Transcriptomics and Proteomics Reveal the Cellulose and Pectin Metabolic Processes in the Tension Wood (Non-G-Layer) of Catalpa bungei

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
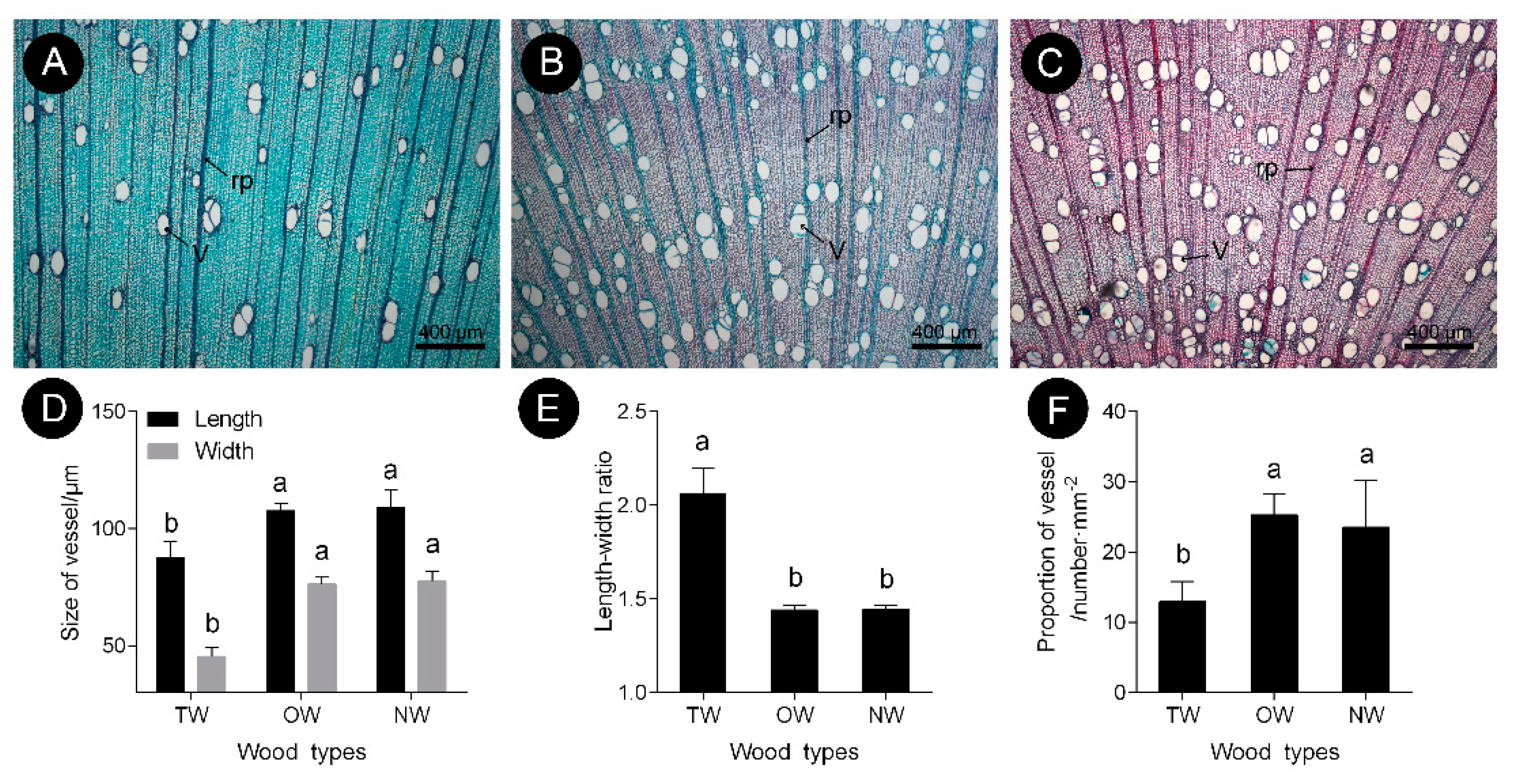
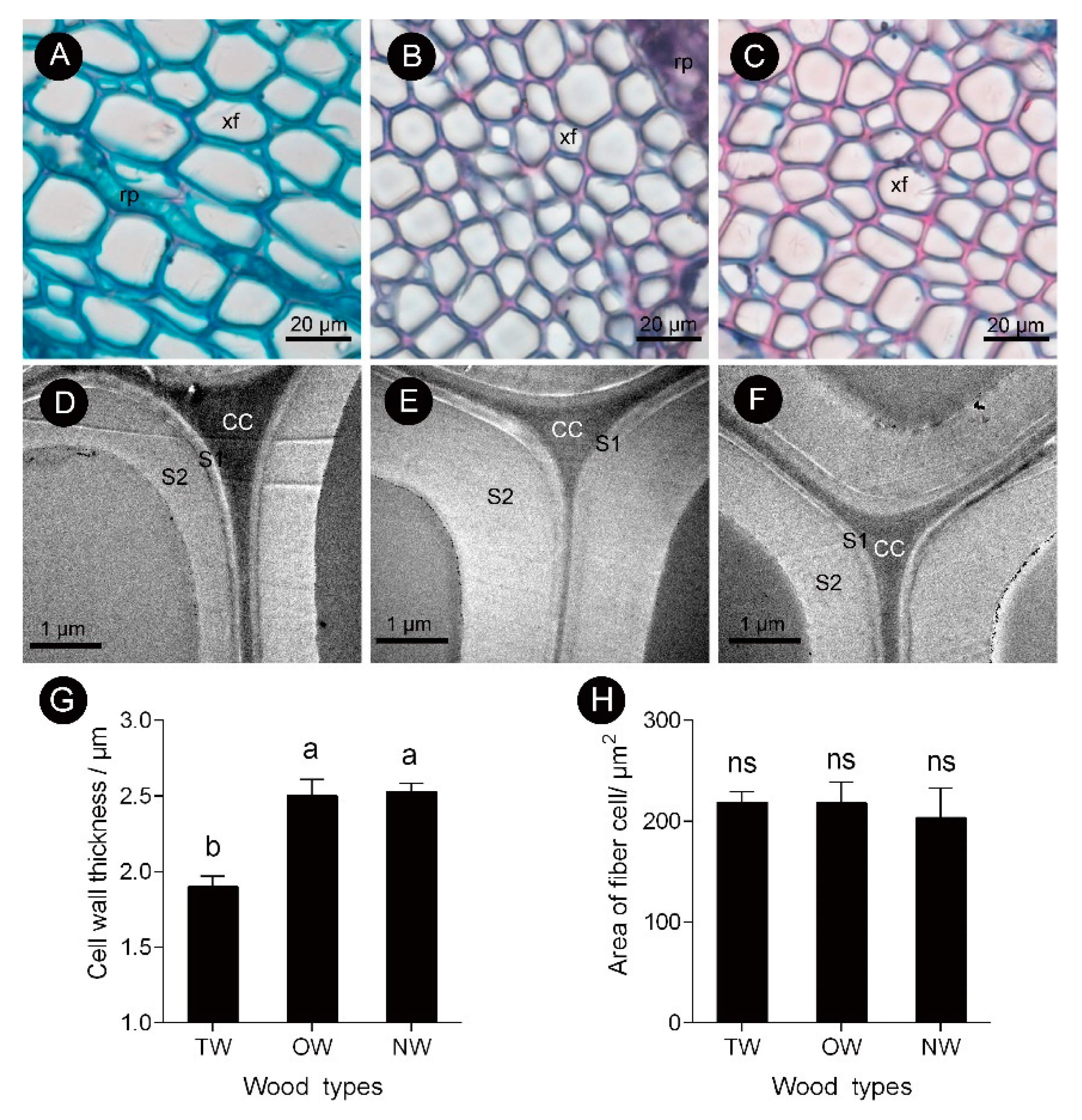
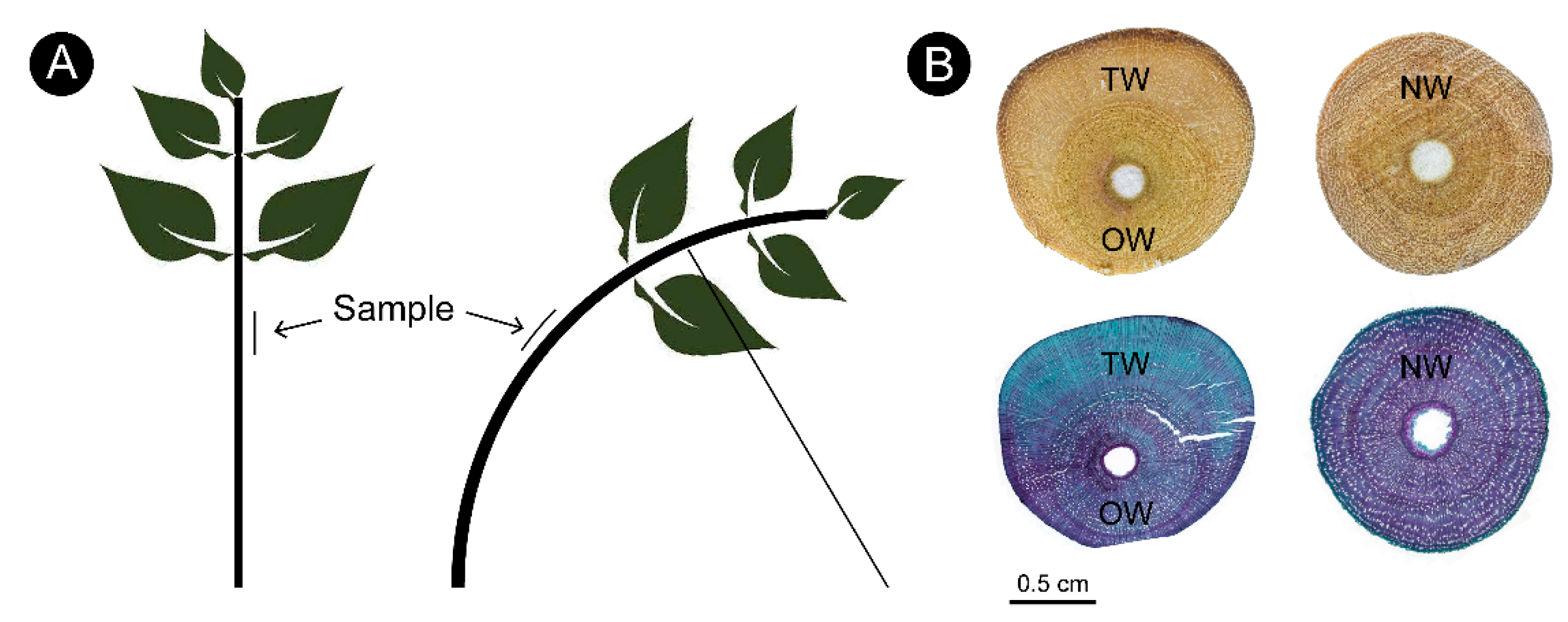
2.1. Anatomical Morphology of Different Types of C. bungei Wood
2.2. Identification and Characterization of mRNA and Protein In Different Types of Wood in C. bungei
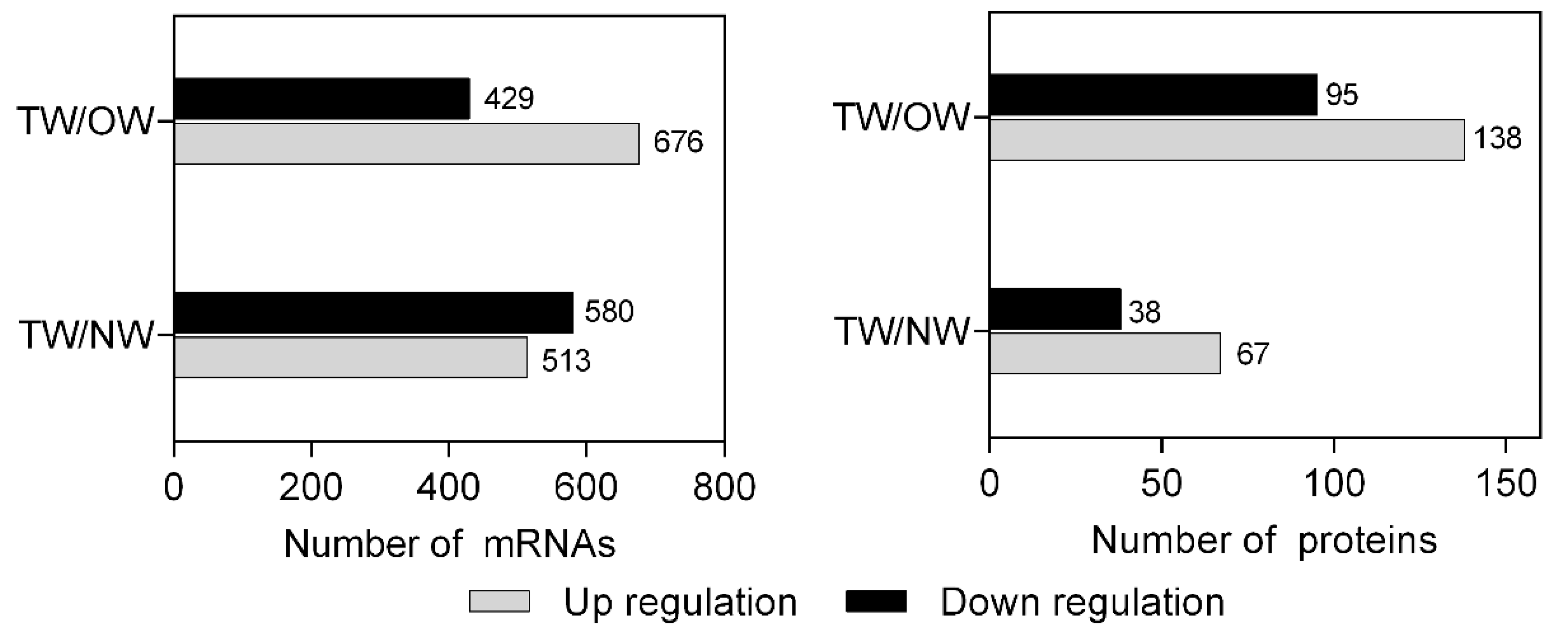
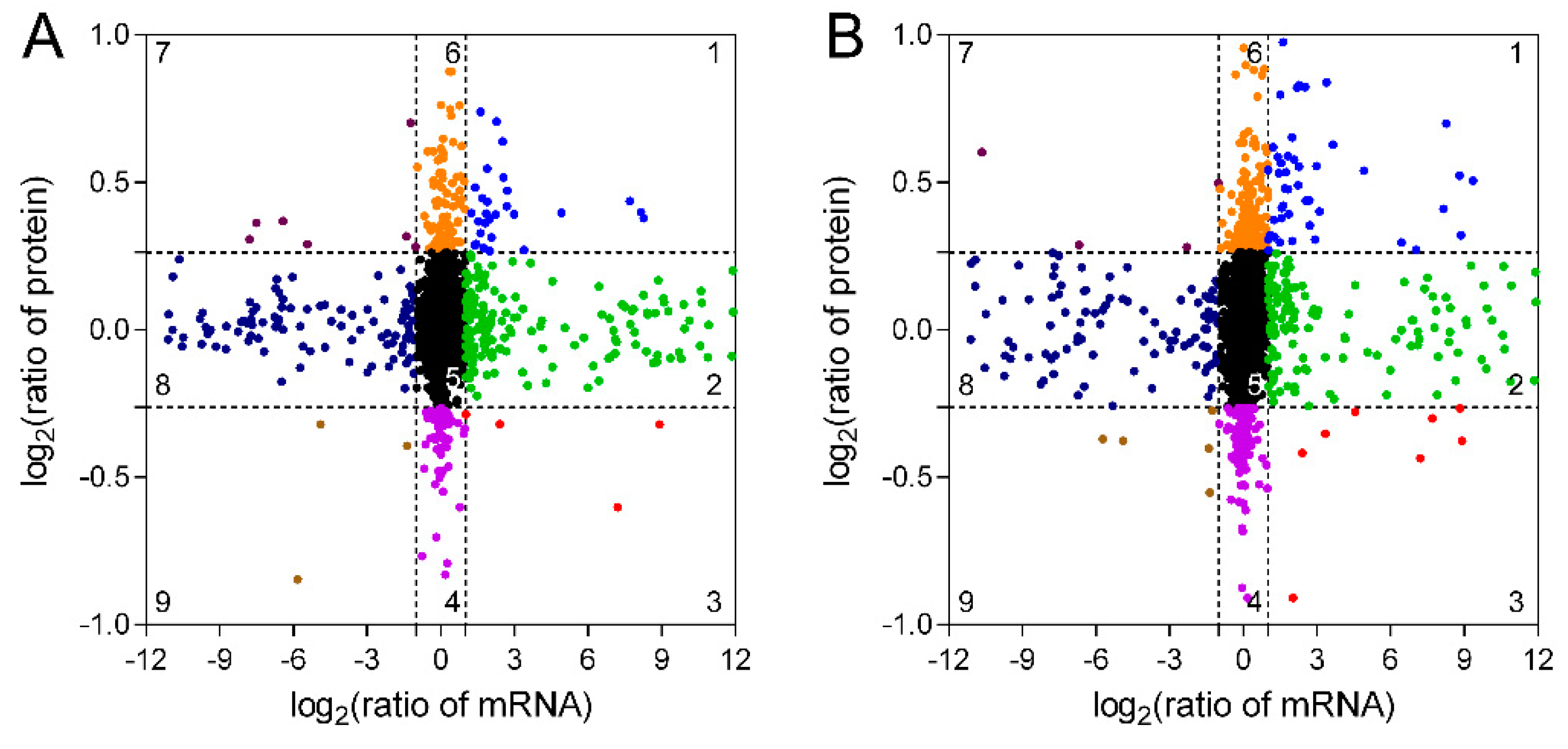
2.3. DEGs and DEPs Between Different Wood Types
2.4. GO Analyses of DEGs and DEPs in Different Wood Types
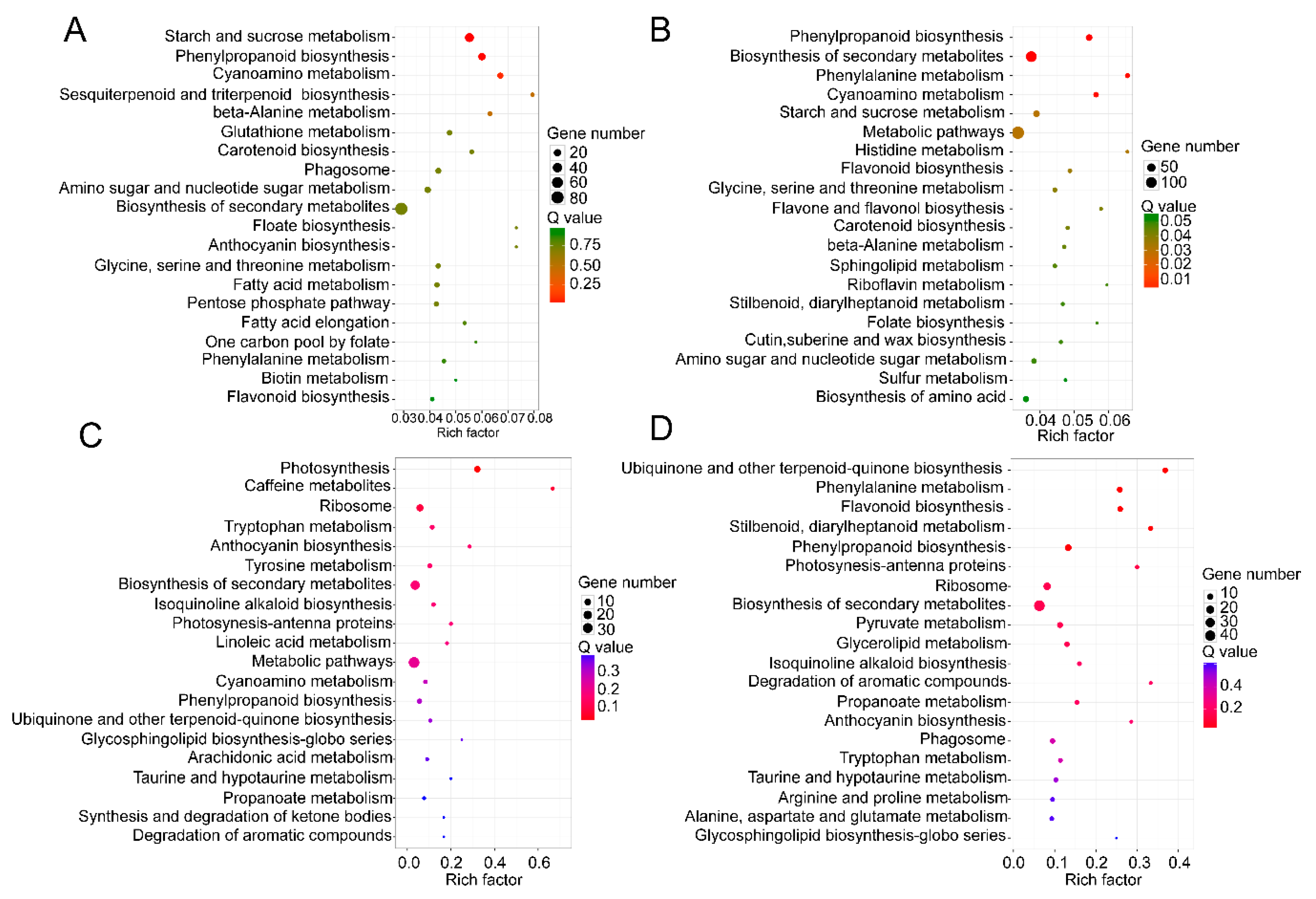
2.5. KEGG Pathway Analyses of DEGs and DEPs in Different Wood Types
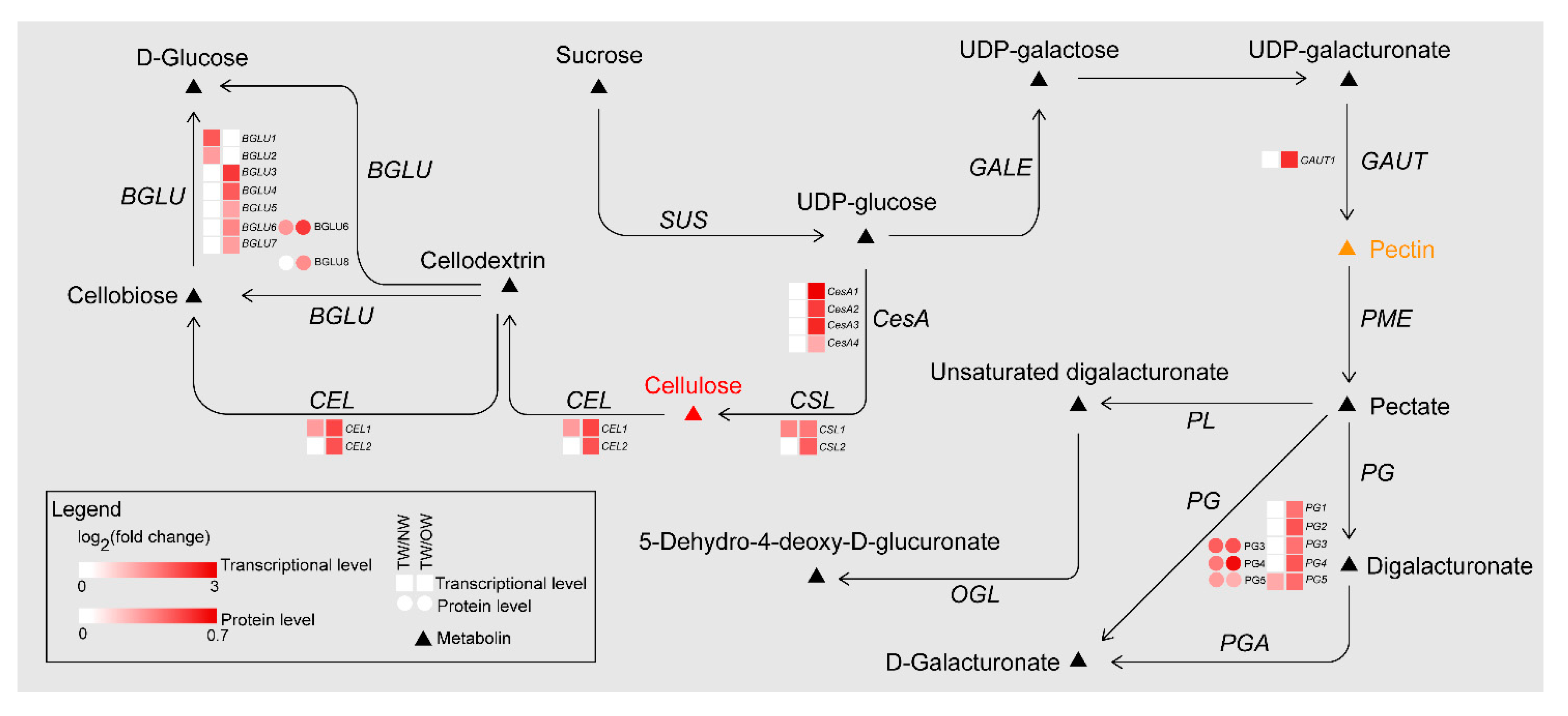
2.6. Changes in Gene and Protein Expression in Cellulose, Hemicellulose and Pectin Biosynthesis during TW Formation
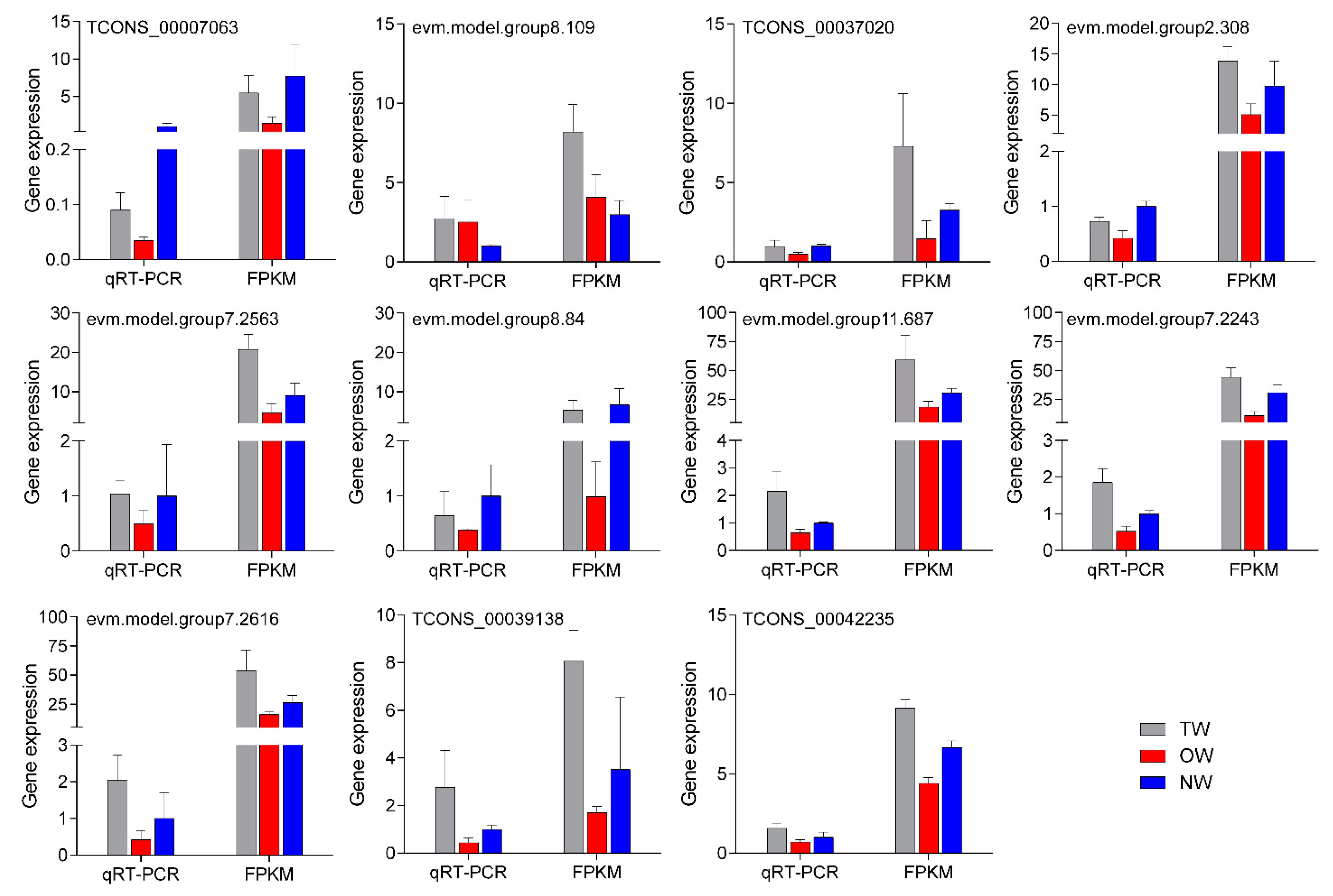
2.7. Transcript Levels and qRT-PCR Validation of mRNA in Different Wood Types
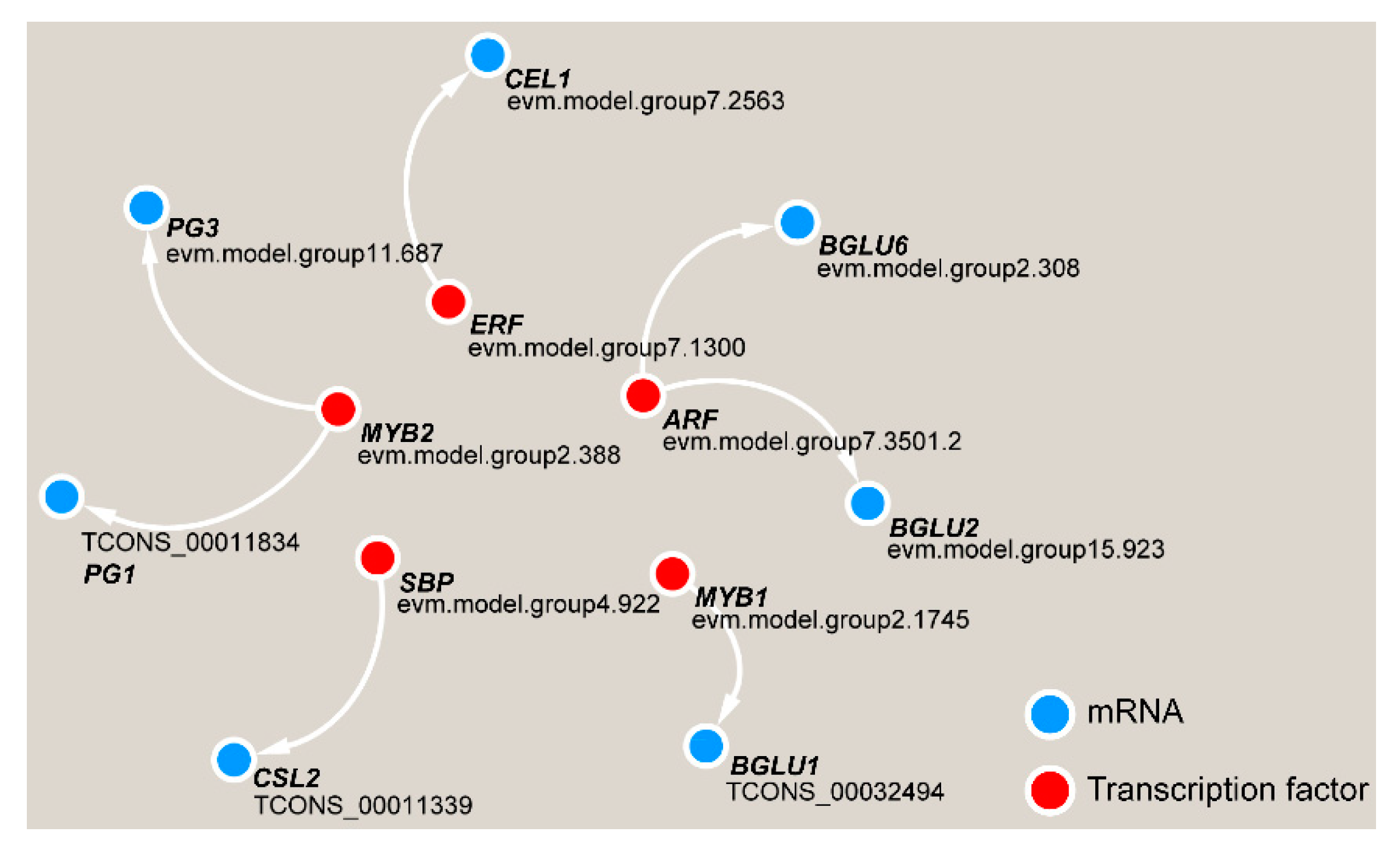
2.8. Transcriptional Regulation Network
3. Discussion
3.1. Anatomical Characteristics
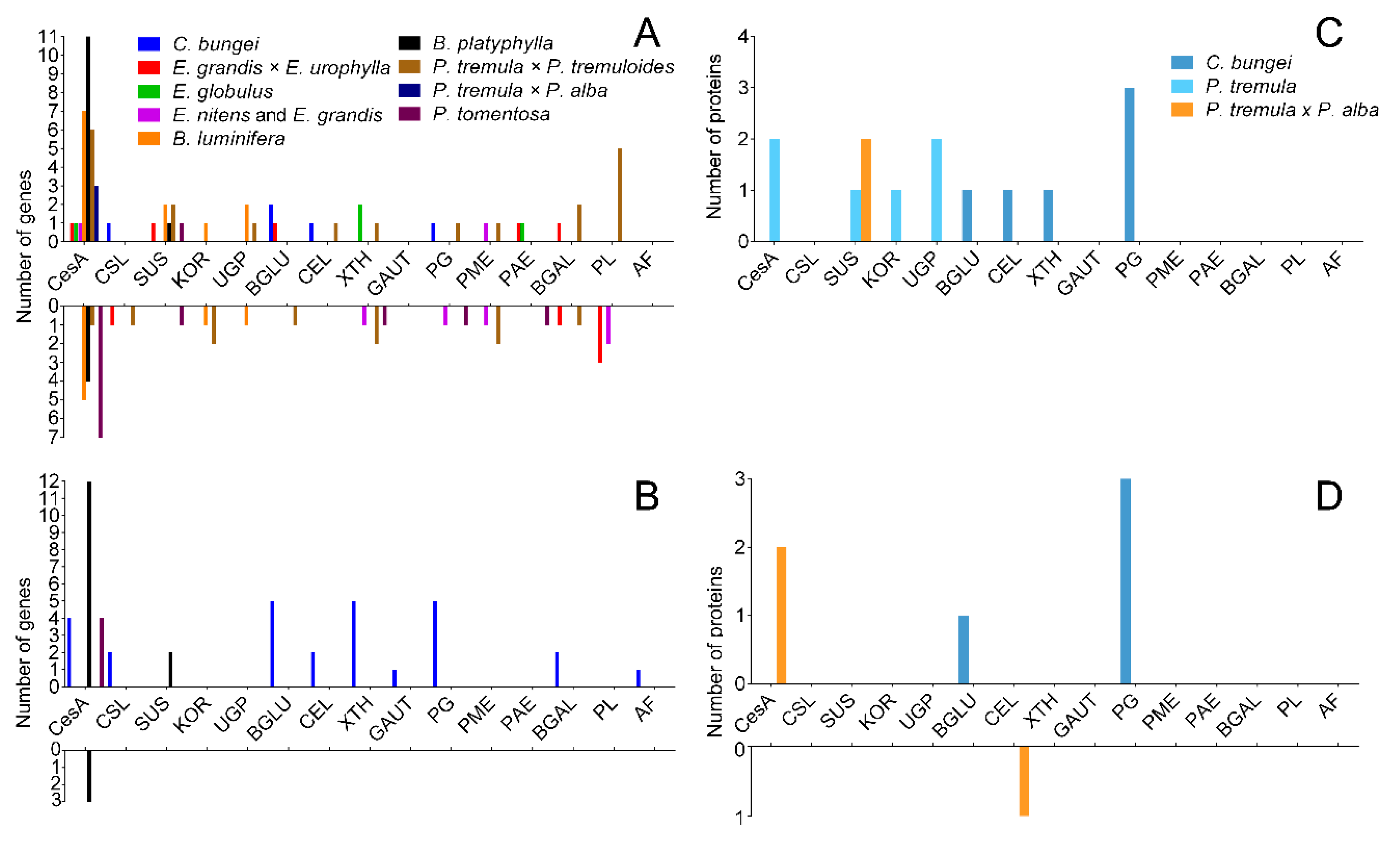
3.2. Cellulose and Hemicellulose Synthesis
3.3. Pectin Synthesis
3.4. Transcriptional Regulation in TW Formation
4. Materials and Methods
4.1. Plant Materials
4.2. Anatomy of Xylem
4.3. Transmission Electron Microscopy
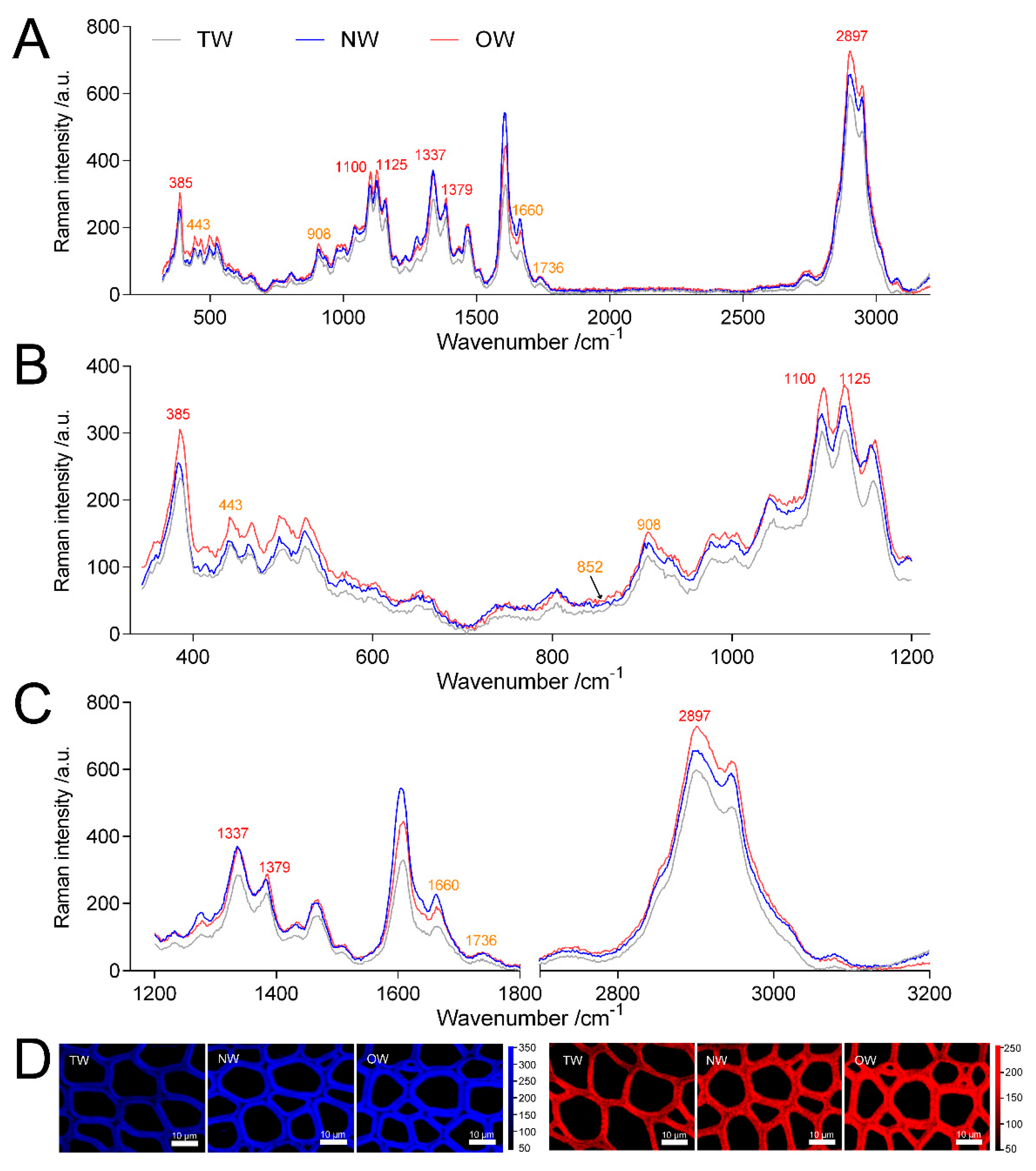
4.4. Confocal Raman Spectra and Raman Mapping
4.5. mRNA Isolation, Library Construction, and Illumina Transcriptome Sequencing
4.6. Quantification of Transcript Abundance and Differential Expression of mRNAs
4.7. Protein Extraction
4.8. Protein Digestion and TMT Labeling
4.9. High pH Reverse Phase Separation
4.10. Protein Identification and Quantification
4.11. Functional Enrichment of Differentially Expressed mRNAs and Proteins
4.12. Quantitative Real-Time (qRT)-PCR
4.13. Data Deposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ye, Z.H.; Zhong, R. Molecular control of wood formation in trees. J. Exp. Bot. 2015, 66, 4119–4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellgren, J.M.; Olofsson, K.; Sundberg, B. Patterns of auxin distribution during gravitational induction of reaction wood in poplar and pine. Plant Physiol. 2004, 135, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plomion, C.; Pionneau, C.; Brach, J.; Costa, P.; Baillères, H. Compression Wood-Responsive Proteins in Developing Xylem of Maritime Pine (Pinus pinaster Ait.). Plant Physiol. 2000, 123, 959–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilate, G.; Déjardin, A.; Laurans, F.; Leplé, J.-C. Tension wood as a model for functional genomics of wood formation. New Phytol. 2004, 164, 63–72. [Google Scholar] [CrossRef]
- Clair, B.; Gril, J.; Di Renzo, F.; Yamamoto, H.; Quignard, F. Characterization of a Gel in the Cell Wall To Elucidate the Paradoxical Shrinkage of Tension Wood. Biomacromolecules 2008, 9, 494–498. [Google Scholar] [CrossRef]
- Gritsch, C.; Wan, Y.; Mitchell, R.A.; Shewry, P.R.; Hanley, S.J.; Karp, A. G-fibre cell wall development in willow stems during tension wood induction. J. Exp. Bot. 2015, 66, 6447–6459. [Google Scholar] [CrossRef] [Green Version]
- Nishikubo, N.; Awano, T.; Banasiak, A.; Bourquin, V.; Ibatullin, F.; Funada, R.; Brumer, H.; Teeri, T.T.; Hayashi, T.; Sundberg, B.; et al. Xyloglucan endo-transglycosylase (XET) functions in gelatinous layers of tension wood fibers in poplar--a glimpse into the mechanism of the balancing act of trees. Plant Cell Physiol. 2007, 48, 843–855. [Google Scholar] [CrossRef] [Green Version]
- Clair, B.; Ruelle, J.; Beauchene, J.; Prévost, M.; Clair, B.; Fournier, M. Tension Wood and Opposite wood in 21 Tropical Rain Forest Species. IAWA J. 2006, 27, 341–376. [Google Scholar] [CrossRef] [Green Version]
- Yoshizawa, N.; Inami, A.; Miyake, S.; Ishiguri, F.; Yokota, S. Anatomy and lignin distribution of reaction wood in two Magnolia species. Wood Sci. Technol. 2000, 34, 183–196. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724–732. [Google Scholar] [CrossRef]
- Andersson-Gunnerås, S.; Mellerowicz, E.J.; Love, J.; Segerman, B.; Ohmiya, Y.; Coutinho, P.M.; Nilsson, P.; Henrissat, B.; Moritz, T.; Sundberg, B. Biosynthesis of cellulose-enriched tension wood in Populus: Global analysis of transcripts and metabolites identifies biochemical and developmental regulators in secondary wall biosynthesis. Plant J. 2006, 45, 144–165. [Google Scholar] [CrossRef]
- Foston, M.; Hubbell, C.A.; Samuel, R.; Jung, S.; Fan, H.; Ding, S.-Y.; Zeng, Y.; Jawdy, S.; Davis, M.; Sykes, R.; et al. Chemical, ultrastructural and supramolecular analysis of tension wood in Populus tremula x alba as a model substrate for reduced recalcitrance. Energ. Environ. Sci. 2011, 4, 4962. [Google Scholar] [CrossRef]
- Gierlinger, N.; Schwanninger, M. Chemical Imaging of Poplar Wood Cell Walls by Confocal Raman Microscopy. Plant Physiol. 2006, 140, 1246–1254. [Google Scholar] [CrossRef] [Green Version]
- McNeil, M.; Darvill, A.; Fry, S.; Albersheim, P. Structure and Function of the Primary Cell Walls of Plants. Annu. Rev. Biochem. 1984, 53, 625–663. [Google Scholar] [CrossRef]
- Robertsen, B. Elicitors of the production of lignin-like compounds in cucumber hypocotyls. Physiol. Mol. Plant Pathol. 1986, 28, 137–148. [Google Scholar] [CrossRef]
- Altaner, C.M.; Tokareva, E.N.; Jarvis, M.C.; Harris, P.J. Distribution of (1->4)- -galactans, arabinogalactan proteins, xylans and (1->3)- -glucans in tracheid cell walls of softwoods. Tree Physiol. 2010, 30, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Guedes, F.T.P.; Laurans, F.; Quemener, B.; Assor, C.; Laine-Prade, V.; Boizot, N.; Vigouroux, J.; Lesage-Descauses, M.C.; Leple, J.C.; Dejardin, A.; et al. Non-cellulosic polysaccharide distribution during G-layer formation in poplar tension wood fibers: Abundance of rhamnogalacturonan I and arabinogalactan proteins but no evidence of xyloglucan. Planta 2017, 246, 857–878. [Google Scholar] [CrossRef] [PubMed]
- Arend, M. Immunolocalization of (1,4)- -galactan in tension wood fibers of poplar. Tree Physiol. 2008, 28, 1263–1267. [Google Scholar] [CrossRef]
- Christiaens, S.; Van Buggenhout, S.; Ngouémazong, E.D.; Vandevenne, E.; Fraeye, I.; Duvetter, T.; Van Loey, A.M.; Hendrickx, M.E. Anti-homogalacturonan antibodies: A way to explore the effect of processing on pectin in fruits and vegetables? Food Res. Int. 2011, 44, 225–234. [Google Scholar] [CrossRef]
- Verhertbruggen, Y.; Marcus, S.E.; Haeger, A.; Ordaz-Ortiz, J.J.; Knox, J.P. An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr. Res. 2009, 344, 1858–1862. [Google Scholar] [CrossRef] [PubMed]
- Bowling, A.J.; Vaughn, K.C. Immunocytochemical characterization of tension wood: Gelatinous fibers contain more than just cellulose. Am. J. Bot. 2008, 95, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Mizrachi, E.; Maloney, V.; Silberbauer, J.; Hefer, C.; Berger, D.; Mansfield, S.; Myburg, A. Investigating the molecular underpinnings underlying morphology and changes in carbon partitioning during tension wood formation in Eucalyptus. New Phytol. 2015, 206, 1351–1363. [Google Scholar] [CrossRef]
- Xiao, Y.; Ma, W.; Lu, N.; Wang, Z.; Wang, N.; Zhai, W.; Kong, L.; Qu, G.; Wang, Q.; Wang, J. Genetic Variation of Growth Traits and Genotype-by-Environment Interactions in Clones of Catalpa bungei and Catalpa fargesii f. duclouxii. Forests 2019, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Socrates, G. Infrared and Raman Characteristic Group Frequencies, 3rd ed.; J. Wiley & Sons: Chichester, UK; New York, NY, USA; Weinheim, Germany; Toronto, ON, Canada, 2001; Chapter 2. [Google Scholar]
- Szymańska-Chargot, M.; Cybulska, M.; Zdunek, A. Sensing the structural differences in cellulose from apple and bacterial cell wall materials by Raman and FT-IR spectroscopy. Sensors 2011, 11, 5543–5560. [Google Scholar] [CrossRef] [Green Version]
- Wróbel-Kwiatkowska, M.; Żuk, M.; Szopa, J.; Dymińska, L.; Mączka, M.; Hanuza, J. Poly-3-hydroxy butyric acid interaction with the transgenic flax fibers: FT-IR and Raman spectra of the composite extracted from a GM flax. Spectrochim. Acta A 2009, 73, 286–294. [Google Scholar] [CrossRef]
- Synytsya, A. Fourier transform Raman and infrared spectroscopy of pectins. Carbohyd. Polym. 2003, 54, 97–106. [Google Scholar] [CrossRef]
- Jin, K.; Liu, X.; Wang, K.; Jiang, Z.; Tian, G.; Yang, S.; Shang, L.; Ma, J. Imaging the dynamic deposition of cell wall polymer in xylem and phloem in Populus x euramericana. Planta 2018, 248, 849–858. [Google Scholar] [CrossRef]
- Agarwal, U.P.; Ralph, S.A. FT-Raman Spectroscopy of Wood: Identifying Contributions of Lignin and Carbohydrate Polymers in the Spectrum of Black Spruce (Picea Mariana). Appl. Spectrosc. 2016, 51, 1648–1655. [Google Scholar] [CrossRef]
- Edwards, H.; Farwell, D.; Webster, D. FT Raman microscopy of untreated natural plant fibres. Spectrochim. Acta. A 1997, 53A, 2383–2392. [Google Scholar] [CrossRef]
- Paux, E.; Carocha, V.; Marques, C.; Mendes de Sousa, A.; Borralho, N.; Sivadon, P.; Grima-Pettenati, J. Transcript profiling of Eucalyptus xylem genes during tension wood formation. New Phytol. 2005, 167, 89–100. [Google Scholar] [CrossRef]
- Qiu, D.; Wilson, I.W.; Gan, S.; Washusen, R.; Moran, G.F.; Southerton, S.G. Gene expression in Eucalyptus branch wood with marked variation in cellulose microfibril orientation and lacking G-layers. New Phytol. 2008, 179, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Huang, H.; Ni, F.; Tong, Z.; Lin, E.; Zhu, M. RNA-Seq analysis of differential gene expression in Betula luminifera xylem during the early stages of tension wood formation. Peer J. 2018, 6, e5427. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, N.; Gao, C.; Cui, Z.; Sun, D.; Yang, C.; Wang, Y. Comprehensive Transcriptome Analysis of Developing Xylem Responding to Artificial Bending and Gravitational Stimuli in Betula platyphylla. PLoS ONE 2014, 9, e87566. [Google Scholar] [CrossRef]
- Azri, W.; Ennajah, A.; Nasr, Z.; Woo, S.-Y.; Khaldi, A. Transcriptome profiling the basal region of poplar stems during the early gravitropic response. Biol. Plant. 2014, 58, 55–63. [Google Scholar] [CrossRef]
- Bygdell, J.; Srivastava, V.; Obudulu, O.; Srivastava, M.K.; Nilsson, R.; Sundberg, B.; Trygg, J.; Mellerowicz, E.J.; Wingsle, G. Protein expression in tension wood formation monitored at high tissue resolution in Populus. J. Exp. Bot. 2017, 68, 3405–3417. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chen, B.; Zhang, D. Transcript profiling of Populus tomentosa genes in normal, tension, and opposite wood by RNA-seq. BMC Genom. 2015, 16, 164. [Google Scholar] [CrossRef] [Green Version]
- Mauriat, M.; Leplé, J.-C.; Claverol, S.; Bartholomé, J.; Negroni, L.; Richet, N.; Lalanne, C.; Bonneu, M.; Coutand, C.; Plomion, C. Quantitative Proteomic and Phosphoproteomic Approaches for Deciphering the Signaling Pathway for Tension Wood Formation in Poplar. J. Proteome Res. 2015, 14, 3188–3203. [Google Scholar] [CrossRef]
- Clair, B.; Déjardin, A.; Pilate, G.; Alméras, T. Is the G-Layer a Tertiary Cell Wall? Front. Plant Sci. 2018, 9, 623. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.S.; Clair, B.; Ruelle, J.; Beauchene, J.; Di Renzo, F.; Quignard, F.; Zhao, G.J.; Yamamoto, H.; Gril, J. Mesoporosity as a new parameter for understanding tension stress generation in trees. J. Exp. Bot. 2009, 60, 3023–3030. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.; Stevenson, J. Occurrence of Reaction Wood in Branches of Dicotyledons and Its Role in Tree Architecture. Bot. Gaz. 1981, 142, 82–95. [Google Scholar] [CrossRef]
- Roussel, J.R.; Clair, B. Evidence of the late lignification of the G-layer in Simarouba tension wood, to assist understanding how non-G-layer species produce tensile stress. Tree Physiol. 2015, 35, 1366–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endler, A.; Persson, S. Cellulose Synthases and Synthesis in Arabidopsis. Mol. Plant 2011, 4, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Haigler, C.H.; Ivanova-Datcheva, M.; Hogan, P.S.; Salnikov, V.V.; Hwang, S.; Martin, K.; Delmer, D.P. Carbon partitioning to cellulose synthesis. Plant. Mol. Biol. 2001, 47, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Dhugga, K. Building the wall: Genes and enzyme complexes for polysaccharide synthases. Curr. Opin. Plant Biol. 2001, 4, 488–493. [Google Scholar] [CrossRef]
- Liepman, A.; Wilkerson, C.; Keegstra, K. Expression of cellulose synthase-like (Csl) genes in insect cells reveals that CslA family members encode mannan synthases. Proc. Natl. Acad. Sci. USA 2005, 102, 2221–2226. [Google Scholar] [CrossRef] [Green Version]
- Bernal, A.J.; Yoo, C.-M.; Mutwil, M.; Jensen, J.K.; Hou, G.; Blaukopf, C.; Sørensen, I.; Blancaflor, E.B.; Scheller, H.V.; Willats, W.G.T. Functional Analysis of the Cellulose Synthase-Like Genes CSLD1, CSLD2, and CSLD4 in Tip-Growing Arabidopsis Cells. Plant Physiol. 2008, 148, 1238–1253. [Google Scholar] [CrossRef] [Green Version]
- Mellerowicz, E.J.; Peter, I.; Takahisa, H. Xyloglucan: The Molecular Muscle of Trees. Ann. Bot. 2008, 102, 659–665. [Google Scholar] [CrossRef] [Green Version]
- Fagerstedt, K.V.; Mellerowicz, E.; Gorshkova, T.; Ruel, K.; Joseleau, J.P. Cell Wall Polymers in Reaction Wood; Springer: Berlin/Heidelberg, Germany, 2014; pp. 37–106. [Google Scholar]
- Chang, S.S.; Shi, Y.; Liu, T.; Hu, J.B. Anatomical structure and structure characteristic of chemical composition of gelatinous layer in tension wood. Sci. Silva Sin. 2018, 54, 153–161. [Google Scholar]
- Libertini, E.; Li, Y.; McQueen-Mason, S.J. Phylogenetic Analysis of the Plant Endo-β-1,4-Glucanase Gene Family. J. Mol. Evol. 2004, 58, 506–515. [Google Scholar] [CrossRef]
- Voragen, A.G.J.; Coenen, G.-J.; Verhoef, R.P.; Schols, H.A. Pectin, a versatile polysaccharide present in plant cell walls. Struct. Chem. 2009, 20, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, W.; Wang, Y.Q.; Liu, Y.Y.; Wang, J.X.; Zhang, X.Q.; Ye, D.; Chen, L.Q. Arabidopsis galacturonosyltransferase (GAUT) 13 and GAUT14 have redundant functions in pollen tube growth. Mol. Plant 2013, 6, 1131–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.; Yadav, P.K.; Yadav, D.; Yadav, K.D.S. Pectin lyase: A review. Process Biochem. 2009, 44, 1–10. [Google Scholar] [CrossRef]
- Chang, S.-S.; Quignard, F.; Alméras, T.; Clair, B. Mesoporosity changes from cambium to mature tension wood: A new step toward the understanding of maturation stress generation in trees. New Phytol. 2015, 205, 1277–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, L.; Dunlop, J.W.; Jungnikl, K.; Eder, M.; Gierlinger, N.; Coutand, C.; Jeronimidis, G.; Fratzl, P.; Burgert, I. Stress generation in the tension wood of poplar is based on the lateral swelling power of the G-layer. Plant J. 2008, 56, 531–538. [Google Scholar] [CrossRef]
- Zhong, R.; McCarthy, R.L.; Lee, C.; Ye, Z.H. Dissection of the transcriptional program regulating secondary wall biosynthesis during wood formation in poplar. Plant Physiol. 2011, 157, 1452–1468. [Google Scholar] [CrossRef] [Green Version]
- Du, S.; Uno, H.; Yamamoto, F. Roles of Auxin and gibberellin in gravity-induced tension wood formation in Aesculus Turbinata seedlings. IAWA J. 2004, 25, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.; Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 12:323. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Aoki-Kinoshita, K.; Kanehisa, M. Gene Annotation and Pathway Mapping in KEGG. Methods Mol. Biol. 2007, 396, 71–91. [Google Scholar]
- Jing, D.; Xia, Y.; Chen, F.; Wang, Z.; Zhang, S.; Wang, J. Ectopic expression of a Catalpa bungei (Bignoniaceae) PISTILLATA homologue rescues the petal and stamen identities in Arabidopsis pi-1 mutant. Plant Sci. 2015, 231, 40–51. [Google Scholar] [CrossRef] [PubMed]












© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; Yi, F.; Ling, J.; Wang, Z.; Zhao, K.; Lu, N.; Qu, G.; Kong, L.; Ma, W.; Wang, J. Transcriptomics and Proteomics Reveal the Cellulose and Pectin Metabolic Processes in the Tension Wood (Non-G-Layer) of Catalpa bungei. Int. J. Mol. Sci. 2020, 21, 1686. https://doi.org/10.3390/ijms21051686
Xiao Y, Yi F, Ling J, Wang Z, Zhao K, Lu N, Qu G, Kong L, Ma W, Wang J. Transcriptomics and Proteomics Reveal the Cellulose and Pectin Metabolic Processes in the Tension Wood (Non-G-Layer) of Catalpa bungei. International Journal of Molecular Sciences. 2020; 21(5):1686. https://doi.org/10.3390/ijms21051686
Chicago/Turabian StyleXiao, Yao, Fei Yi, Juanjuan Ling, Zhi Wang, Kun Zhao, Nan Lu, Guanzheng Qu, Lisheng Kong, Wenjun Ma, and Junhui Wang. 2020. "Transcriptomics and Proteomics Reveal the Cellulose and Pectin Metabolic Processes in the Tension Wood (Non-G-Layer) of Catalpa bungei" International Journal of Molecular Sciences 21, no. 5: 1686. https://doi.org/10.3390/ijms21051686
APA StyleXiao, Y., Yi, F., Ling, J., Wang, Z., Zhao, K., Lu, N., Qu, G., Kong, L., Ma, W., & Wang, J. (2020). Transcriptomics and Proteomics Reveal the Cellulose and Pectin Metabolic Processes in the Tension Wood (Non-G-Layer) of Catalpa bungei. International Journal of Molecular Sciences, 21(5), 1686. https://doi.org/10.3390/ijms21051686