 20, 1186–1196. [Google Scholar] [CrossRef] [Green Version]
- Baker, K.; Grainger, A.; Niu, J.; Clancy, M.; Guermazi, A.; Crema, M.; Hughes, L.; Buckwalter, J.; Wooley, A.; Nevitt, M.; et al. Relation of synovitis to knee pain using contrast-enhanced MRIs. Ann. Rheum. Dis. 2010, 69, 1779–1783. [Google Scholar] [CrossRef] [Green Version]
- Mathiessen, A.; Conaghan, P.G. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [Green Version]
- Benito, M.J.; Veale, D.J.; FitzGerald, O.; van den Berg, W.B.; Bresnihan, B. Synovial tissue inflammation in early and late osteoarthritis. Ann. Rheum. Dis. 2005, 64, 1263–1267. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.W.; Wang, J.; Lee, C.J.; Liu, M.; Neelamegham, S.; Canty, J.M.; Nguyen, J. The microRNA regulatory landscape of MSC-derived exosomes: A systems view. Sci. Rep. 2018, 8, 1419. [Google Scholar] [CrossRef]
- Nawaz, M.; Fatima, F.; Vallabhaneni, K.C.; Penfornis, P.; Valadi, H.; Ekström, K.; Kholia, S.; Whitt, J.D.; Fernandes, J.D.; Pochampally, R.; et al. Extracellular Vesicles: Evolving Factors in Stem Cell Biology. Stem Cells Int. 2016, 2016, 1073140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonk, L.A.; van Dooremalen, S.F.J.; Liv, N.; Klumperman, J.; Coffer, P.J.; Saris, D.B.F.; Lorenowicz, M.J. Mesenchymal Stromal/stem Cell-derived Extracellular Vesicles Promote Human Cartilage Regeneration In Vitro. Theranostics 2018, 8, 906–920. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Perucca Orfei, C.; De Luca, P.; Lugano, G.; Viganò, M.; Colombini, A.; Valli, F.; Zacchetti, D.; Bollati, V.; de Girolamo, L. Interaction with hyaluronan matrix and miRNA cargo as contributors for in vitro potential of mesenchymal stem cell-derived extracellular vesicles in a model of human osteoarthritic synoviocytes. Stem Cell Res. Ther. 2019, 10, 109. [Google Scholar] [CrossRef]
- Tao, S.C.; Yuan, T.; Zhang, Y.L.; Yin, W.J.; Guo, S.C.; Zhang, C.Q. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics 2017, 7, 180–195. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Zhao, B.; Niu, X.; Hu, B.; Li, Q.; Zhang, J.; Ding, J.; Chen, Y.; Wang, Y. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res. Ther. 2017, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci. Rep. 2017, 7, 16214. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kuang, L.; Chen, C.; Yang, J.; Zeng, W.N.; Li, T.; Chen, H.; Huang, S.; Fu, Z.; Li, J.; et al. miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials 2019, 206, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Zavatti, M.; Beretti, F.; Casciaro, F.; Bertucci, E.; Maraldi, T. Comparison of the therapeutic effect of amniotic fluid stem cells and their exosomes on monoiodoacetate-induced animal model of osteoarthritis. Biofactors 2019. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Kim, H.S.; Hong, I.S. Stem Cell-Derived Extracellular Vesicles as Immunomodulatory Therapeutics. Stem Cells Int. 2019, 2019, 5126156. [Google Scholar] [CrossRef]
- Sun, M.; Sun, L.; Huang, C.; Chen, B.C.; Zhou, Z. Induction of Macrophage M2b/c Polarization by Adipose Tissue-Derived Mesenchymal Stem Cells. J. Immunol. Res. 2019, 2019, 7059680. [Google Scholar] [CrossRef] [Green Version]
- Paolella, F.; Manferdini, C.; Gabusi, E.; Gambari, L.; Filardo, G.; Kon, E.; Mariani, E.; Lisignoli, G. Effect of microfragmented adipose tissue on osteoarthritic synovial macrophage factors. J. Cell Physiol. 2019, 234, 5044–5055. [Google Scholar] [CrossRef] [PubMed]
- Kahle, P.; Saal, J.G.; Schaudt, K.; Zacher, J.; Fritz, P.; Pawelec, G. Determination of cytokines in synovial fluids: Correlation with diagnosis and histomorphological characteristics of synovial tissue. Ann. Rheum. Dis. 1992, 51, 731–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westacott, C.I.; Whicher, J.T.; Barnes, I.C.; Thompson, D.; Swan, A.J.; Dieppe, P.A. Synovial fluid concentration of five different cytokines in rheumatic diseases. Ann. Rheum. Dis. 1990, 49, 676–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchida, A.I.; Beekhuizen, M.; Ct Hart, M.; Radstake, T.R.; Dhert, W.J.; Saris, D.B.; van Osch, G.J.; Creemers, L.B. Cytokine profiles in the joint depend on pathology, but are different between synovial fluid, cartilage tissue and cultured chondrocytes. Arthritis Res. Ther. 2014, 16, 441. [Google Scholar] [CrossRef] [Green Version]
- Mabey, T.; Honsawek, S.; Tanavalee, A.; Yuktanandana, P.; Wilairatana, V.; Poovorawan, Y. Plasma and synovial fluid inflammatory cytokine profiles in primary knee osteoarthritis. Biomarkers 2016, 21, 639–644. [Google Scholar] [CrossRef]
- Siqueira, M.B.; Frangiamore, S.; Klika, A.K.; Gajewski, N.; Barsoum, W.K.; Higuera, C.A. Comparison of Synovial Fluid Cytokine Levels between Traumatic Knee Injury and End-Stage Osteoarthritis. J. Knee Surg. 2017, 30, 128–133. [Google Scholar] [CrossRef]
- Vangsness, C.T., Jr.; Burke, W.S.; Narvy, S.J.; MacPhee, R.D.; Fedenko, A.N. Human knee synovial fluid cytokines correlated with grade of knee osteoarthritis—A pilot study. Bull. NYU Hosp. Jt. Dis. 2011, 69, 122–127. [Google Scholar]
- Orita, S.; Koshi, T.; Mitsuka, T.; Miyagi, M.; Inoue, G.; Arai, G.; Ishikawa, T.; Hanaoka, E.; Yamashita, K.; Yamashita, M.; et al. Associations between proinflammatory cytokines in the synovial fluid and radiographic grading and pain-related scores in 47 consecutive patients with osteoarthritis of the knee. BMC Musculoskelet. Disord. 2011, 12, 144. [Google Scholar] [CrossRef] [Green Version]
- Kokebie, R.; Aggarwal, R.; Lidder, S.; Hakimiyan, A.A.; Rueger, D.C.; Block, J.A.; Chubinskaya, S. The role of synovial fluid markers of catabolism and anabolism in osteoarthritis, rheumatoid arthritis and asymptomatic organ donors. Arthritis Res. Ther. 2011, 13, R50. [Google Scholar] [CrossRef] [Green Version]
- Scanzello, C.R.; Umoh, E.; Pessler, F.; Diaz-Torne, C.; Miles, T.; Dicarlo, E.; Potter, H.G.; Mandl, L.; Marx, R.; Rodeo, S.; et al. Local cytokine profiles in knee osteoarthritis: Elevated synovial fluid interleukin-15 differentiates early from end-stage disease. Osteoarthr. Cartil. 2009, 17, 1040–1048. [Google Scholar] [CrossRef] [Green Version]
- Toh, W.S.; Lai, R.C.; Zhang, B.; Lim, S.K. MSC exosome works through a protein-based mechanism of action. Biochem. Soc. Trans. 2018, 46, 843–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevillet, J.R.; Kang, Q.; Ruf, I.K.; Briggs, H.A.; Vojtech, L.N.; Hughes, S.M.; Cheng, H.H.; Arroyo, J.D.; Meredith, E.K.; Gallichotte, E.N.; et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc. Natl. Acad. Sci. USA 2014, 111, 14888–14893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisch, K.M.; Gamini, R.; Alvarez-Garcia, O.; Akagi, R.; Saito, M.; Muramatsu, Y.; Sasho, T.; Koziol, J.A.; Su, A.I.; Lotz, M.K. Identification of transcription factors responsible for dysregulated networks in human osteoarthritis cartilage by global gene expression analysis. Osteoarthr. Cartil. 2018, 26, 1531–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endisha, H.; Rockel, J.; Jurisica, I.; Kapoor, M. The complex landscape of microRNAs in articular cartilage: Biology, pathology, and therapeutic targets. JCI Insight 2018, 3, e121630. [Google Scholar] [CrossRef] [Green Version]
- Wood, M.J.; Leckenby, A.; Reynolds, G.; Spiering, R.; Pratt, A.G.; Rankin, K.S.; Isaacs, J.D.; Haniffa, M.A.; Milling, S.; Hilkens, C.M. Macrophage proliferation distinguishes 2 subgroups of knee osteoarthritis patients. JCI Insight 2019, 4, e125325. [Google Scholar] [CrossRef] [Green Version]
- Mandelin, A.M.; Homan, P.J.; Shaffer, A.M.; Cuda, C.M.; Dominguez, S.T.; Bacalao, E.; Carns, M.; Hinchcliff, M.; Lee, J.; Aren, K.; et al. Transcriptional Profiling of Synovial Macrophages Using Minimally Invasive Ultrasound-Guided Synovial Biopsies in Rheumatoid Arthritis. Arthritis Rheumatol. 2018, 70, 841–854. [Google Scholar] [CrossRef]
- Xu, S.J.; Hu, H.T.; Li, H.L.; Chang, S. The Role of miRNAs in Immune Cell Development, Immune Cell Activation, and Tumor Immunity: With a Focus on Macrophages and Natural Killer Cells. Cells 2019, 8, 1140. [Google Scholar] [CrossRef] [Green Version]
- Kitade, H.; Sawamoto, K.; Nagashimada, M.; Inoue, H.; Yamamoto, Y.; Sai, Y.; Takamura, T.; Yamamoto, H.; Miyamoto, K.; Ginsberg, H.N.; et al. CCR5 plays a critical role in obesity-induced adipose tissue inflammation and insulin resistance by regulating both macrophage recruitment and M1/M2 status. Diabetes 2012, 61, 1680–1690. [Google Scholar] [CrossRef] [Green Version]
- Toh, W.S.; Lai, R.C.; Hui, J.H.P.; Lim, S.K. MSC exosome as a cell-free MSC therapy for cartilage regeneration: Implications for osteoarthritis treatment. Semin. Cell Dev. Biol. 2017, 67, 56–64. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, Y.; Zhang, J.; Yuan, Y.; Wang, J. Exosomes: A Novel Therapeutic Agent for Cartilage and Bone Tissue Regeneration. Dose Response 2019, 17, 1559325819892702. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, D.; Liu, Z.; Zhou, F.; Dai, J.; Wu, B.; Zhou, J.; Heng, B.C.; Zou, X.H.; Ouyang, H.; et al. Exosomes from embryonic mesenchymal stem cells alleviate osteoarthritis through balancing synthesis and degradation of cartilage extracellular matrix. Stem Cell Res. Ther. 2017, 8, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.; Shah, N.; Zanetti, B.R.; Maugeri, M.; Silvestre, R.N.; Fatima, F.; Neder, L.; Valadi, H. Extracellular Vesicles and Matrix Remodeling Enzymes: The Emerging Roles in Extracellular Matrix Remodeling, Progression of Diseases and Tissue Repair. Cells 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, C.H.; Lee, Y.G.; Shin, W.H.; Kim, H.; Chai, J.W.; Jeong, E.C.; Kim, J.E.; Shim, H.; Shin, J.S.; Shin, I.S.; et al. Intra-articular injection of mesenchymal stem cells for the treatment of osteoarthritis of the knee: A proof-of-concept clinical trial. Stem Cells 2014, 32, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Pers, Y.M.; Rackwitz, L.; Ferreira, R.; Pullig, O.; Delfour, C.; Barry, F.; Sensebe, L.; Casteilla, L.; Fleury, S.; Bourin, P.; et al. ADIPOA Consortium. Adipose Mesenchymal Stromal Cell-Based Therapy for Severe Osteoarthritis of the Knee: A Phase I Dose-Escalation Trial. Stem Cells Transl. Med. 2016, 5, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Di Matteo, B.; Vandenbulcke, F.; Vitale, N.D.; Iacono, F.; Ashmore, K.; Marcacci, M.; Kon, E. Minimally Manipulated Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis: A Systematic Review of Clinical Evidence. Stem Cells Int. 2019, 2019, 1735242. [Google Scholar] [CrossRef] [Green Version]
- Nava, S.; Sordi, V.; Pascucci, L.; Tremolada, C.; Ciusani, E.; Zeira, O.; Cadei, M.; Soldati, G.; Pessina, A.; Parati, E.; et al. Long-Lasting Anti-Inflammatory Activity of Human Microfragmented Adipose Tissue. Stem Cells Int. 2019, 2019, 5901479. [Google Scholar] [CrossRef]
- Mancuso, P.; Raman, S.; Glynn, A.; Barry, F.; Murphy, J.M. Mesenchymal Stem Cell Therapy for Osteoarthritis: The Critical Role of the Cell Secretome. Front. Bioeng. Biotechnol. 2019, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- D’Arrigo, D.; Roffi, A.; Cucchiarini, M.; Moretti, M.; Candrian, C.; Filardo, G. Secretome and Extracellular Vesicles as New Biological Therapies for Knee Osteoarthritis: A Systematic Review. J. Clin. Med. 2019, 8, 1867. [Google Scholar] [CrossRef] [Green Version]
- Castañeda, S.; Vicente, E.F. Osteoarthritis: More than Cartilage Degeneration. Clin. Rev. Bone Miner. Metab. 2017, 15, 69–81. [Google Scholar] [CrossRef]
- Borić, I.; Hudetz, D.; Rod, E.; Jeleč, Ž.; Vrdoljak, T.; Skelin, A.; Polašek, O.; Plečko, M.; Trbojević-Akmačić, I.; Lauc, G.; et al. A 24-Month Follow-Up Study of the Effect of Intra-Articular Injection of Autologous Microfragmented Fat Tissue on Proteoglycan Synthesis in Patients with Knee Osteoarthritis. Genes 2019, 10, 1051. [Google Scholar] [CrossRef] [Green Version]
- Bansal, H.; Comella, K.; Leon, J.; Verma, P.; Agrawal, D.; Koka, P.; Ichim, T. Intra-articular injection in the knee of adipose derived stromal cells (stromal vascular fraction) and platelet rich plasma for osteoarthritis. J. Transl. Med. 2017, 15, 141. [Google Scholar] [CrossRef] [Green Version]
- Pham, P.V.; Bui, K.H.-T.; Duong, T.D.; Nguyen, N.T.; Nguyen, T.D.; Le, V.T.; Mai, V.T.; Phan, N.L.-C.; Le, D.M.; Ngoc, N.M. Symptomatic knee osteoarthritis treatment using autologous adipose derived stem cells and platelet-rich plasma: A clinical study. Biomed. Res. Ther. 2014, 1, 2. [Google Scholar] [CrossRef]
- Hong, Z.; Chen, J.; Zhang, S.; Zhao, C.; Bi, M.; Chen, X.; Bi, Q. Intra-articular injection of autologous adipose-derived stromal vascular fractions for knee osteoarthritis: A double-blind randomized self-controlled trial. Int. Orthop. 2019, 43, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.G.; Jo, S.B.; Kwon, O.R.; Suh, D.S.; Lee, S.W.; Park, S.H.; Choi, Y.J. Mesenchymal stem cell injections improve symptoms of knee osteoarthritis. Arthroscopy 2013, 29, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Du, H.; Dai, C.; Zhang, L.; Li, S.; Hunter, D.J.; Lu, L.; Bao, C. Human adipose-derived mesenchymal stem cells for osteoarthritis: A pilot study with long-term follow-up and repeated injections. Regen. Med. 2018, 13, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.H.; Chai, J.W.; Jeong, E.C.; Oh, S.; Shin, J.S.; Shim, H.; Yoon, K.S. Intra-articular Injection of Mesenchymal Stem Cells for the Treatment of Osteoarthritis of the Knee: A 2-Year Follow-up Study. Am. J. Sports Med. 2017, 45, 2774–2783. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.D.; Martel-Pelletier, J.; Pelletier, J.P.; Howell, D.S.; Woessner, J.F., Jr. Evidence for metalloproteinase and metalloproteinase inhibitor imbalance in human osteoarthritic cartilage. J. Clin. Investig. 1989, 84, 678–685. [Google Scholar] [CrossRef] [Green Version]
- Clements, D.N.; Fitzpatrick, N.; Carter, S.D.; Day, P.J. Cartilage gene expression correlates with radiographic severity of canine elbow osteoarthritis. Vet. J. 2009, 179, 211–218. [Google Scholar] [CrossRef]
- Alam, M.R.; Ji, J.R.; Kim, M.S.; Kim, N.S. Biomarkers for identifying the early phases of osteoarthritis secondary to medial patellar luxation in dogs. J. Vet. Sci. 2011, 12, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Mi, M.; Shi, S.; Li, T.; Holz, J.; Lee, Y.J.; Sheu, T.J.; Liao, Q.; Xiao, T. TIMP2 deficient mice develop accelerated osteoarthritis via promotion of angiogenesis upon destabilization of the medial meniscus. Biochem. Biophys. Res. Commun. 2012, 423, 366–372. [Google Scholar] [CrossRef]
- Yamada, J.; Tsuji, K.; Miyatake, K.; Matsukura, Y.; Abula, K.; Inoue, M.; Sekiya, I.; Muneta, T. Follistatin alleviates synovitis and articular cartilage degeneration induced by carrageenan. Int. J. Inflam. 2014, 2014, 959271. [Google Scholar] [CrossRef]
- Li, J.; Ny, A.; Leonardsson, G.; Nandakumar, K.S.; Holmdahl, R.; Ny, T. The plasminogen activator/plasmin system is essential for development of the joint inflammatory phase of collagen type II-induced arthritis. Am. J. Pathol. 2005, 166, 783–792. [Google Scholar] [CrossRef] [Green Version]
- Blanco Garcia, F.J. Catabolic events in osteoarthritic cartilage. Osteoarthr. Cartil. 1999, 7, 308–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreier, R.; Wallace, S.; Fuchs, S.; Bruckner, P.; Grässel, S. Paracrine interactions of chondrocytes and macrophages in cartilage degradation: Articular chondrocytes provide factors that activate macrophage-derived pro-gelatinase B (pro-MMP-9). J. Cell. Sci. 2001, 114, 3813–3822. [Google Scholar] [PubMed]
- Alejandre Alcazar, M.A.; Kaschwich, M.; Ertsey, R.; Preuss, S.; Milla, C.; Mujahid, S.; Masumi, J.; Khan, S.; Mokres, L.M.; Tian, L.; et al. Elafin Treatment Rescues EGFR-Klf4 Signaling and Lung Cell Survival in Ventilated Newborn Mice. Am. J. Respir. Cell Mol. Biol. 2018, 59, 623–634. [Google Scholar] [CrossRef]
- McQuillan, D.J.; Handley, C.J.; Campbell, M.A.; Bolis, S.; Milway, V.E.; Herington, A.C. Stimulation of proteoglycan biosynthesis by serum and insulin-like growth factor-I in cultured bovine articular cartilage. Biochem. J. 1986, 240, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Ekenstedt, K.J.; Sonntag, W.E.; Loeser, R.F.; Lindgren, B.R.; Carlson, C.S. Effects of chronic growth hormone and insulin-like growth factor 1 deficiency on osteoarthritis severity in rat knee joints. Arthritis Rheum. 2006, 54, 3850–3858. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.Y.; Lee, J.K.; Wei, L.; Qu, F.; Zhang, J.Z. Correlation of insulin-like growth factor 1 and osteoarthritic cartilage degradation: A spontaneous osteoarthritis in guinea-pig. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4493–4500. [Google Scholar]
- Hou, W.S.; Li, Z.; Büttner, F.H.; Bartnik, E.; Brömme, D. Cleavage site specificity of cathepsin K toward cartilage proteoglycans and protease complex formation. Biol. Chem. 2003, 384, 891–897. [Google Scholar] [CrossRef]
- Oh, H.; Chun, C.H.; Chun, J.S. Dkk-1 expression in chondrocytes inhibits experimental osteoarthritic cartilage destruction in mice. Arthritis Rheum. 2012, 64, 2568–2578. [Google Scholar] [CrossRef]
- Weng, L.H.; Wang, C.J.; Ko, J.Y.; Sun, Y.C.; Wang, F.S. Control of Dkk-1 ameliorates chondrocyte apoptosis, cartilage destruction, and subchondral bone deterioration in osteoarthritic knees. Arthritis Rheum. 2010, 62, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- De Santis, M.; Di Matteo, B.; Chisari, E.; Cincinelli, G.; Angele, P.; Lattermann, C.; Filardo, G.; Vitale, N.D.; Selmi, C.; Kon, E. The Role of Wnt Pathway in the Pathogenesis of OA and Its Potential Therapeutic Implications in the Field of Regenerative Medicine. Biomed. Res. Int. 2018, 2018, 7402947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuolteenaho, K.; Koskinen-Kolasa, A.; Laavola, M.; Nieminen, R.; Moilanen, T.; Moilanen, E. High synovial fluid interleukin-6 levels are associated with increased matrix metalloproteinase levels and severe radiographic changes in osteoarthritis patients. Osteoarthr. Cartil. 2017, 25, S92–S93. [Google Scholar] [CrossRef]
- Miller, R.E.; Miller, R.J.; Malfait, A.M. Osteoarthritis joint pain: The cytokine connection. Cytokine 2014, 70, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Finnson, K.W.; Chi, Y.; Bou-Gharios, G.; Leask, A.; Philip, A. TGF-b signaling in cartilage homeostasis and osteoarthritis. Front. Biosci. 2012, 4, 251–268. [Google Scholar] [CrossRef]
- Blaney Davidson, E.N.; Remst, D.F.; Vitters, E.L.; van Beuningen, H.M.; Blom, A.B.; Goumans, M.J.; van den Berg, W.B.; van der Kraan, P.M. Increase in ALK1/ALK5 ratio as a cause for elevated MMP-13 expression in osteoarthritis in humans and mice. J. Immunol. 2009, 182, 7937–7945. [Google Scholar] [CrossRef] [Green Version]
- Van Beuningen, H.M.; Glansbeek, H.L.; van der Kraan, P.M.; van den Berg, W.B. Osteoarthritis-like changes in the murine knee joint resulting from intra-articular transforming growth factor-beta injections. Osteoarthr. Cartil. 2000, 8, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Kouri, J.B.; Jiménez, S.A.; Quintero, M.; Chico, A. Ultrastructural study of chondrocytes from fibrillated and non-fibrillated human osteoarthritic cartilage. Osteoarthr. Cartil. 1996, 4, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Philipot, D.; Guérit, D.; Platano, D.; Chuchana, P.; Olivotto, E.; Espinoza, F.; Dorandeu, A.; Pers, Y.M.; Piette, J.; Borzi, R.M.; et al. p16INK4a and its regulator miR-24 link senescence and chondrocyte terminal differentiation-associated matrix remodeling in osteoarthritis. Arthritis Res. Ther. 2014, 16, R58. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.H.; Liu, W.; Zhang, L.; Liu, X.Y.; Wang, Y.; Xue, B.; Liu, B.; Duan, R.; Zhang, B.; Ji, Y. Effects of microRNA-24 targeting C-myc on apoptosis, proliferation, and cytokine expressions in chondrocytes of rats with osteoarthritis via MAPK signaling pathway. J. Cell. Biochem. 2018, 119, 7944–7958. [Google Scholar] [CrossRef]
- Benigni, G.; Dimitrova, P.; Antonangeli, F.; Sanseviero, E.; Milanova, V.; Blom, A.; van Lent, P.; Morrone, S.; Santoni, A.; Bernardini, G. CXCR3/CXCL10 Axis Regulates Neutrophil-NK Cell Cross-Talk Determining the Severity of Experimental Osteoarthritis. J. Immunol. 2017, 198, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Endres, M.; Andreas, K.; Kalwitz, G.; Freymann, U.; Neumann, K.; Ringe, J.; Sittinger, M.; Häupl, T.; Kaps, C. Chemokine profile of synovial fluid from normal, osteoarthritis and rheumatoid arthritis patients: CCL25, CXCL10 and XCL1 recruit human subchondral mesenchymal progenitor cells. Osteoarthr. Cartil. 2010, 18, 1458–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponte, A.L.; Marais, E.; Gallay, N.; Langonné, A.; Delorme, B.; Hérault, O.; Charbord, P.; Domenech, J. The in vitro migration capacity of human bone marrow mesenchymal stem cells: Comparison of chemokine and growth factor chemotactic activities. Stem Cells 2007, 25, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Ringe, J.; Strassburg, S.; Neumann, K.; Endres, M.; Notter, M.; Burmester, G.R.; Kaps, C.; Sittinger, M. Towards in situ tissue repair: Human mesenchymal stem cells express chemokine receptors CXCR1, CXCR2 and CCR2, and migrate upon stimulation with CXCL8 but not CCL2. J. Cell. Biochem. 2007, 101, 135–146. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Aristizábal, A.; Sharma, A.; Bakooshli, M.A.; Kapoor, M.; Gilbert, P.M.; Viswanathan, S.; Gandhi, R. Stage-specific differences in secretory profile of mesenchymal stromal cells (MSCs) subjected to early- vs late-stage OA synovial fluid. Osteoarthr. Cartil. 2017, 25, 737–741. [Google Scholar] [CrossRef] [Green Version]
- Sierra-Filardi, E.; Nieto, C.; Domínguez-Soto, A.; Barroso, R.; Sánchez-Mateos, P.; Puig-Kroger, A.; López-Bravo, M.; Joven, J.; Ardavín, C.; Rodríguez-Fernández, J.L.; et al. CCL2 shapes macrophage polarization by GM-CSF and M-CSF: Identification of CCL2/CCR2-dependent gene expression profile. J. Immunol. 2014, 192, 3858–3867. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.I.; Kim, M.R.; Jeong, H.Y.; Jeong, H.C.; Jeong, M.H.; Yoon, S.H.; Kim, Y.S.; Ahn, Y. Mesenchymal stem cells reciprocally regulate the M1/M2 balance in mouse bone marrow-derived macrophages. Exp. Mol. Med. 2014, 46, e70. [Google Scholar] [CrossRef]
- Zhang, Y.; Grosfeld, E.C.; Camargo, W.A.; Tang, H.; Magri, A.M.P.; van den Beucken, J.J.J.P. Efficacy of intraoperatively prepared cell-based constructs for bone regeneration. Stem Cell Res. Ther. 2018, 9, 283. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [Green Version]
- Self-Fordham, J.B.; Naqvi, A.R.; Uttamani, J.R.; Kulkarni, V.; Nares, S. MicroRNA: Dynamic Regulators of Macrophage Polarization and Plasticity. Front. Immunol. 2017, 8, 1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fordham, J.B.; Naqvi, A.R.; Nares, S. Regulation of miR-24, miR-30b, and miR-142-3p during macrophage and dendritic cell differentiation potentiates innate immunity. J. Leukoc. Biol. 2015, 98, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naqvi, A.R.; Fordham, J.B.; Nares, S. miR-24, miR-30b, and miR-142-3p regulate phagocytosis in myeloid inflammatory cells. J. Immunol. 2015, 194, 1916–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, X.; Wu, Q.; Wu, X.; Zhu, Q.; Wang, X.; Jiang, L.; Chen, X.; Wang, X. Epithelial ovarian cancer-secreted exosomal miR-222-3p induces polarization of tumor-associated macrophages. Oncotarget 2016, 7, 43076–43087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.L.; McNeill, J.; Goon, K.; Little, D.; Kimmerling, K.; Huebner, J.; Kraus, V.; Guilak, F. Conditional Macrophage Depletion Increases Inflammation and Does Not Inhibit the Development of Osteoarthritis in Obese Macrophage Fas-Induced Apoptosis-Transgenic Mice. Arthritis Rheumatol. 2017, 69, 1772–1783. [Google Scholar] [CrossRef] [PubMed]
- Pergoli, L.; Cantone, L.; Favero, C.; Angelici, L.; Iodice, S.; Pinatel, E.; Hoxha, M.; Dioni, L.; Letizia, M.; Albetti, B.; et al. Extracellular vesicle-packaged miRNA release after short-term exposure to particulate matter is associated with increased coagulation. Part. Fibre Toxicol. 2017, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- D’haene, B.; Mestdagh, P.; Hellemans, J.; Vandesompele, J. miRNA expression profiling: From reference genes to global mean normalization. Methods Mol. Biol. 2012, 822, 261–272. [Google Scholar] [CrossRef]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinform. 2009, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Eden, E.; Lipson, D.; Yogev, S.; Yakhini, Z. Discovering motifs in ranked lists of DNA sequences. PLoS Comput. Biol. 2007, 3, e39. [Google Scholar] [CrossRef]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FACTOR | ASC 1 | ASC 2 | ASC 3 | ASC 4 | MEAN | SD | DESCRIPTION |
|---|---|---|---|---|---|---|---|
| FST | 1,492,673 | 1,081,479 | 570,416 | 1,004,932 | 1,037,375 | 377,865 | Follistatin |
| TIMP2 | 167,790 | 162,350 | 198,481 | 256,519 | 196,285 | 43,191 | Metalloproteinase inhibitor 2 |
| IGFBP4 | 223,536 | 276,754 | 145,198 | 84,223 | 182,428 | 84,887 | Insulin-like growth factor-binding protein 4 |
| SERPINE1 | 109,335 | 103,661 | 98,032 | 119,475 | 10,7626 | 9148 | Plasminogen activator inhibitor 1 |
| IGFBP6 | 86,610 | 83,502 | 77,842 | 94,332 | 85,572 | 6877 | Insulin-like growth factor-binding protein 6 |
| IL6ST | 68,647 | 68,361 | 36,585 | 50,182 | 55,944 | 15,530 | Interleukin-6 receptor subunit beta |
| CTSS | 52,275 | 50,380 | 40,226 | 64,808 | 51,922 | 10,089 | Cathepsin S |
| IL6 | 43,025 | 30,591 | 40,435 | 82,769 | 49,205 | 23,008 | Interleukin-6 |
| TIMP1 | 34,985 | 27,024 | 33,210 | 41,839 | 34,265 | 6094 | Metalloproteinase inhibitor 1 |
| ICAM1 | 28,715 | 21,857 | 22,377 | 33,311 | 26,565 | 5472 | Intercellular adhesion molecule 1 |
| CXCL10 | 28,629 | 21,386 | 16,094 | 38,665 | 26,193 | 9774 | C-X-C motif chemokine 10 |
| IL2RB | 28,885 | 20,962 | 18,101 | 27,057 | 23,751 | 5066 | Interleukin-2 receptor subunit beta |
| TNFRSF1A | 15,652 | 21,277 | 21,328 | 13,791 | 18,012 | 3875 | Tumor necrosis factor receptor superfamily member 1A |
| CCL2 | 16,286 | 13,411 | 15,771 | 19,076 | 16,136 | 2325 | C-C motif chemokine 2 |
| PLG | 17,218 | 14,055 | 14,936 | 17,793 | 16,001 | 1790 | Plasminogen |
| CXCL5 | 15,226 | 5628 | 5204 | 36,614 | 15,668 | 14,711 | C-X-C motif chemokine 5 |
| DKK1 | 17,275 | 12,422 | 8703 | 9540 | 11,985 | 3870 | Dickkopf-related protein 1 |
| PI3 | 18,953 | 3218 | 6567 | 20,207 | 12,236 | 8605 | Elafin (Peptidase inhibitor 3) |
| CCL5 | 15,611 | 4502 | 4407 | 11,970 | 9122 | 5591 | C-C motif chemokine 5 |
| FLT1 | 5604 | 4854 | 13,916 | 10,834 | 8802 | 4324 | Vascular endothelial growth factor receptor 1 |
| IGFBP3 | 4898 | 11,609 | 5145 | 11,349 | 8250 | 3731 | Insulin-like growth factor-binding protein 3 |
| ANG | 4612 | 8990 | 5615 | 2158 | 5344 | 2831 | Angiogenin |
| SHH | 6957 | 5751 | 3445 | 5437 | 5398 | 1457 | Sonic hedgehog protein |
| GDF15 | 5128 | 6376 | 4534 | 4167 | 5051 | 968 | Growth/differentiation factor 15 |
| CXCL9 | 4391 | 2472 | 1272 | 10,677 | 4703 | 4185 | C-X-C motif chemokine 9 |
| CSF1 | 4488 | 3510 | 3624 | 2797 | 3604 | 693 | Macrophage colony-stimulating factor 1 |
| CXCL1 | 3334 | 3781 | 2516 | 4276 | 3477 | 747 | Growth-regulated alpha protein |
| IL1RL1 | 5212 | 1440 | 3249 | 3991 | 3473 | 1579 | Interleukin-1 receptor-like 1 |
| TNFRSF11B | 4378 | 1861 | 4234 | 2934 | 3352 | 1187 | Tumor necrosis factor receptor superfamily member 11B |
| CXCL8 | 2521 | 3046 | 2715 | 3839 | 3030 | 581 | Interleukin-8 |
| TNFRSF1B | 2277 | 3327 | 1014 | 5025 | 2911 | 1697 | Tumor necrosis factor receptor superfamily member 1B |
| VEGFA | 1956 | 2378 | 1738 | 5600 | 2918 | 1808 | Vascular endothelial growth factor A |
| PLAUR | 2404 | 3497 | 2581 | 1114 | 2399 | 981 | Urokinase plasminogen activator surface receptor |
| CSF3 | 1989 | 897 | 770 | 6605 | 2565 | 2748 | Granulocyte colony-stimulating factor |
| CCL8 | 1107 | 2046 | 1098 | 3351 | 1900 | 1064 | C-C motif chemokine 8 |
| CCL4 | 1851 | 1055 | 1312 | 2689 | 1727 | 722 | C-C motif chemokine 4 |
| CD40 | 1730 | 1681 | 841 | 1380 | 1408 | 408 | Tumor necrosis factor receptor superfamily member 5 |
| CD14 | 1467 | 2114 | 1175 | 555 | 1328 | 648 | Monocyte differentiation antigen CD14 |
| EGFR | 1006 | 1602 | 792 | 1938 | 1335 | 529 | Epidermal growth factor receptor |
| AXL | 958 | 2294 | 1183 | 277 | 1178 | 838 | Tyrosine-protein kinase receptor UFO |
| TREM1 | 961 | 1063 | 709 | 1549 | 1071 | 352 | Triggering receptor expressed on myeloid cells 1 |
| TYRO3 | 2334 | 123 | 1155 | 494 | 1026 | 971 | Tyrosine-protein kinase receptor TYRO3 |
| HGF | 823 | 832 | 1407 | 778 | 960 | 299 | Hepatocyte growth factor |
| LIF | 676 | 834 | 1014 | 1295 | 954 | 265 | Leukemia inhibitory factor |
| CXCL16 | 514 | 1256 | 539 | 993 | 825 | 362 | C-X-C motif chemokine 16 |
| CXCL6 | 1144 | 546 | 41 | 1123 | 714 | 527 | C-X-C motif chemokine 6 |
| NRCAM | 574 | 723 | 50 | 1424 | 693 | 567 | Neuronal cell adhesion molecule |
| CCL21 | 731 | 1164 | 384 | 289 | 642 | 397 | C-C motif chemokine 21 |
| TGFB1 | 769 | 793 | 630 | 363 | 639 | 197 | Human TGF-beta 1 |
| SPP1 | 189 | 434 | 635 | 917 | 544 | 308 | Osteopontin |
| TNFRSF10D | 637 | 103 | 330 | 1033 | 526 | 403 | Tumor necrosis factor receptor superfamily member 10D |
| FAS | 608 | 399 | 540 | 203 | 438 | 179 | Tumor necrosis factor receptor superfamily member 6 |
| FGF7 | 617 | 308 | 575 | 208 | 427 | 200 | Fibroblast growth factor 7 |
| SIGLEC5 | 456 | 545 | 288 | 446 | 434 | 107 | Sialic acid-binding Ig-like lectin 5 |
| IL23A | 441 | 425 | 166 | 655 | 422 | 200 | Interleukin-23 subunit alpha |
| IL17B | 532 | 453 | 438 | 156 | 395 | 164 | Interleukin-17B |
| MIF | 376 | 285 | 446 | 479 | 396 | 86 | Macrophage migration inhibitory factor |
| CCL1 | 88 | 112 | 73 | 1214 | 371 | 562 | C-C motif chemokine 1 |
| CD40LG | 80 | 268 | 220 | 588 | 289 | 215 | CD40 ligand |
| CCL13 | 166 | 155 | 297 | 490 | 277 | 156 | C-C motif chemokine 13 |
| VEGFC | 568 | 215 | 160 | 23 | 241 | 232 | Vascular endothelial growth factor C |
| CCL11 | 159 | 103 | 416 | 294 | 243 | 141 | Eotaxin |
| IL1B | 271 | 179 | 187 | 294 | 233 | 58 | Interleukin-1 beta |
| TNF | 81 | 123 | 151 | 590 | 236 | 237 | Tumor necrosis factor |
| ENG | 162 | 128 | 254 | 179 | 181 | 53 | Endoglin |
| TNFRSF21 | 62 | 199 | 76 | 192 | 132 | 73 | Tumor necrosis factor receptor superfamily member 21 |
| CCL7 | 58 | 72 | 54 | 362 | 137 | 150 | C-C motif chemokine 7 |
| CCL3 | 90 | 19 | 25 | 251 | 96 | 108 | C-C motif chemokine 3 |
| IL2RA | 108 | 74 | 72 | 58 | 78 | 21 | Interleukin-2 receptor subunit alpha |
| AGRP | 17 | 125 | 16 | 113 | 68 | 59 | Agouti-related protein |
| IL1A | 85 | 25 | 26 | 115 | 63 | 45 | Interleukin-1 alpha |
| CXCL12 | 71 | 92 | 19 | 28 | 52 | 35 | Stromal cell-derived factor 1 |
| IL17A | 87 | 1 | 34 | 78 | 50 | 40 | Interleukin-17A |
| PGF | 25 | 8 | 20 | 100 | 38 | 42 | Placenta growth factor |
| IL13 | 18 | 9 | 11 | 25 | 16 | 7 | Interleukin-13 |
| Factor Amount | GO TERM | Description | p-Value | Factors |
|---|---|---|---|---|
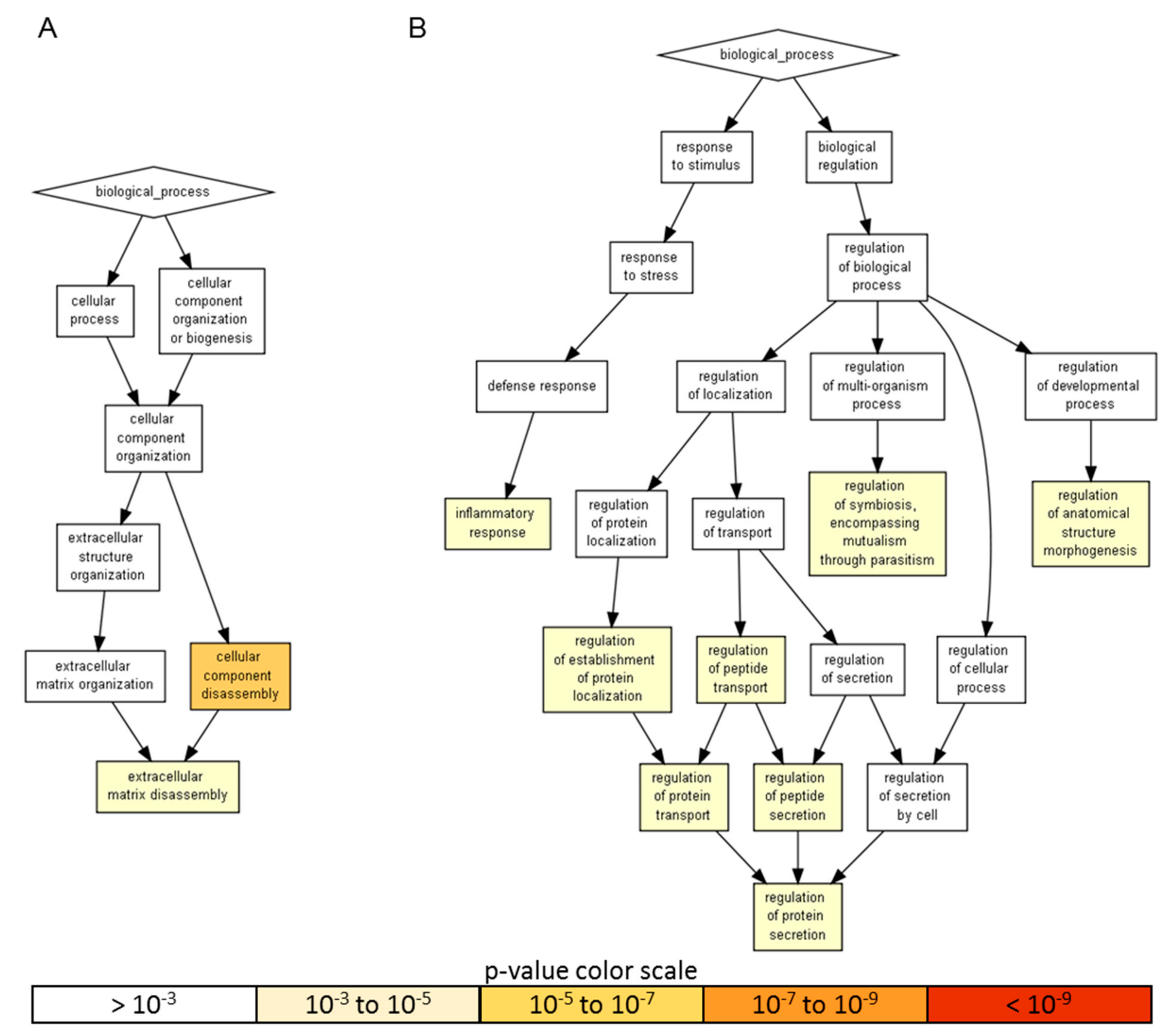
| > 10 ng/106 ASC | GO:0022411 | cellular component disassembly | 3.74 × 10−6 | TIMP1, TIMP2, DKK1, PLG and CTSS |
| GO:0022617 | extracellular matrix disassembly | 5.13 × 10−5 | TIMP1, TIMP2, PLG and CTSS | |
| < 10 ng/106 ASC | GO:0002791 | regulation of peptide secretion | 8.05 × 10−4 | IL17A, ANG, VEGFC, TGFB1, CCL5, IL13, IL1RL1, MIF, CD40LG, IL1B, CD40, CCL1, IL1A, CCL3, TNFRSF21, TNF, EGFR, CD14 and TNFRSF1B |
| GO:0050708 | regulation of protein secretion | 8.05 × 10−4 | IL17A, ANG, VEGFC, TGFB1, CCL5, IL13, IL1RL1, MIF, CD40LG, CCL1, CD40, IL1B, IL1A, CCL3, TNFRSF21, TNF, EGFR, CD14 and TNFRSF1B | |
| GO:0022603 | regulation of anatomical structure morphogenesis | 8.07 × 10−4 | SHH, FLT1, CXCL9, GDF15, LIF, PGF, CXCL12, SPP1, FGF7, ENG, CSF1, VEGFA, CCL13, VEGFC, CCL11, TGFB1, CCL8, CCL7, IL1B, CD40, IL1A, CCL3, HGF, TNF, IL8, TNFRSF1B, NRCAM and TNFSRSF11B | |
| GO:0043903 | regulation of symbiosis | 8.22 × 10−4 | TNF, IL8, CCL5, CCL4, CCL8, CXCL6 and CCL3 | |
| GO:0070201 | regulation of protein localization | 9.16 × 10-4 | IL17A, ANG, VEGFC, TGFB1, SHH, CCL5, IL13, IL1RL1, MIF, CD40LG, CCL1, IL1B, CD40, IL1A, CCL3, TNFRSF21, TNF, EGFR, CD14 and TNFRSF1B | |
| GO:0051223 | regulation of protein transport | 9.16 × 10−4 | IL17A, ANG, VEGFC, TGFB1, SHH, CCL5, IL13, IL1RL1, MIF, CD40LG, IL1B, CD40, CCL1, IL1A, CCL3, TNFRSF21, TNF, EGFR, CD14 and TNFRSF1B | |
| GO:0090087 | regulation of peptide transport | 9.16 × 10−4 | IL17A, ANG, VEGFC, TGFB1, SHH, CCL5, IL13, IL1RL1, MIF, CD40LG, CCL1, CD40, IL1B, IL1A, CCL3, TNFRSF21, TNF, EGFR, CD14 and TNFRSF1B | |
| GO:0006954 | inflammatory response | 9.98 × 10−4 | IL17A, IL17B, CXCL9, IL13, MIF, SPP1, IL23A, AXL, CD14, CSF1, CCL13, CCL11, TGFB1, CCL5, IL2RA, CCL4, CCL8, CCL7, CD40LG, TREM1, IL1B, CD40, CCL1, IL1A, CCL3, TNF, CXCL1, IL8, CXCL6, TNFRSF1B and CCL21 |
| Protective | ||||
|---|---|---|---|---|
| miRNA | % EV-genetic weight | Target | Tissue | Role |
| hsa-miR-24-3p | 19.0 | P16INK4A | Cartilage | Regulates senescence |
| hsa-miR-222-3p | 5.8 | HDAC4 | Cartilage | Prevents cartilage degradation |
| hsa-miR-193b-3p | 5.0 | TGFB2-3/MMP19 | Cartilage | Regulates inflammation |
| hsa-miR-92a-3p | 1.5 | ADAMTS4-5/HDAC2 | Cartilage | Anti-catabolic |
| hsa-miR-29a-3p | 1.1 | VEGF | Synovium | Inhibits synovial remodeling |
| hsa-miR-199a-3p | 0.9 | COX2 | Cartilage | Anti-catabolic |
| hsa-miR-27a-3p | 0.8 | FSTL1 | Synovium | Prevents synoviocyte migration |
| hsa-miR-26a-5p | 0.6 | iNOS | Cartilage | Cartilage homeostasis |
| hsa-miR-130a-3p | 0.6 | TNFa | Cartilage | Anti-inflammatory |
| hsa-miR-320a-3p | 0.5 | BMI1/RUNX2/MMP13 | Cartilage | Chondrogenic |
| hsa-miR-152-3p | 0.5 | DNMT | Synovium | Prevents synoviocyte migration |
| hsa-miR-17-5p | 0.5 | SQSTM1 | Cartilage | Induces Autophagy |
| hsa-miR-27b-3p | 0.3 | MMP13/LEPTIN | Cartilage | Anti-catabolic |
| hsa-miR-210-3p | 0.2 | DR6/HIF3a | Cartilage | Anti-apoptotic/promotes ECM |
| hsa-miR-30a-5p | 0.2 | ADAMTS5 | Cartilage | Cartilage homeostasis |
| hsa-miR-26b-5p | 0.1 | KPNA3 | Cartilage | Promotes NF-kB pathway |
| hsa-miR-149-5p | 0.1 | TNFa | Cartilage | Anti-inflammatory |
| Total | 37.9 | |||
| Destructive | ||||
| miRNA | % EV-genetic weight | Target | Tissue | Role |
| hsa-miR-21-5p | 4.7 | GDF5 | Cartilage | Chondrogenesis inhibitor |
| hsa-miR-30b-5p | 2.4 | BECN1/ATG5 | Cartilage | ECM degradation |
| hsa-miR-19b-3p | 1.4 | nd | Synovium | Induces NF-kB signaling |
| hsa-miR-34a-5p | 0.7 | SIRT1 | Cartilage | Apoptotic |
| hsa-miR-16-5p | 0.4 | SMAD3 | Cartilage | Cartilage degradation |
| hsa-miR-138-5p | 0.2 | FOXC1 | Cartilage | Promotes cartilage degradation |
| hsa-miR-483-5p | 0.1 | MATN3/TIMP2 | Cartilage | ECM degradation |
| Total | 9.8 | |||
| Protective/Destructive | ||||
| miRNA | % EV-genetic weight | Target | Tissue | Role |
| hsa-miR-125b-5p | 12.6 | ADAMTS4/SYVN1 | Cartilage/Synovium | Prevents aggrecan loss/Pro-apoptotic |
| hsa-miR-221-3p | 4.7 | SDF1/nd | Cartilage | Prevents ECM degradation/Pro-inflammatory |
| hsa-miR-145-5p | 1.7 | TNFRSF11B/SOX9-SMAD3 | Cartilage | Chondrocyte proliferation/Cartilage degradation |
| hsa-miR-365-3p | 0.2 | HIF2a/HDAC4 | Cartilage | Prevents ECM loss/Pro-inflammatory |
| Total | 19.2 | |||
| M1 Polarization | ||
|---|---|---|
| miRNAs | % EV-Genetic Weight | Regulation of Macrophage Phenotype |
| hsa-miR-145-5p | 1.7 | Promotes M1 |
| hsa-miR-27a-3p | 0.8 | Promotes M1, suppresses M2 |
| hsa-miR-27b-3p | 0.3 | Promotes M1, suppresses M2 |
| hsa-miR-130a-3p | 0.6 | Promotes M1, suppresses M2 |
| hsa-miR-26a-5p | 0.6 | Suppresses M2 |
| hsa-miR-26b-5p | 0.1 | Suppresses M2 |
| hsa-miR-720 | 3.0 | Suppresses M2 |
| Total | 7.2 | |
| M2 Polarization | ||
| miRNAs | % EV-Genetic Weight | Regulation of Macrophage Phenotype |
| hsa-miR-34a-5p | 0.7 | Promotes M2 |
| hsa-miR-222-3p | 5.8 | Promotes M2 |
| hsa-let-7b-5p | 0.6 | Promotes M2 |
| hsa-miR-24-3p | 19.0 | Promotes M2, suppresses M1 |
| Total | 26.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragni, E.; Perucca Orfei, C.; De Luca, P.; Colombini, A.; Viganò, M.; de Girolamo, L. Secreted Factors and EV-miRNAs Orchestrate the Healing Capacity of Adipose Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis. Int. J. Mol. Sci. 2020, 21, 1582. https://doi.org/10.3390/ijms21051582
Ragni E, Perucca Orfei C, De Luca P, Colombini A, Viganò M, de Girolamo L. Secreted Factors and EV-miRNAs Orchestrate the Healing Capacity of Adipose Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis. International Journal of Molecular Sciences. 2020; 21(5):1582. https://doi.org/10.3390/ijms21051582
Chicago/Turabian StyleRagni, Enrico, Carlotta Perucca Orfei, Paola De Luca, Alessandra Colombini, Marco Viganò, and Laura de Girolamo. 2020. "Secreted Factors and EV-miRNAs Orchestrate the Healing Capacity of Adipose Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis" International Journal of Molecular Sciences 21, no. 5: 1582. https://doi.org/10.3390/ijms21051582
APA StyleRagni, E., Perucca Orfei, C., De Luca, P., Colombini, A., Viganò, M., & de Girolamo, L. (2020). Secreted Factors and EV-miRNAs Orchestrate the Healing Capacity of Adipose Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis. International Journal of Molecular Sciences, 21(5), 1582. https://doi.org/10.3390/ijms21051582