Mitochondrial Epigenetics: Non-Coding RNAs as a Novel Layer of Complexity

 , and
, and
Abstract
:1. Introduction
2. Mitochondrial Functions
3. Nuclei–Mitochondria Communication
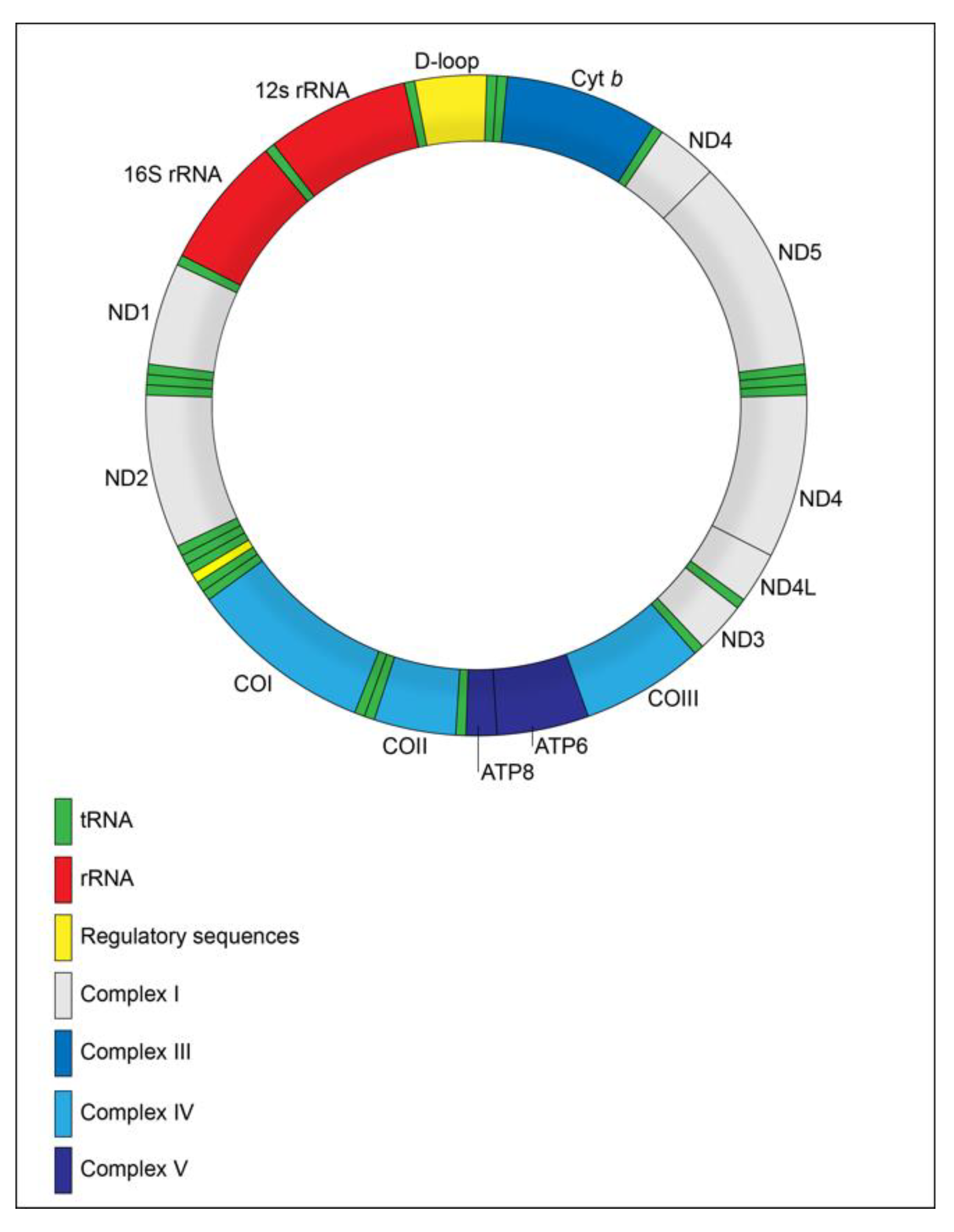
4. Mitochondrial Genome
5. Epigenetics
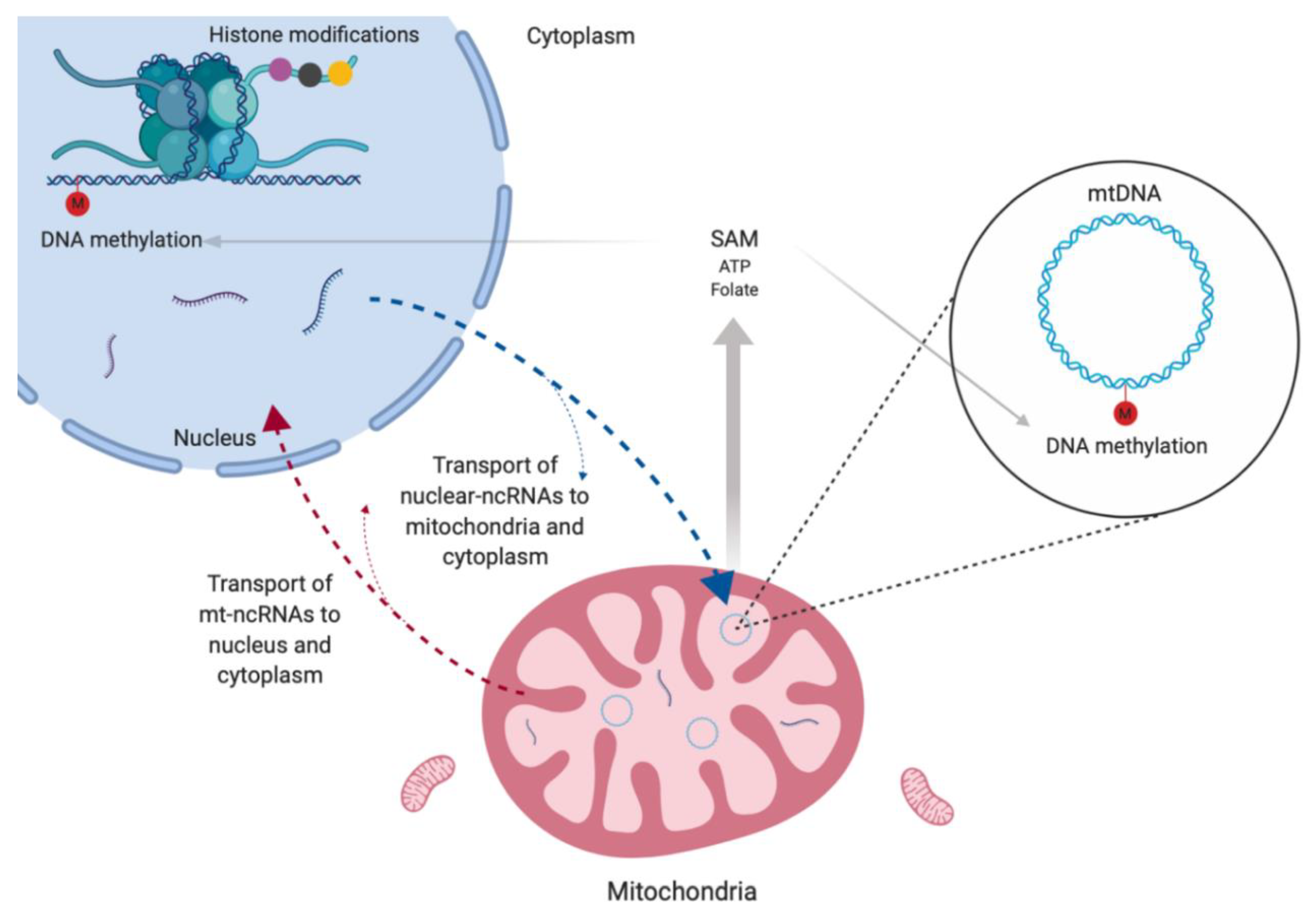
6. Mitochondrial Epigenetics
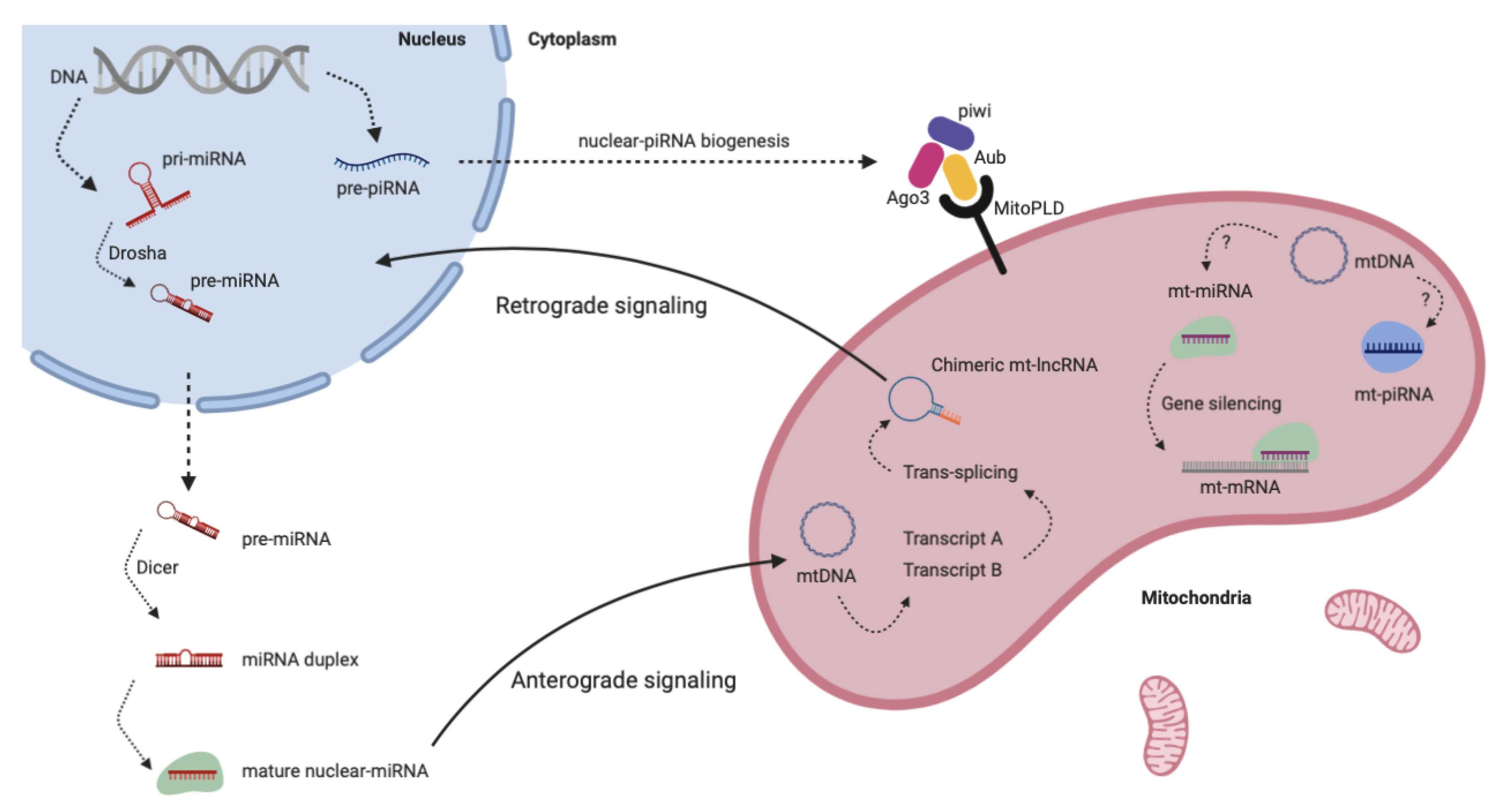
7. Non-Coding RNAs in Mitochondria
7.1. Linear Long Non-coding RNAs
7.2. Circular RNAs
7.3. miRNAs
7.4. piRNAs
8. Non-coding RNAs and Mitochondrial Diseases
9. Perspectives
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| OXPHOS | oxidative phosphorylation |
| mtGenome | mitochondrial genome |
| mtDNA | mitochondrial DNA |
| nDNA | nuclear DNA |
| ncRNAs | non-coding RNAs |
| ETC | electron transport chain |
| ROS | reactive oxygen species |
| ADP | adenosine diphosphate |
| ATP | adenosine triphosphate |
| MOMP | mitochondrial outer membrane permeabilization |
| TFAM | transcription and mtDNA maintenance factor |
| HIF1 | hypoxia inducible factor 1 |
| SIRT | sirtuins |
| tRNAs | transfer RNAs |
| rRNAs | ribosomal RNAs |
| D-loop | displacement loop |
| DNMTs | DNA-methyltransferases |
| CpG sites | cytosine-guanine dinucleotides |
| lncRNAs | linear long non-coding RNAs |
| circRNAs | circular RNAs |
| miRNAs | microRNA |
| piRNAs | piwi-interacting RNAs |
| nuclear-ncRNAs | nuclear-encoded non-coding RNAs |
| mt-ncRNAs | mitochondrial encoded non-coding RNAs |
| HuR | human anti-gen R |
| GRSF1 | G-rich RNA sequence-binding factor 1 |
| RNase MRP | RNA processing endoribonuclease |
| SRA | steroid receptor RNA |
| RBP | RNA binding protein |
| Ago2 | Argonaute |
| RISC | RNA-induced silencing complex |
| RNAi | RNA interference |
| Aub | aubergine |
| Zuc | zucchini |
| MELAS | Mitochondrial encephalopathy, lactic acidosis, and stroke-like episodes |
| MERRF | myoclonic epilepsy with ragged-red fibers |
| lincRNA | long intergenic non-coding RNAs |
References
- Shaughnessy, D.T.; McAllister, K.; Worth, L.; Haugen, A.C.; Meyer, J.N.; Domann, F.E.; Van Houten, B.; Mostoslavsky, R.; Bultman, S.J.; Baccarelli, A.A.; et al. Mitochondria, Energetics, Epigenetics, and Cellular Responses to Stress. Environ. Health Perspect. 2014, 122, 1271–1278. [Google Scholar] [CrossRef]
- Sagan, L. On the origin of mitosing cells. J. Theor. Biol. 1967, 14, 225–IN6. [Google Scholar] [CrossRef]
- Yang, D.; Oyaizu, Y.; Oyaizu, H.; Olsen, G.J.; Woese, C.R. Mitochondrial origins. Proc. Natl. Acad. Sci. USA 1985, 82, 4443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, A.J.; Munoz-Gomez, S.A.; Kamikawa, R. The origin and diversification of mitochondria. Curr. Biol. 2017, 27, R1177–R1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manev, H.; Dzitoyeva, S. Progress in mitochondrial epigenetics. Biomol Concepts 2013, 4, 381–389. [Google Scholar] [CrossRef]
- Burke, P.J. Mitochondria, Bioenergetics and Apoptosis in Cancer. Trends Cancer 2017, 3, 857–870. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 7th ed.; Freeman, W.H. & Company: New York, NY, USA, 2017. [Google Scholar]
- Sousa, J.S.; D’Imprima, E.; Vonck, J. Mitochondrial respiratory chain complexes. In Membrane Protein Complexes: Structure and Function; Springer: Basel, Switzerland, 2018; pp. 167–227. [Google Scholar]
- Chipuk, J.E.; Bouchier-Hayes, L.; Green, D.R. Mitochondrial outer membrane permeabilization during apoptosis: The innocent bystander scenario. Cell Death Differ. 2006, 13, 1396. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, G.C.; Schaan, A.P.; Cabral, G.F.; Santana-da-Silva, M.N.; Pinto, P.; Vidal, A.F.; Ribeiro-dos-Santos, Â. A Cell’s Fate: An Overview of the Molecular Biology and Genetics of Apoptosis. Int. J. Mol. Sci. 2019, 20, 4133. [Google Scholar] [CrossRef] [Green Version]
- Van Gisbergen, M.W.; Voets, A.M.; Starmans, M.H.W.; de Coo, I.F.M.; Yadak, R.; Hoffmann, R.F.; Boutros, P.C.; Smeets, H.J.M.; Dubois, L.; Lambin, P. How do changes in the mtDNA and mitochondrial dysfunction influence cancer and cancer therapy? Challenges, opportunities and models. Mutat. Res. /Rev. Mutat. Res. 2015, 764, 16–30. [Google Scholar] [CrossRef]
- Enns, G.M. The contribution of mitochondria to common disorders. Mol. Genet. Metab. 2003, 80, 11–26. [Google Scholar] [CrossRef]
- Bose, A.; Beal, M.F. Mitochondrial dysfunction in Parkinson’s disease. J. Neurochem. 2016, 139, 216–231. [Google Scholar] [CrossRef] [PubMed]
- Oliver, D.; Reddy, P.H. Molecular Basis of Alzheimer’s Disease: Focus on Mitochondria. J. Alzheimer’s Dis. 2019, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, R.; Nakahira, K.; Gu, Z. Mitochondrial DNA Mutation, Diseases, and Nutrient-Regulated Mitophagy. Annu. Rev. Nutr. 2019, 39, 201–226. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yoshitomi, T.; Hu, J.-F.; Cui, J. Long noncoding RNAs coordinate functions between mitochondria and the nucleus. Epigenetics Chromatin 2017, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, S.E.; Sena, L.A.; Chandel, N.S. Mitochondria in the regulation of innate and adaptive immunity. Immunity 2015, 42, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpulla, R.C. Nuclear control of respiratory gene expression in mammalian cells. J. Cell. Biochem. 2006, 97, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.T.; Hoogenraad, N.J. Mitochondrial-nuclear communications. Annu. Rev. Biochem. 2007, 76, 701–722. [Google Scholar] [CrossRef]
- Vendramin, R.; Marine, J.; Leucci, E. Non-coding RNAs: The dark side of nuclear–mitochondrial communication. Embo J. 2017, 36, 1123–1133. [Google Scholar] [CrossRef]
- De Souza, A.; Wang, J.-Z.; Dehesh, K. Retrograde signals: Integrators of interorganellar communication and orchestrators of plant development. Annu. Rev. Plant. Biol. 2017, 68, 85–108. [Google Scholar] [CrossRef]
- Verdin, E.; Hirschey, M.D.; Finley, L.W.S.; Haigis, M.C. Sirtuin regulation of mitochondria: Energy production, apoptosis, and signaling. Trends Biochem. Sci. 2010, 35, 669–675. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Sun, L.; Wang, R.R.; Hu, J.-F.; Cui, J. The effects of mitochondria-associated long noncoding RNAs in cancer mitochondria: New players in an old arena. Crit. Rev. Oncol. /Hematol. 2018, 131, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.W.; Turnbull, D.M. Mitochondrial DNA mutations in human disease. Nat. Rev. Genet. 2005, 6, 389–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-H.; Kim, D.-K. Microsatellite Instability of Nuclear and Mitochondrial DNAs in Gastric Carcinogenesis. Asian Pac. J. Cancer Prev. 2014, 15, 8027–8034. [Google Scholar] [CrossRef] [Green Version]
- Snustad, D.P.; Simmons, M.J. Principles of Genetics, Binder ready Version; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- He, Y.; Wu, J.; Dressman, D.C.; Iacobuzio-Donahue, C.; Markowitz, S.D.; Velculescu, V.E.; Diaz, L.A., Jr.; Kinzler, K.W.; Vogelstein, B.; Papadopoulos, N. Heteroplasmic mitochondrial DNA mutations in normal and tumour cells. Nature 2010, 464, 610–614. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, R.; Faustin, B.; Rocher, C.; Malgat, M.; Mazat, J.-P.; Letellier, T. Mitochondrial threshold effects. Biochem. J. 2003, 370, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.A. The human genome. In Genomes, 2nd ed.; Wiley-Liss: Hoboken, NJ, USA, 2002. [Google Scholar]
- Neiman, M.; Taylor, D.R. The causes of mutation accumulation in mitochondrial genomes. Proc. R. Soc. B: Biol. Sci. 2009, 276, 1201–1209. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Lee, J.-H.; Kim, D.-C.; Hwang, I.; Kang, Y.-N.; Gwon, G.-J.; Choi, I.-J.; Kim, S. Is Mitochondrial DNA Copy Number Associated with Clinical Characteristics and Prognosis in Gastric Cancer? Asian Pac. J. Cancer Prev. 2015, 16, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Weigl, S.; Paradiso, A.; Tommasi, S. Mitochondria and Familial Predisposition to Breast Cancer. Curr. Genom. 2013, 14, 195–203. [Google Scholar] [CrossRef]
- Kolesnikov, A.A. The Mitochondrial Genome. The Nucleoid. Biochem. Mosc. 2016, 81, 1057–1065. [Google Scholar] [CrossRef]
- Luo, S.; Valencia, C.A.; Zhang, J.; Lee, N.-C.; Slone, J.; Gui, B.; Wang, X.; Li, Z.; Dell, S.; Brown, J.; et al. Biparental Inheritance of Mitochondrial DNA in Humans. Proc. Natl. Acad. Sci. 2018, 115, 13039–13044. [Google Scholar] [CrossRef] [Green Version]
- Waddington, C.H. The epigenotype. Int. J. Epidemiol. 2011, 41, 10–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Rao, C.M. Epigenetic tools (The Writers, The Readers and The Erasers) and their implications in cancer therapy. Eur. J. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; He, X. Integrating Epigenomics into the Understanding of Biomedical Insight. Bioinform Biol Insights 2016, 10, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Romani, M.; Pistillo, M.P.; Banelli, B. Environmental epigenetics: Crossroad between public health, lifestyle, and cancer prevention. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, A.-T.; Robaa, D.; Jung, M. Epigenetic small molecule modulators of histone and DNA methylation. Curr. Opin. Chem. Biol. 2018, 45, 73–85. [Google Scholar] [CrossRef]
- Morris, K.V.; Mattick, J.S. The rise of regulatory RNA. Nat. Rev. Genet. 2014, 15, 423. [Google Scholar] [CrossRef] [Green Version]
- D’Aquila, P.; Bellizzi, D.; Passarino, G. Mitochondria in health, aging and diseases: The epigenetic perspective. Biogerontology 2015, 16, 569–585. [Google Scholar] [CrossRef]
- Iacobazzi, V.; Castegna, A.; Infantino, V.; Andria, G. Mitochondrial DNA methylation as a next-generation biomarker and diagnostic tool. Mol. Genet. Metab. 2013, 110, 25–34. [Google Scholar] [CrossRef]
- Matilainen, O.; Quirós, P.M.; Auwerx, J. Mitochondria and Epigenetics - Crosstalk in Homeostasis and Stress. Trends Cell Biol. 2017, 27, 453–463. [Google Scholar] [CrossRef]
- Mposhi, A.; Van der Wijst, M.; Faber, K.N.; Rots, M.G. Regulation of mitochondrial gene expression, the epigenetic enigma. Front. Biosci. (Landmark Ed.) 2017, 22, 1099–1113. [Google Scholar]
- Choi, Y.-S.; Jeong, J.H.; Min, H.-K.; Jung, H.-J.; Hwang, D.; Lee, S.-W.; Pak, Y.K. Shot-gun proteomic analysis of mitochondrial D-loop DNA binding proteins: Identification of mitochondrial histones. Mol. Biosyst. 2011, 7, 1523–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, T.R.; Neph, S.; Dinger, M.E.; Crawford, J.; Smith, M.A.; Shearwood, A.-M.J.; Haugen, E.; Bracken, C.P.; Rackham, O.; Stamatoyannopoulos, J.A.; et al. The human mitochondrial transcriptome. Cell 2011, 146, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, F.; Palmeira, C.; Rolo, A. The role of microRNAs in mitochondria: Small players acting wide. Genes 2014, 5, 865–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas, J.; Zárraga, A.M.; Muller, I.; Montecinos, L.; Werner, E.; Brito, M.; Meneses, A.M.; Burzio, L.O. A novel chimeric mitochondrial RNA localized in the nucleus of mouse sperm. Dna Cell Biol. 2000, 19, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Rackham, O.; Shearwood, A.-M.J.; Mercer, T.R.; Davies, S.M.; Mattick, J.S.; Filipovska, A. Long noncoding RNAs are generated from the mitochondrial genome and regulated by nuclear-encoded proteins. Rna 2011, 17, 2085–2093. [Google Scholar] [CrossRef] [Green Version]
- Villegas, J.; Müller, I.; Arredondo, J.; Pinto, R.; Burzio, L.O. A putative RNA editing from U to C in a mouse mitochondrial transcript. Nucleic Acids Res. 2002, 30, 1895–1901. [Google Scholar] [CrossRef] [Green Version]
- Villegas, J.; Araya, P.; Bustos-Obregon, E.; Burzio, L.O. Localization of the 16S mitochondrial rRNA in the nucleus of mammalian spermatogenic cells. Mol. Hum. Reprod. 2002, 8, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Villegas, J.; Burzio, V.; Villota, C.; Landerer, E.; Martinez, R.; Santander, M.; Martinez, R.; Pinto, R.; Vera, M.I.; Boccardo, E.; et al. Expression of a novel non-coding mitochondrial RNA in human proliferating cells. Nucleic Acids Res. 2007, 35, 7336–7347. [Google Scholar] [CrossRef] [Green Version]
- Burzio, V.A.; Villota, C.; Villegas, J.; Landerer, E.; Boccardo, E.; Villa, L.L.; Martínez, R.; Lopez, C.; Gaete, F.; Toro, V.; et al. Expression of a family of noncoding mitochondrial RNAs distinguishes normal from cancer cells. Proc. Natl. Acad. Sci. USA 2009, 106, 9430–9434. [Google Scholar] [CrossRef] [Green Version]
- Borgna, V.; Villegas, J.; Burzio, V.A.; Belmar, S.; Araya, M.; Jeldes, E.; Lobos-González, L.; Silva, V.; Villota, C.; Oliveira-Cruz, L.; et al. Mitochondrial ASncmtRNA-1 and ASncmtRNA-2 as potent targets to inhibit tumor growth and metastasis in the RenCa murine renal adenocarcinoma model. Oncotarget 2017, 8, 43692–43708. [Google Scholar] [CrossRef] [Green Version]
- Vidaurre, S.; Fitzpatrick, C.; Burzio, V.A.; Briones, M.; Villota, C.; Villegas, J.; Echenique, J.; Oliveira-Cruz, L.; Araya, M.; Borgna, V.; et al. Down-regulation of the antisense mitochondrial non-coding RNAs (ncRNAs) is a unique vulnerability of cancer cells and a potential target for cancer therapy. J. Biol. Chem. 2014, 289, 27182–27198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landerer, E.; Villegas, J.; Burzio, V.A.; Oliveira, L.; Villota, C.; Lopez, C.; Restovic, F.; Martinez, R.; Castillo, O.; Burzio, L.O. Nuclear localization of the mitochondrial ncRNAs in normal and cancer cells. Cell. Oncol. 2011, 34, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Bianchessi, V.; Badi, I.; Bertolotti, M.; Nigro, P.; D’Alessandra, Y.; Capogrossi, M.C.; Zanobini, M.; Pompilio, G.; Raucci, A.; Lauri, A. The mitochondrial lncRNA ASncmtRNA-2 is induced in aging and replicative senescence in Endothelial Cells. J. Mol. Cell. Cardiol. 2015, 81, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Villota, C.; Campos, A.; Vidaurre, S.; Oliveira-Cruz, L.; Boccardo, E.; Burzio, V.A.; Varas, M.; Villegas, J.; Villa, L.L.; Valenzuela, P.D.; et al. Expression of mitochondrial non-coding RNAs (ncRNAs) is modulated by high risk human papillomavirus (HPV) oncogenes. J. Biol. Chem. 2012, 287, 21303–21315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.-C.; Yamada, K.A.; Patel, A.Y.; Topkara, V.K.; George, I.; Cheema, F.H.; Ewald, G.A.; Mann, D.L.; Nerbonne, J.M. Deep RNA sequencing reveals dynamic regulation of myocardial noncoding RNAs in failing human heart and remodeling with mechanical circulatory support. Circulation 2014, 129, 1009–1021. [Google Scholar] [CrossRef]
- Noh, J.H.; Kim, K.M.; Abdelmohsen, K.; Yoon, J.-H.; Panda, A.C.; Munk, R.; Kim, J.; Curtis, J.; Moad, C.A.; Wohler, C.M.; et al. HuR and GRSF1 modulate the nuclear export and mitochondrial localization of the lncRNA RMRP. Genes Dev. 2016, 30, 1224–1239. [Google Scholar]
- Li, K.; Smagula, C.S.; Parsons, W.J.; Richardson, J.A.; Gonzalez, M.; Hagler, H.K.; Williams, R.S. Subcellular partitioning of MRP RNA assessed by ultrastructural and biochemical analysis. J. Cell Biol. 1994, 124, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Colley, S.M.; Iyer, K.R.; Leedman, P.J. The RNA coregulator SRA, its binding proteins and nuclear receptor signaling activity. Iubmb Life 2008, 60, 159–164. [Google Scholar] [CrossRef]
- Doersen, C.-J.; Guerrier-Takada, C.; Altman, S.; Attardi, G. Characterization of an RNase P activity from HeLa cell mitochondria. Comparison with the cytosol RNase P activity. J. Biol. Chem. 1985, 260, 5942–5949. [Google Scholar]
- Wang, G.; Chen, H.-W.; Oktay, Y.; Zhang, J.; Allen, E.L.; Smith, G.M.; Fan, K.C.; Hong, J.S.; French, S.W.; McCaffery, J.M.; et al. PNPASE regulates RNA import into mitochondria. Cell 2010, 142, 456–467. [Google Scholar] [CrossRef] [Green Version]
- Halbreich, A.; Pajot, P.; Foucher, M.; Grandchamp, C.; Slonimski, P. A pathway of cytochrome b mRNA processing in yeast mitochondria: Specific splicing steps and an intron-derived circular RNA. Cell 1980, 19, 321–329. [Google Scholar] [CrossRef]
- Arnberg, A.; Van Ommen, G.-J.; Grivell, L.; Van Bruggen, E.; Borst, P. Some yeast mitochondrial RNAs are circular. Cell 1980, 19, 313–319. [Google Scholar] [CrossRef]
- Schroeder, R.; Breitenbach, M.; Schweyen, R.J. Mitochondria circular RN As are absent in sporulating cells of Saccharomyces cerevisiae. Nucleic Acids Res. 1983, 11, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Li-Pook-Than, J.; Bonen, L. Multiple physical forms of excised group II intron RNAs in wheat mitochondria. Nucleic Acids Res. 2006, 34, 2782–2790. [Google Scholar] [CrossRef] [Green Version]
- Darbani, B.; Noeparvar, S.; Borg, S. Identification of circular RNAs from the parental genes involved in multiple aspects of cellular metabolism in barley. Front. Plant. Sci. 2016, 7, 776. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wu, M.; Fan, Y.; Li, S.; Lai, Z.; Huang, Y.; Lan, X.; Lei, C.; Chen, H.; Dang, R. Identification and characterization of circular RNAs in Qinchuan cattle testis. R. Soc. Open Sci. 2018, 5, 180413. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Gan, T.-Y.; Li, N.; Liu, C.-Y.; Zhou, L.-Y.; Gao, J.-N.; Chen, C.; Yan, K.-W.; Ponnusamy, M.; Zhang, Y.-H.; et al. Circular RNA mediates cardiomyocyte death via miRNA-dependent upregulation of MTP18 expression. Cell Death Differ. 2017, 24, 1111. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Li, C.; Yue, L.; Ding, N.; Riordan, T.; Yang, L.; Li, Y.; Jen, C.; Lin, S.; et al. Circular RNA profiling provides insights into their subcellular distribution and molecular characteristics in HepG2 cells. Rna Biol. 2019, 16, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Kren, B.T.; Wong, P.Y.-P.; Sarver, A.; Zhang, X.; Zeng, Y.; Steer, C.J. MicroRNAs identified in highly purified liver-derived mitochondria may play a role in apoptosis. Rna Biol 2009, 6, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.; Li, L.-M.; Tang, R.; Hou, D.-X.; Chen, X.; Zhang, C.-Y.; Zen, K. Identification of mouse liver mitochondria-associated miRNAs and their potential biological functions. Cell Res. 2010, 20, 1076. [Google Scholar] [CrossRef] [Green Version]
- Bandiera, S.; Rüberg, S.; Girard, M.; Cagnard, N.; Hanein, S.; Chrétien, D.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. Nuclear outsourcing of RNA interference components to human mitochondria. PLoS ONE 2011, 6, e20746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ro, S.; Ma, H.-Y.; Park, C.; Ortogero, N.; Song, R.; Hennig, G.W.; Zheng, H.; Lin, Y.-M.; Moro, L.; Hsieh, J.-T.; et al. The mitochondrial genome encodes abundant small noncoding RNAs. Cell Res. 2013, 23, 759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrey, E.; Saint-Auret, G.; Bonnamy, B.; Damas, D.; Boyer, O.; Gidrol, X. Pre-microRNA and mature microRNA in human mitochondria. PLoS ONE 2011, 6, e20220. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Ferlito, M.; Kent, O.A.; Fox-Talbot, K.; Wang, R.; Liu, D.; Raghavachari, N.; Yang, Y.; Wheelan, S.J.; Murphy, E.; et al. Nuclear miRNA regulates the mitochondrial genome in the heart. Circ. Res. 2012, 110, 1596–1603. [Google Scholar] [CrossRef] [Green Version]
- Sripada, L.; Tomar, D.; Prajapati, P.; Singh, R.; Singh, A.K.; Singh, R. Systematic analysis of small RNAs associated with human mitochondria by deep sequencing: Detailed analysis of mitochondrial associated miRNA. PLoS ONE 2012, 7, e44873. [Google Scholar] [CrossRef]
- Jagannathan, R.; Thapa, D.; Nichols, C.E.; Shepherd, D.L.; Stricker, J.C.; Croston, T.L.; Baseler, W.A.; Lewis, S.E.; Martinez, I.; Hollander, J.M. Translational regulation of the mitochondrial genome following redistribution of mitochondrial microRNA in the diabetic heart. Circ. Cardiovasc. Genet. 2015, 8, 785–802. [Google Scholar] [CrossRef] [Green Version]
- Kwon, C.; Tak, H.; Rho, M.; Chang, H.R.; Kim, Y.H.; Kim, K.T.; Balch, C.; Lee, E.K.; Nam, S. Detection of PIWI and piRNAs in the mitochondria of mammalian cancer cells. Biochem. Biophys. Res. Commun. 2014, 446, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Richard, J.L.C.; Eichhorn, P.J.A. Deciphering the roles of lncRNAs in breast development and disease. Oncotarget 2018, 9, 20179. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Sunwoo, H.; Spector, D.L. Long noncoding RNAs: Functional surprises from the RNA world. Genes Dev. 2009, 23, 1494–1504. [Google Scholar] [CrossRef] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333. [Google Scholar] [CrossRef]
- Wilusz, J.E. A 360 view of circular RNAs: From biogenesis to functions. Wiley Interdiscip. Rev. Rna 2018, 9, e1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, A.F.; Sandoval, G.T.; Magalhães, L.; Santos, S.E.; Ribeiro-dos-Santos, Â. Circular RNAs as a new field in gene regulation and their implications in translational research. Epigenomics 2016, 8, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Danan, M.; Schwartz, S.; Edelheit, S.; Sorek, R. Transcriptome-wide discovery of circular RNAs in Archaea. Nucleic Acids Res. 2011, 40, 3131–3142. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.; Fiskaa, T.; Birgisdottir, A.B.; Haugen, P.; Einvik, C.; Johansen, S. The ability to form full-length intron RNA circles is a general property of nuclear group I introns. RNA 2003, 9, 1464–1475. [Google Scholar] [CrossRef] [Green Version]
- Mohr, A.M.; Mott, J.L. Overview of MicroRNA Biology. In Seminars in liver disease; Thieme Medical Publishers: New York, NY, USA, 2015; Volume 35, pp. 3–11. [Google Scholar]
- Huang, L.; Mollet, S.; Souquere, S.; Le Roy, F.; Ernoult-Lange, M.; Pierron, G.; Dautry, F.; Weil, D. Mitochondria associate with P-bodies and modulate microRNA-mediated RNA interference. J. Biol. Chem. 2011, 286, 24219–24230. [Google Scholar] [CrossRef] [Green Version]
- Bandiera, S.; Hanein, S.; Lyonnet, S.; Henrion-Caude, A. Mitochondria as novel players of the cellular RNA interference. J. Biol. Chem. 2011, 286, 19. [Google Scholar] [CrossRef] [Green Version]
- V Duarte, F.; A Amorim, J.; M Palmeira, C.; P Rolo, A. Regulation of mitochondrial function and its impact in metabolic stress. Curr. Med. Chem. 2015, 22, 2468–2479. [Google Scholar] [CrossRef]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2018, 1, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Mani, S.R.; Juliano, C.E. Untangling the web: The diverse functions of the PIWI/piRNA pathway. Mol. Reprod. Dev. 2013, 80, 632–664. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Gao, Q.; Peng, X.; Choi, S.-Y.; Sarma, K.; Ren, H.; Morris, A.J.; Frohman, M.A. piRNA-associated germline nuage formation and spermatogenesis require MitoPLD profusogenic mitochondrial-surface lipid signaling. Dev. Cell 2011, 20, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Frohman, M.A. Roles for the lipid-signaling enzyme MitoPLD in mitochondrial dynamics, piRNA biogenesis, and spermatogenesis. Bmb Rep. 2012, 45, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czech, B.; Hannon, G.J. One loop to rule them all: The ping-pong cycle and piRNA-guided silencing. Trends Biochem. Sci. 2016, 41, 324–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.; Liu, J.; Dong, K.; Melnick, A.F.; Latham, K.E.; Chen, C. Mitochondrial membrane-based initial separation of MIWI and MILI functions during pachytene piRNA biogenesis. Nucleic Acids Res. 2018, 47, 2594–2608. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Suzuki, T. A complete landscape of post-transcriptional modifications in mammalian mitochondrial tRNAs. Nucleic Acids Res. 2014, 42, 7346–7357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meseguer, S.; Martínez-Zamora, A.; García-Arumí, E.; Andreu, A.L.; Armengod, M.-E. The ROS-sensitive microRNA-9/9* controls the expression of mitochondrial tRNA-modifying enzymes and is involved in the molecular mechanism of MELAS syndrome. Hum. Mol. Genet. 2014, 24, 167–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, J.A.; Francklyn, C.S.; Robey-Bond, S.M. Transfer RNA and human disease. Front. Genet. 2014, 5, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Zhang, H.; Mei, Y.; Wu, M. Reciprocal regulation of HIF-1α and lincRNA-p21 modulates the Warburg effect. Mol. Cell 2014, 53, 88–100. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ncRNAs in Mitochondria | Type | Genomic Origin | Probable Signaling | Function | Related Diseases | Organism | Reference |
|---|---|---|---|---|---|---|---|
| 1 lncRNA | Chimeric | Mitochondrial | Retrograde | NS | NA | Mus musculus | [48] |
| 3 lncRNAs | Simple | Mitochondrial | Retrograde | NS | Cervical cancer | Homo sapiens | [49] |
| 1 lncRNA | Chimeric | Mitochondrial | Retrograde | NS | NA | Mus musculus | [50] |
| 1 lncRNA | Chimeric | Mitochondrial | Retrograde | NS | NA | Mus musculus and Homo sapiens | [51] |
| 1 lncRNA | Chimeric | Mitochondrial | Retrograde | Cell proliferation | Multiple cancers | Homo sapiens | [52] |
| 2 lncRNAs | Chimeric | Mitochondrial | Retrograde | Cell cycle regulation | Multiple cancers | Homo sapiens | [53] |
| 2 lncRNAs | Chimeric | Mitochondrial | Retrograde | Cell survival | Renal cell carcinoma | Homo sapiens | [54] |
| 2 lncRNAs | Chimeric | Mitochondrial | Retrograde | Apoptosis | Multiple cancers | Homo sapiens | [55] |
| 2 lncRNAs | Chimeric | Mitochondrial | Retrograde | NS | Melanoma | Mus musculus and Homo sapiens | [56] |
| 1 lncRNA | Chimeric | Mitochondrial | Retrograde | Cell cycle regulation | NA | Mus musculus and Homo sapiens | [57] |
| 2 lncRNAs | Chimeric | Mitochondrial | Retrograde | Cell proliferation | HPV infection and cervical cancer | Homo sapiens | [58] |
| 9 lncRNAs | NS | Mitochondrial | Retrograde | NS | Heart failure | Homo sapiens | [59] |
| 1 lncRNAs | NA | Nuclear | Anterograde | Mitochondrial DNA replication | Cervical cancer | Homo sapiens | [60] |
| 1 lncRNA | NA | Nuclear | Anterograde | NS | NA | Mus musculus | [61] |
| 1 lncRNA | NA | Nuclear | Anterograde | Estrogen transduction signaling | Multiple cancers | Homo sapiens | [62] |
| 1 lncRNA | NA | Nuclear | Anterograde | mt-tRNA maturation | Cervical cancer | Homo sapiens | [63] |
| 1 lncRNA | NA | Nuclear | Anterograde | RNA transport | NA | Yeast and mammalian | [64] |
| 1 circRNA | Intronic | Mitochondrial | Retrograde | NS | NA | Saccharomyces cerevisiae | [65] |
| 2 circRNAs | Intronic | Mitochondrial | Retrograde | NS | NA | Saccharomyces cerevisiae | [66] |
| circRNAs* | Intronic | Mitochondrial | Retrograde | NS | NA | Saccharomyces cerevisiae | [67] |
| circRNAs* | Intronic | Mitochondrial | Retrograde | NS | NA | Triticum aestivum | [68] |
| 62 circRNAs | Exonic | Mitochondrial | Retrograde | Micronutrient response | NA | Hordeum vulgare | [69] |
| 3 circRNAs | NS | Mitochondrial | Retrograde | NS | NA | Bos taurus | [70] |
| 1 circRNA | Exonic | Nuclear | Anterograde | Regulation of apoptosis and mitochondrial fission | Cardiovascular diseases | Mus musculus | [71] |
| 118 circRNAs | NS | Mitochondrial | Retrograde | NS | Liver cancer | Homo sapiens | [72] |
| 31 sncRNAs | NA | Mitochondrial | Retrograde | NS | Osteosarcoma | Homo sapiens | [46] |
| 15 miRNAs | NA | Nuclear | Anterograde | Apoptosis, cell proliferation and differentiation | NA | Mus musculus | [73] |
| 40 miRNAs | NA | Nuclear | Anterograde | Apoptosis, cell proliferation and differentiation | NA | Mus musculus | [74] |
| 57 miRNAs | NA | 54 nuclear | Anterograde | Cell cycle, ATP synthesis and mitochondrial translation | Cervical cancer | Homo sapiens | [75] |
| NA | 3 mitochondrial | Retrograde | |||||
| 1499 sncRNAs | NA | Mitochondrial | Retrograde | Control of mitochondrial gene expression | NA | Mus musculus | [76] |
| 2540 sncRNAs | NA | Retrograde | Homo sapiens | ||||
| 25 miRNAs | NA | Mitochondrial | Retrograde | Silencing of mitochondrial mRNAs | NA | Homo sapiens | [77] |
| 46 miRNAs | NA | Nuclear | Anterograde | ||||
| 15 miRNAs | NA | Nuclear | Anterograde | Regulate mitochondrial functions | NA | Mus musculus | [78] |
| 428 miRNAs (HEK293) | NA | Nuclear | Anterograde | Apoptosis, cell cycle and nucleotide metabolism | NA | Homo sapiens | [79] |
| 327 miRNAs (HeLa) | NA | Nuclear | Anterograde | Cervical cancer | |||
| 4 miRNAs (HEK293/HeLa) | NA | Mitochondrial | Retrograde | NA/Cervical cancer | |||
| piRNAs (HEK293/HeLa)* | NA | Mitochondrial | Retrograde | NS | NA/Cervical cancer | ||
| 78 miRNAs | NA | Nuclear | Anterograde | Silencing of mitochondrial mRNAs | Type 1 Diabetes Mellitus | Mus musculus | [80] |
| 29 piRNAs | NA | Mitochondrial | Retrograde | NS | Multiple cancers | Homo sapiens | [81] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalcante, G.C.; Magalhães, L.; Ribeiro-dos-Santos, Â.; Vidal, A.F. Mitochondrial Epigenetics: Non-Coding RNAs as a Novel Layer of Complexity. Int. J. Mol. Sci. 2020, 21, 1838. https://doi.org/10.3390/ijms21051838
Cavalcante GC, Magalhães L, Ribeiro-dos-Santos Â, Vidal AF. Mitochondrial Epigenetics: Non-Coding RNAs as a Novel Layer of Complexity. International Journal of Molecular Sciences. 2020; 21(5):1838. https://doi.org/10.3390/ijms21051838
Chicago/Turabian StyleCavalcante, Giovanna C., Leandro Magalhães, Ândrea Ribeiro-dos-Santos, and Amanda F. Vidal. 2020. "Mitochondrial Epigenetics: Non-Coding RNAs as a Novel Layer of Complexity" International Journal of Molecular Sciences 21, no. 5: 1838. https://doi.org/10.3390/ijms21051838
APA StyleCavalcante, G. C., Magalhães, L., Ribeiro-dos-Santos, Â., & Vidal, A. F. (2020). Mitochondrial Epigenetics: Non-Coding RNAs as a Novel Layer of Complexity. International Journal of Molecular Sciences, 21(5), 1838. https://doi.org/10.3390/ijms21051838