Nano-Zn Increased Zn Accumulation and Triglyceride Content by Up-Regulating Lipogenesis in Freshwater Teleost, Yellow Catfish Pelteobagrus fulvidraco



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Growth Performance, Feed Utilization, and Morphological Parameters
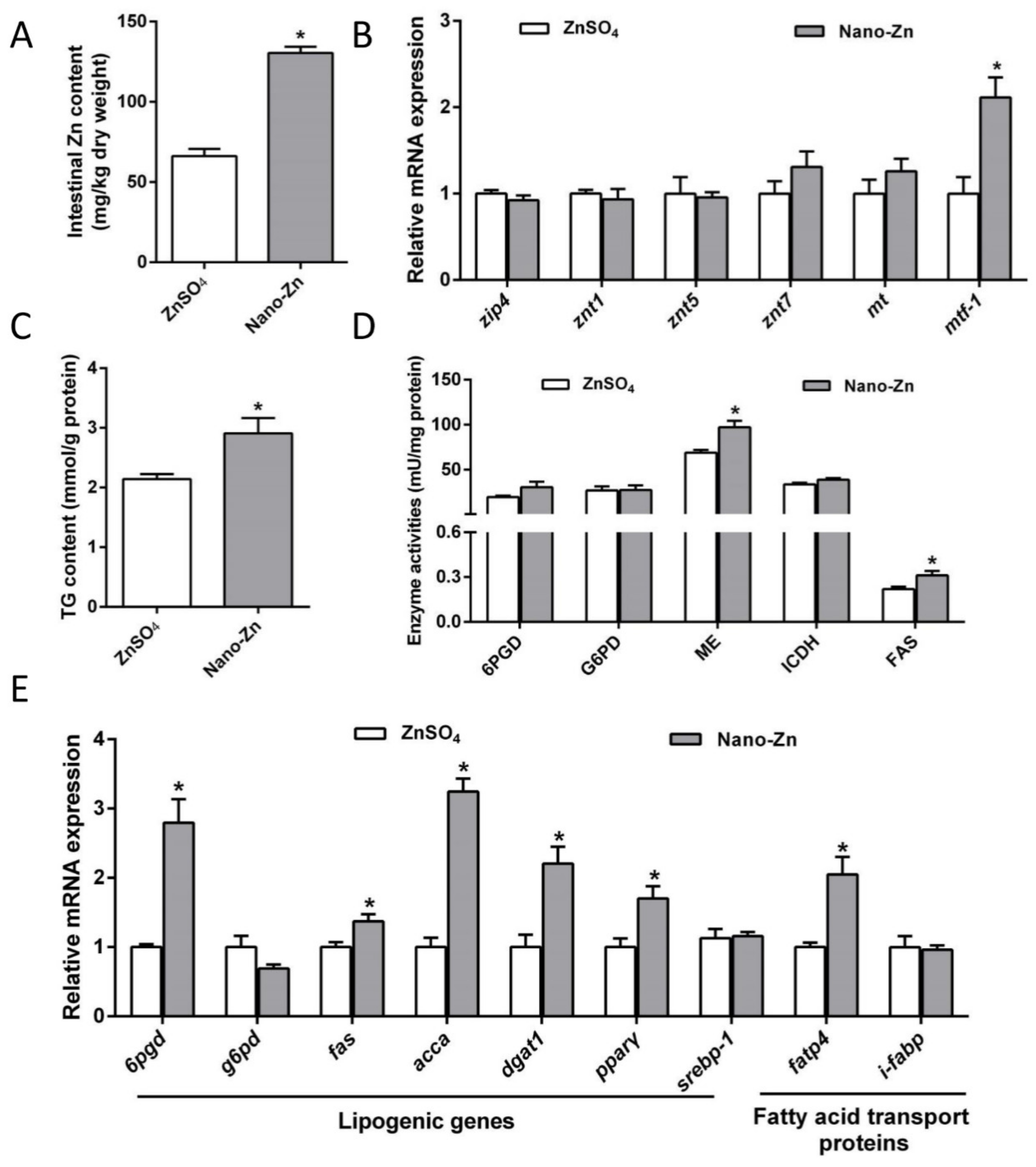
2.2. Zn Accumulation, Zn Absorption, and Lipid Metabolism in the Intestine
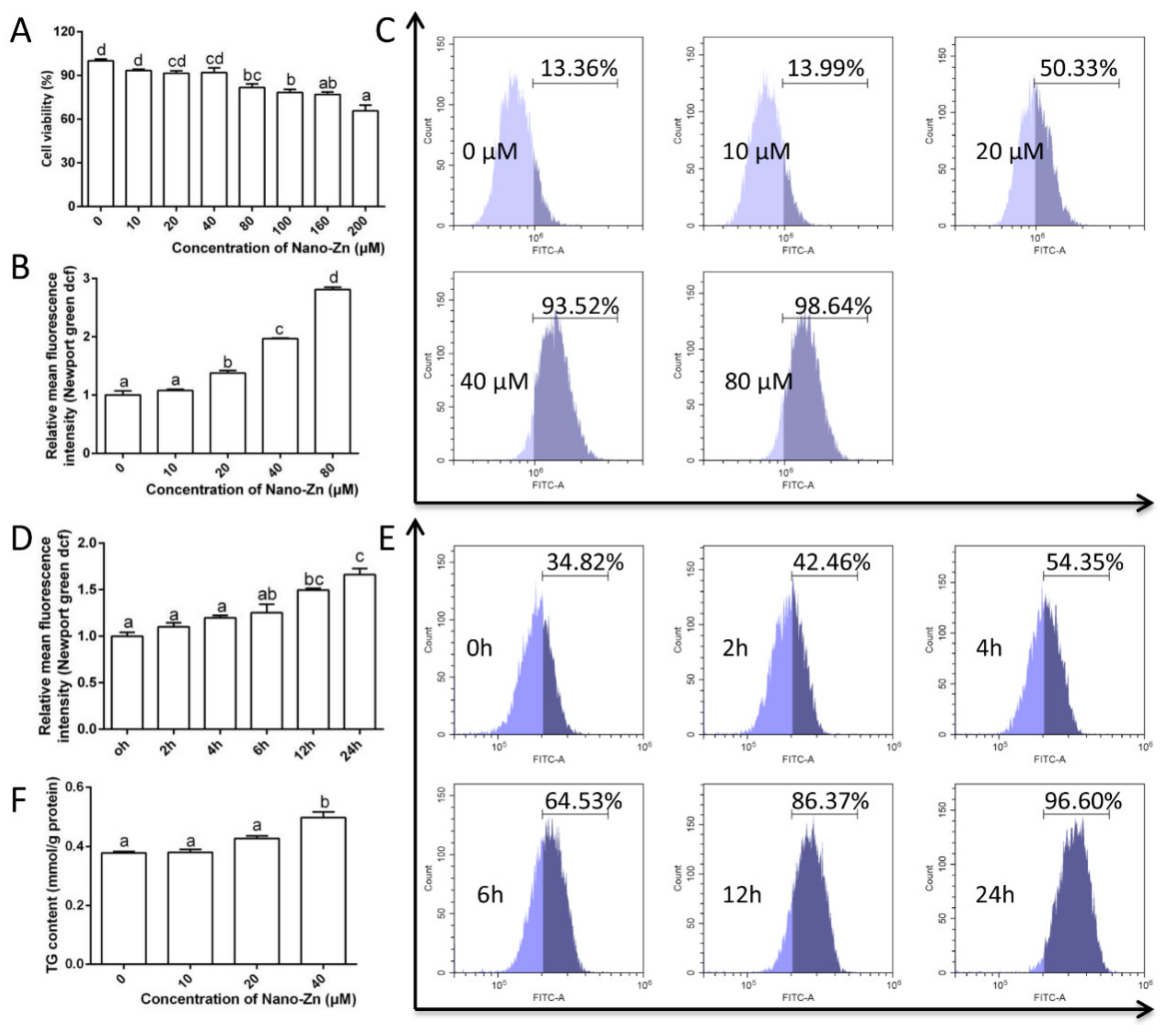
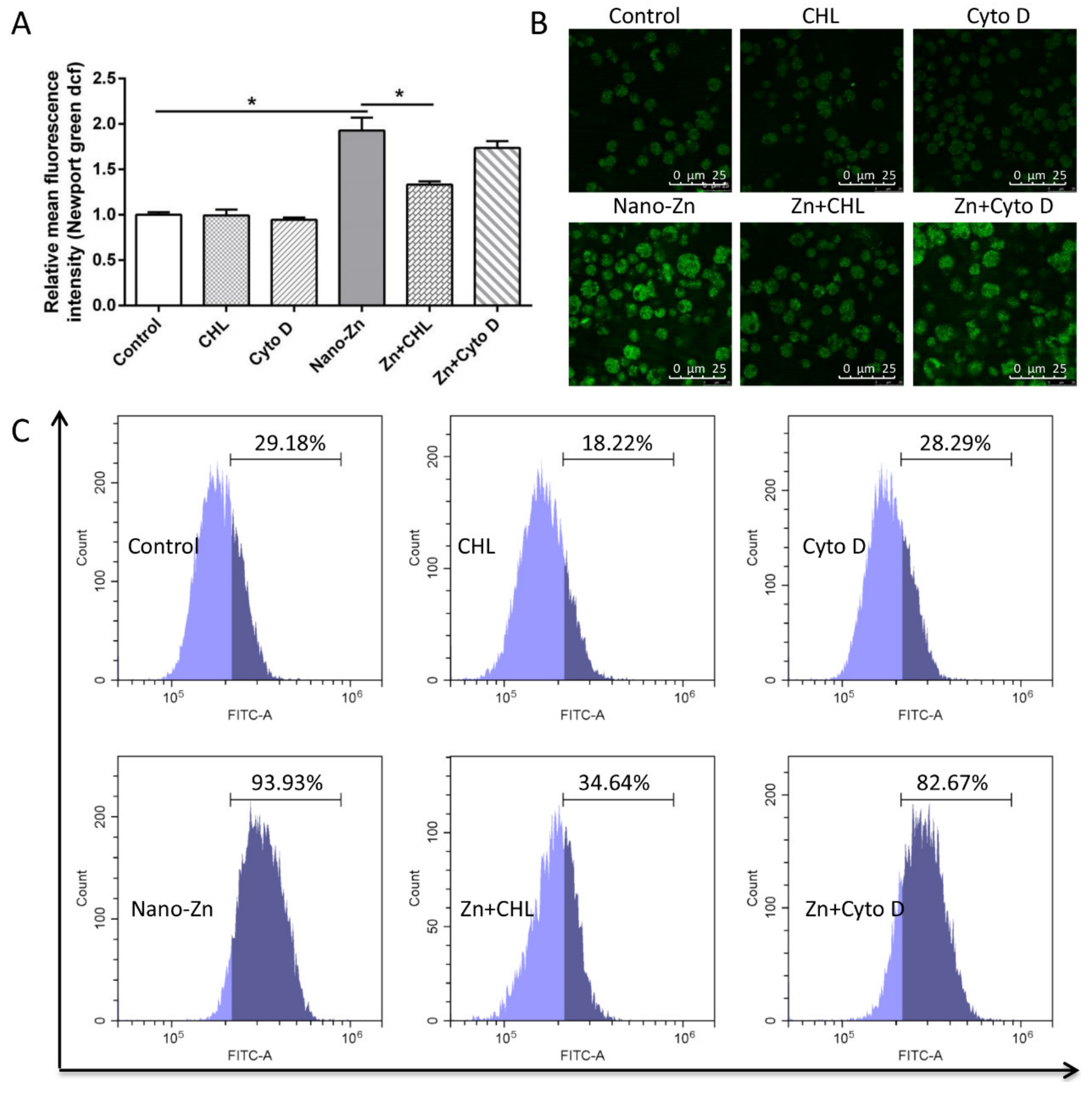
2.3. Intestinal Epithelial Cells Absorb Nano-Zn via Clathrin Pathway
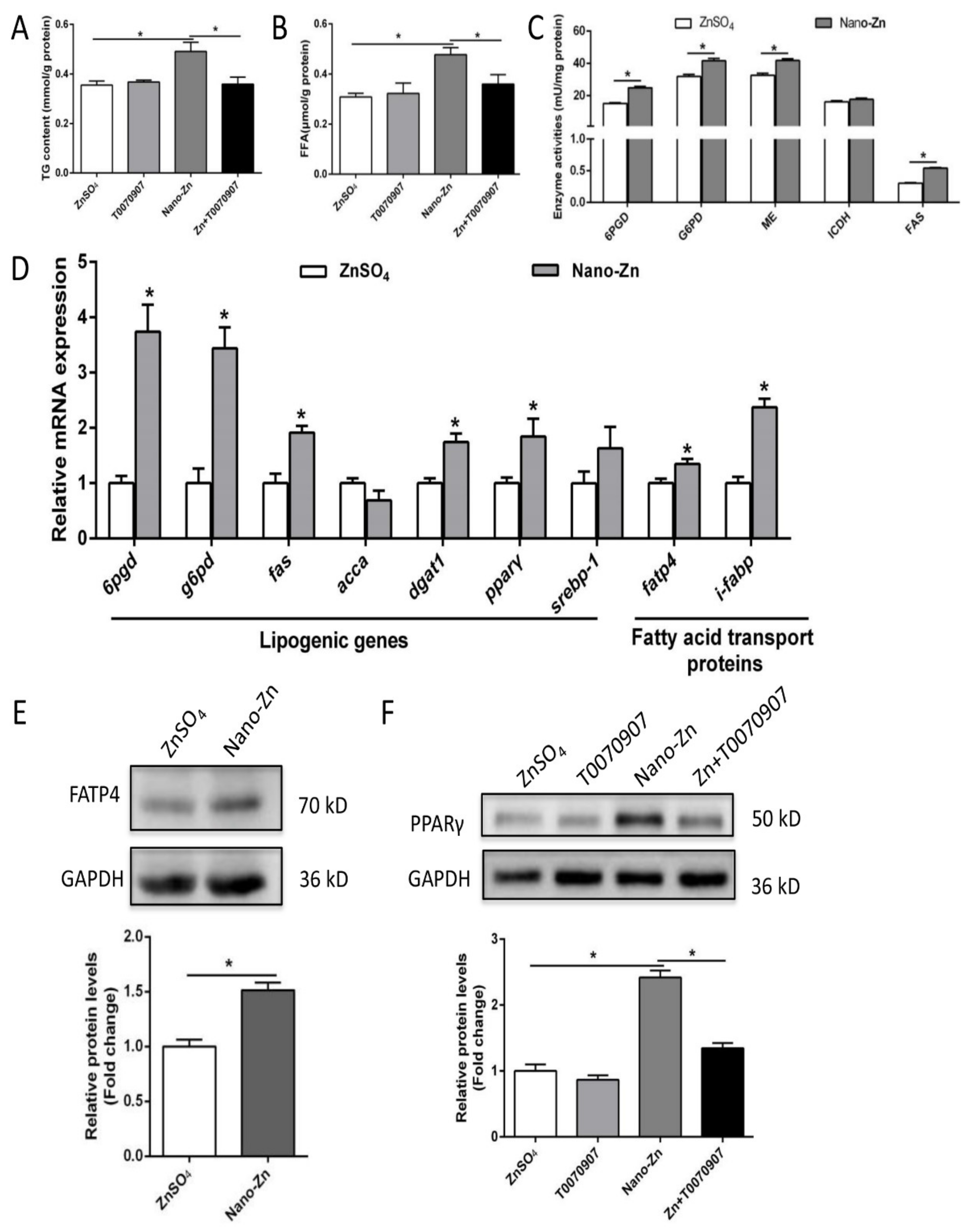
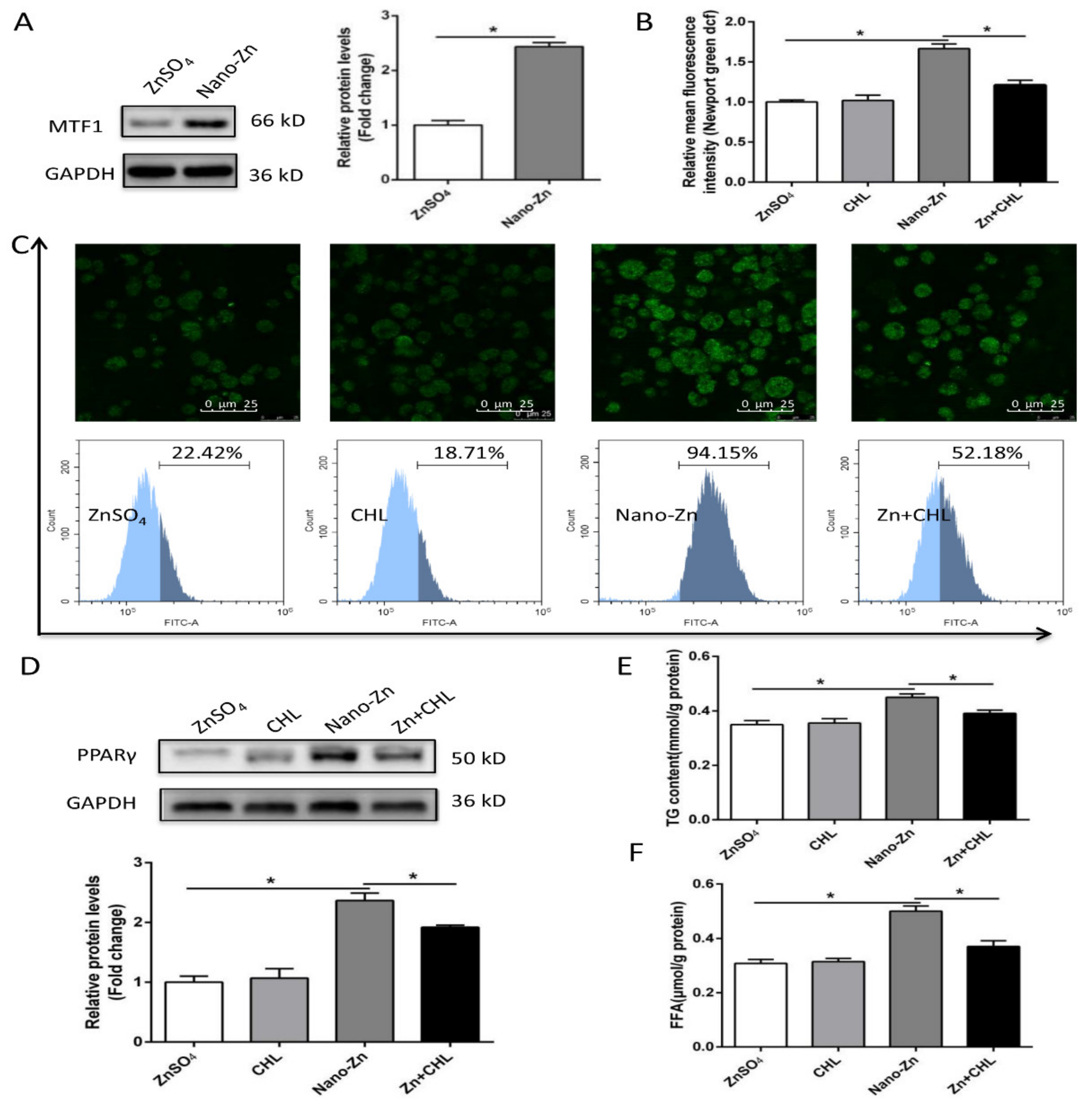
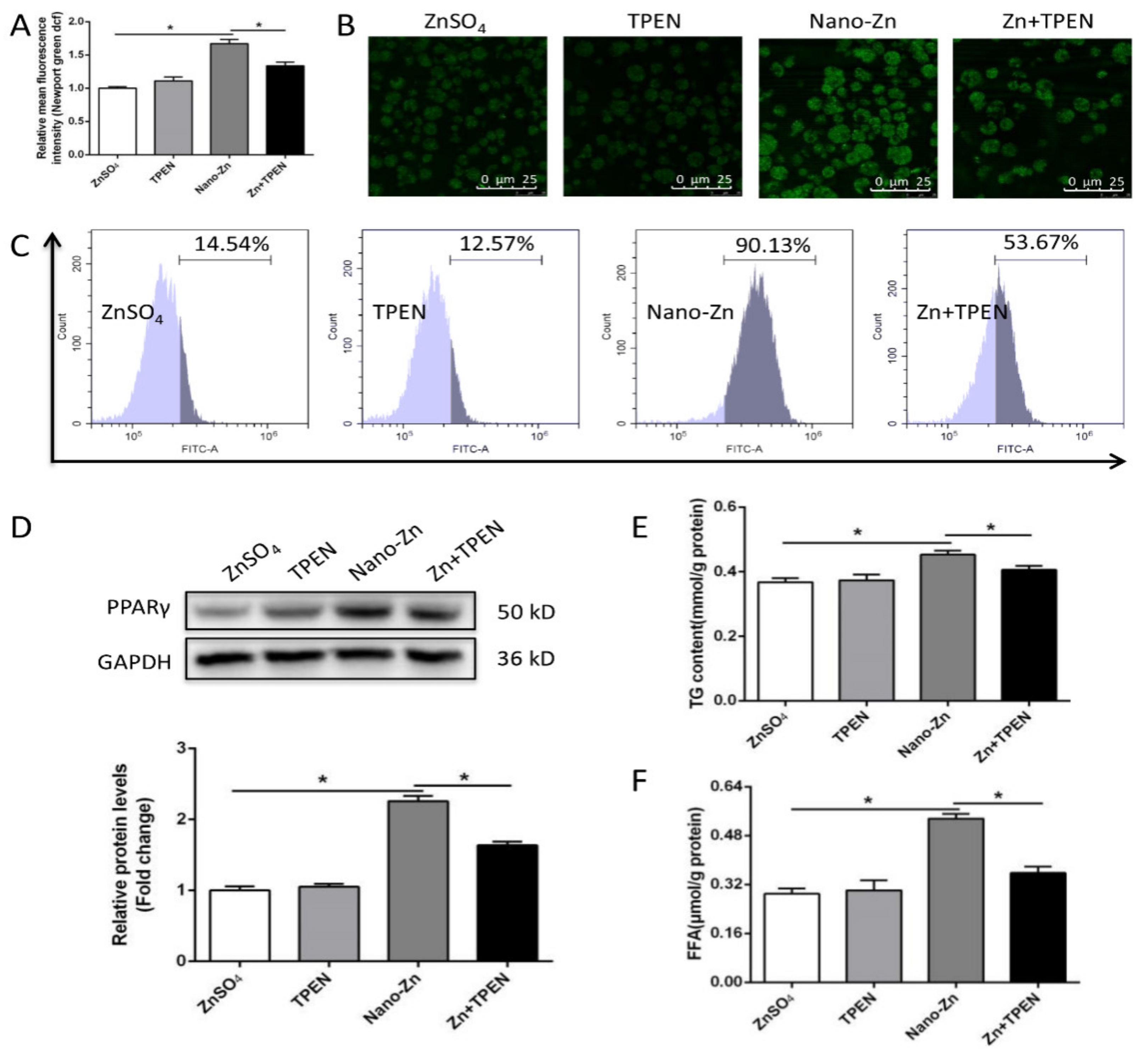
2.4. Higher TG Accumulation in Nano-Zn Group Than in Znso4 Group was Attributable to the Nano-Zn-Induced Activation of PPARγ
3. Discussion
4. Materials and Methods
4.1. Expt. 1: In Vivo Study
4.2. Expt. 2: In Vitro Study
4.3. Cell Viability, TG Content, and Enzymatic Activity Assays
4.4. Real-Time Quantitative PCR (qPCR)
4.5. Immunoblotting Analysis
4.6. Zn2+ Measurement
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 6PGD | 6-phosphogluconate dehydrogenase |
| ACC | acetyl-CoA carboxylase |
| ANOVA | one-way analysis of variance |
| CF | condition factor |
| DGAT | diacylglycerol acyltransferase |
| FAS | fatty acid synthase |
| FATP4 | fatty acid transport protein 4 |
| FCR | feed conversion rate |
| FFA | free fatty acid |
| FI | feed intake |
| G6PD | glucose 6-phosphate dehydrogenase |
| ICDH | isocitrate dehydrogenase |
| I-FABP | intestine fatty acid binding protein |
| ISI | intestinal somatic index |
| ME | malic enzyme |
| MS-222 | tricaine methanesulfonate |
| MT | metallothionein |
| MTF-1 | metal response element-binding transcription factor-1 |
| NaN3 | sodium azide |
| PPAR | peroxisome proliferator activated receptor |
| SEM | standard error of mean |
| SGR | specific growth rate |
| SREBP | sterol regulatory element-binding protein |
| TG | triglyceride |
| TPEN | N,N,N′,N′-tetrakis (2-pyridylmethyl) ethylenediamine |
| VSI | viscerosomatic index |
| WG | weight gain |
| ZIP | ZRT, IRT-like protein |
| ZnT | zinc transporter |
| Zn | zinc |
| ZnSO4 | zinc sulfate |
| Nano-Zn | zinc nanoparticles |
References
- Watanabe, T.; Kiron, V.; Satoh, S.; Wilson, R.P.; Wee, K.L. Trace minerals in fish nutrition. Aquaculture 1997, 151, 185–207. [Google Scholar] [CrossRef]
- Bury, N.R.; Walker, P.A.; Glover, C.N. Nutritive metal uptake in teleost fish. J. Exp. Biol. 2003, 206, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.H.; Luo, Z.; Wei, C.C.; Li, D.D.; Pan, Y.X. Six indicator genes for zinc (Zn) homeostasis in freshwater teleost yellow catfish Pelteobagrus fulvidraco: molecular characterization, mRNA tissue expression and transcriptional changes to Zn exposure. Biometals 2018, 31, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Findley, S.D. Cloning and functional characterization of a mammalian zinc transporter that confers resistance to zinc. EMBO J. 1995, 14, 639–649. [Google Scholar] [CrossRef]
- Ishihara, K.; Yamazaki, T.; Ishida, Y.; Suzuki, T.; Oda, K.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Zinc transport complexes contribute to the homeostasis maintenance of secretory pathway function in vertebrate cells. J. Biol. Chem. 2006, 281, 17743–17750. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Tan, X.Y.; Zheng, J.L.; Chen, Q.L.; Liu, C.X. Quantitative dietary zinc requirement of juvenile yellow catfish Pelteobagrus fulvidraco, and effects on hepatic intermediary metabolism and antioxidant responses. Aquac. 2011, 319, 150–155. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc status is associated with inflammation, oxidative stress, lipid and glucose metabolism. J. Physiol. Sci. 2018, 68, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.L.; Luo, Z.; Hu, W.; Liu, C.X.; Chen, Q.L.; Zhu, Q.L.; Gong, Y. Different effects of dietary Zn deficiency and excess on lipid metabolism in yellow catfish Pelteobagrus fulvidraco. Aquaculture 2015, 435, 10–17. [Google Scholar] [CrossRef]
- Chithrani, B.D.; Chan, W.C.W. Elucidating the mechanism of cellular uptake and removal of protein-coated gold nanoparticles of different sizes and shapes. Nano Lett. 2007, 7, 1542–1550. [Google Scholar] [CrossRef]
- Zhao, C.Y.; Tan, S.X.; Xiao, X.Y.; Qiu, X.S.; Pan, J.Q.; Tang, Z.X. Effects of dietary zinc oxide nanoparticles on growth performance and antioxidant status in broilers. Biol. Trace Elem. Res. 2014, 160, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Faiz, H.; Zuberi, A.; Nazir, S.; Rauf, M.; Younus, N. Zinc oxide, zinc sulfate and zinc oxide nanoparticles as source of dietary zinc: comparative effects on growth and hematological indices of juvenile grass carp (Ctenopharyngodon idela). Int. J. Agric. Biol. 2015, 17, 568–574. [Google Scholar] [CrossRef]
- Award, A.; Zaglool, A.W.; Ahmed, S.A.A.; Khalil, S.R. Transcriptomic profile change, immunological response and disease resistance of Oreochromis niloticus fed with conventional and Nano-Zinc oxide dietary supplements. Fish Shellfish Immunol. 2019, 93, 336–343. [Google Scholar]
- Oh, J.M.; Choi, S.J.; Kim, S.T.; Choy, J.H. Cellular uptake mechanism of an inorganic nanovehicle and its drug conjugates: enhanced efficacy due to clathrin-mediated endocytosis. Bioconjugate Chem. 2006, 17, 1411–1417. [Google Scholar] [CrossRef]
- Conner, S.D.; Schmid, S.L. Regulated portals of entry into the cell. Nature 2003, 422, 37–44. [Google Scholar] [CrossRef]
- Lina, T.; Jianyang, J.; Fenghua, Z.; Huiying, R.; Wenli, L. Effect of nano-zinc oxide on the production and dressing performance of broiler. Chin. Agricult. Sci. Bull. 2009, 25, 1–5, (in Chinese with English abstract). [Google Scholar]
- Mishra, A.; Swain, R.K.; Mishra, S.K.; Panda, N.; Sethy, K. Growth performance and serum biochemical parameters as affected by nano zinc supplementation in layer chicks. Ind J. Anim. Nutr. 2014, 31, 384–388. [Google Scholar]
- Horký, P.; Skalickova, S.; Urbankova, L.; Baholet, D.; Kociova, S.; Richtera, L.; Kabourkova, E.; Lacková, Z.; Cernei, N.; Gagic, M.; et al. Zinc phosphate-based nanoparticles as a novel antibacterial agent: in vivo study on rats after dietary exposure. J. Anim. Sci. Biotechnol. 2019, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.H.; Hogstrand, C.; Luo, Z.; Zhang, D.G.; Ling, S.C.; Wu, K. Dietary zinc addition influenced zinc and lipid deposition in the fore-and mid-intestine of juvenile yellow catfish Pelteobagrus fulvidraco. Br. J. Nutr. 2017, 118, 570–579. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, A.; Wang, W.X. Evaluation of nano-ZnOs as a novel Zn source for marine fish: importance of digestive physiology. Nanotoxicology 2017, 11, 1026–1039. [Google Scholar] [CrossRef]
- Matito, E.; Putz, M.V. New link between conceptual density functional theory and electron delocalization. J. Phys. Chem. A 2011, 115, 12459–12462. [Google Scholar] [CrossRef] [PubMed]
- Putz, M.V.; Chattaraj, P.K. Electrophilicity kernel and its hierarchy through softness in concenptual density functional theory. Int. J. Quant. Chem. 2013, 113, 2163–2171. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Blanchard, R.K.; Cousins, R.J. Zinc supplementation of young men alters metallothionein, zinc transporter, and cytokine gene expression in leukocyte populations. Proc. Natl. Acad. Sci. USA 2006, 103, 1699–1704. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Qin, H.; Guo, J. Cooperation of metallothionein and zinc transporters for regulating zinc homeostasis in human intestinal Caco-2 cells. Nutr. Res. 2008, 28, 406–413. [Google Scholar] [CrossRef]
- Kirschke, C.P.; Huang, L. ZnT7, a novel mammalian zinc transporter, accumulates zinc in the Golgi apparatus. J. Biol. Chem. 2003, 278, 4096–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, S.K.; Czuba, E.; Selby, L.I.; Such, G.K.; Johnston, A.P.R. Quantifying nanoparticle internalization using a high throughput internalization assay. Pharm. Res. 2016, 33, 2421–2432. [Google Scholar] [CrossRef]
- Yumoto, R.; Nishikawa, H.; Okamoto, M.; Katayama, H.; Nagai, J.; Takano, M. Clathrin-mediated endocytosis of FITC-albumin in alveolar type II epithelial cell line RLE-6TN. Am. J. Physiol. 2006, 290, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Ranasinghe, P.; Wathurapatha, W.S.; Ishara, M.H.; Jayawardana, R.; Galappatthy, P.; Katulanda, P.; Constantine, G.R. Effects of zinc supplementation on serum lipids: a systematic review and meta-analysis. Nutr. Metab. (Lond.) 2015, 12, 26. [Google Scholar] [CrossRef]
- Wei, C.C.; Luo, Z.; Hogstrand, C.; Xu, Y.H.; Wu, L.X.; Chen, G.H.; Pan, Y.X.; Song, Y.F. Zinc reduces hepatic lipid deposition and activates lipophagy via Zn2+/MTF-1/PPARα and Ca2+/CaMKKβ/AMPK pathways. FASEB J. 2018, 32, 6666–6680. [Google Scholar] [CrossRef]
- Song, Y.F.; Hogstrand, C.; Wei, C.C.; Wu, K.; Pan, Y.X.; Luo, Z. Endoplasmic reticulum (ER) stress and cAMP/PKA pathway mediated Zn-induced hepatic lipolysis. Environ. Pollut. 2017, 228, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell. 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Meng, H.W.; You, H.M.; Yang, Y.; Zhang, Y.L.; Meng, X.M.; Ma, T.T.; Huang, C.; Li, J. 4-methylcoumarin-[5,6-g]-hesperetin attenuates inflammatory responses in alcoholic hepatitis through PPAR-γ activation. Toxicol. 2019, 421, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, D.S.; Kang, S. Vitamin D deficiency impairs glucose-stimulated insulin secretion and increases insulin resistance by reducing PPAR-γ expression in nonobese type 2 diabetic rats. J. Nutr. Biochem. 2016, 27, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, X.; Bian, F.; Wu, P.; Xing, S.; Xu, G.; Li, W.; Chi, J.; Ouyang, C.; Zheng, T.; et al. TNF-a promotes early atherosclerosis by increasing transcytosis of LDL across endothelial cells: crosstalk between NF-kB and PPAR-γ. J. Mol. Cell Cardiol. 2014, 72, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPAR[gamma] 2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef]
- Zheng, J.L.; Luo, Z.; Liu, C.X.; Chen, Q.L.; Zhu, Q.L.; Gong, Y. Differential effects of acute and chronic zinc (Zn) exposure on hepatic lipid deposition and metabolism in yellow catfish Pelteobagrus fulvidraco. Aquat. Toxicol. 2013, 132–133, 173–181. [Google Scholar] [CrossRef]
- Wu, K.; Luo, Z.; Hogstrand, C.; Chen, G.H.; Wei, C.C.; Li, D.D. Zn stimulates the phospholipids biosynthesis via the pathways of oxidative and endoplasmic reticulum stress in the intestine of freshwater teleost yellow catfish. Environ. Sci. Technol. 2018, 52, 9206–9214. [Google Scholar] [CrossRef] [Green Version]
- Khurana, R.K.; Kumar, R.; Gaspar, B.L.; Welsby, G.; Welsby, P.; Kesharwani, P.; Katare, O.P.; Singh, K.K.; Singh, B. Clathrin-mediated endocytic uptake of PUFA enriched self-nanoemulsifying lipidic systems (SNELS) of an anticancer drug against triple negative cancer and DMBA induced preclinical tumor model. Materials Sci. Engin. C 2018, 91, 645–658. [Google Scholar] [CrossRef]
- Dutta, D.; Donaldson, J.G. Search for inhibitors of endocytosis: Intended specificity and unintended consequences. Cell. Logist. 2012, 2, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by genometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, S.-C.; Zhuo, M.-Q.; Zhang, D.-G.; Cui, H.-Y.; Luo, Z. Nano-Zn Increased Zn Accumulation and Triglyceride Content by Up-Regulating Lipogenesis in Freshwater Teleost, Yellow Catfish Pelteobagrus fulvidraco. Int. J. Mol. Sci. 2020, 21, 1615. https://doi.org/10.3390/ijms21051615
Ling S-C, Zhuo M-Q, Zhang D-G, Cui H-Y, Luo Z. Nano-Zn Increased Zn Accumulation and Triglyceride Content by Up-Regulating Lipogenesis in Freshwater Teleost, Yellow Catfish Pelteobagrus fulvidraco. International Journal of Molecular Sciences. 2020; 21(5):1615. https://doi.org/10.3390/ijms21051615
Chicago/Turabian StyleLing, Shi-Cheng, Mei-Qin Zhuo, Dian-Guang Zhang, Heng-Yang Cui, and Zhi Luo. 2020. "Nano-Zn Increased Zn Accumulation and Triglyceride Content by Up-Regulating Lipogenesis in Freshwater Teleost, Yellow Catfish Pelteobagrus fulvidraco" International Journal of Molecular Sciences 21, no. 5: 1615. https://doi.org/10.3390/ijms21051615
APA StyleLing, S. -C., Zhuo, M. -Q., Zhang, D. -G., Cui, H. -Y., & Luo, Z. (2020). Nano-Zn Increased Zn Accumulation and Triglyceride Content by Up-Regulating Lipogenesis in Freshwater Teleost, Yellow Catfish Pelteobagrus fulvidraco. International Journal of Molecular Sciences, 21(5), 1615. https://doi.org/10.3390/ijms21051615