Characterization and Evaluation of Resistance to Powdery Mildew of Wheat–Aegilops geniculata Roth 7Mg (7A) Alien Disomic Substitution Line W16998


Abstract
:1. Introduction
2. Results
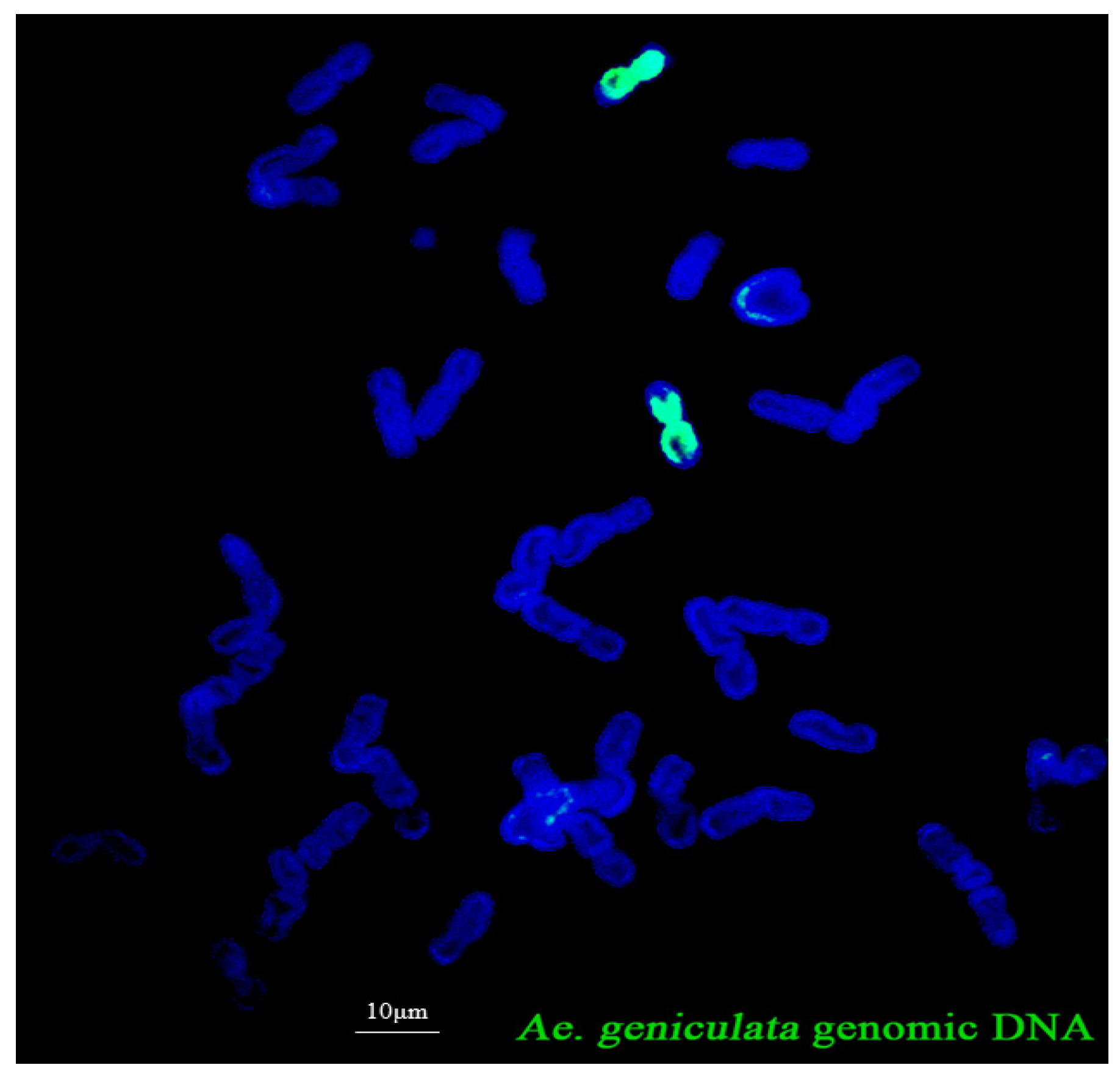
2.1. Cytological Characterization
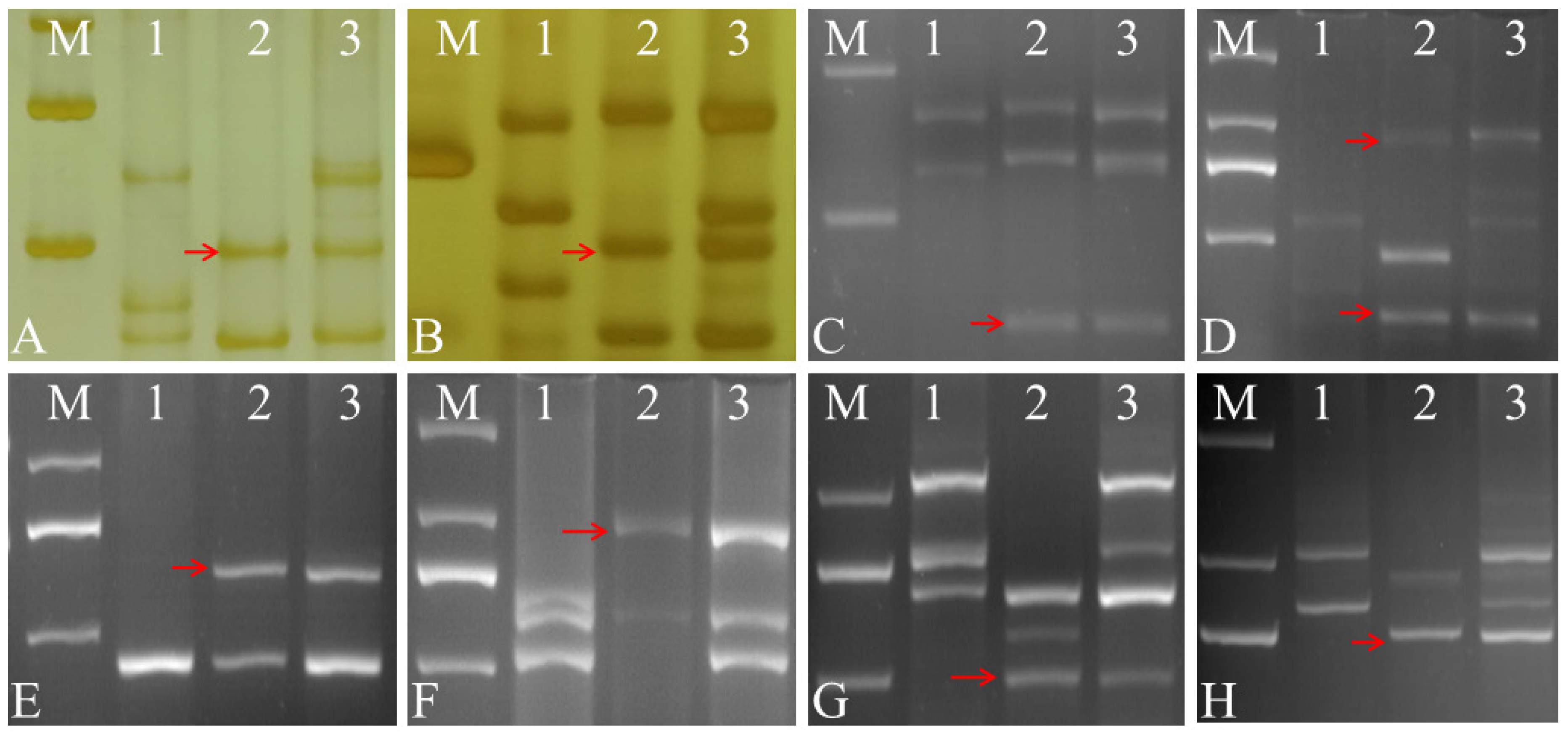
2.2. Molecular Marker Analysis
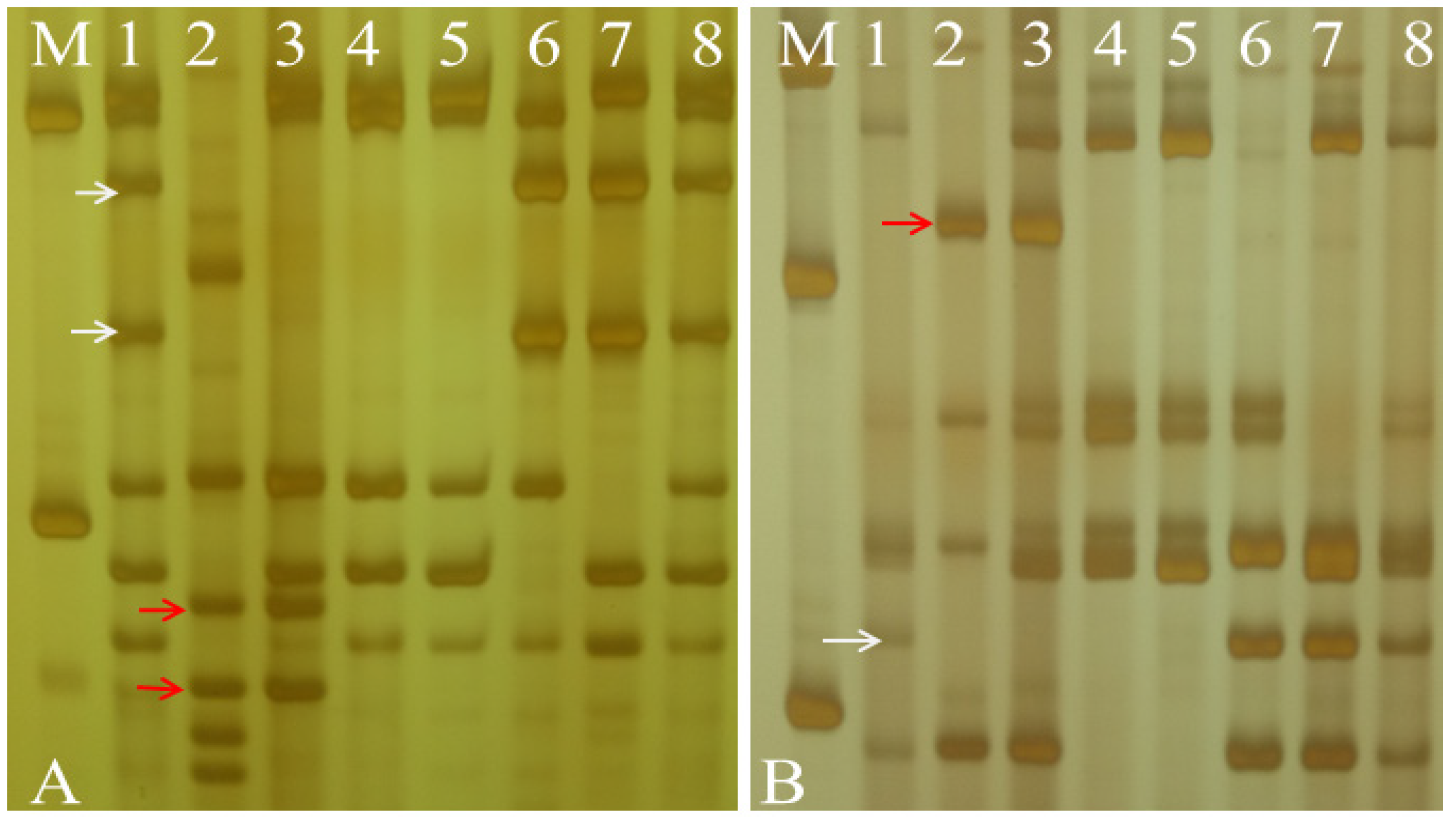
2.3. Nullisomic–Tetrasomic Analysis
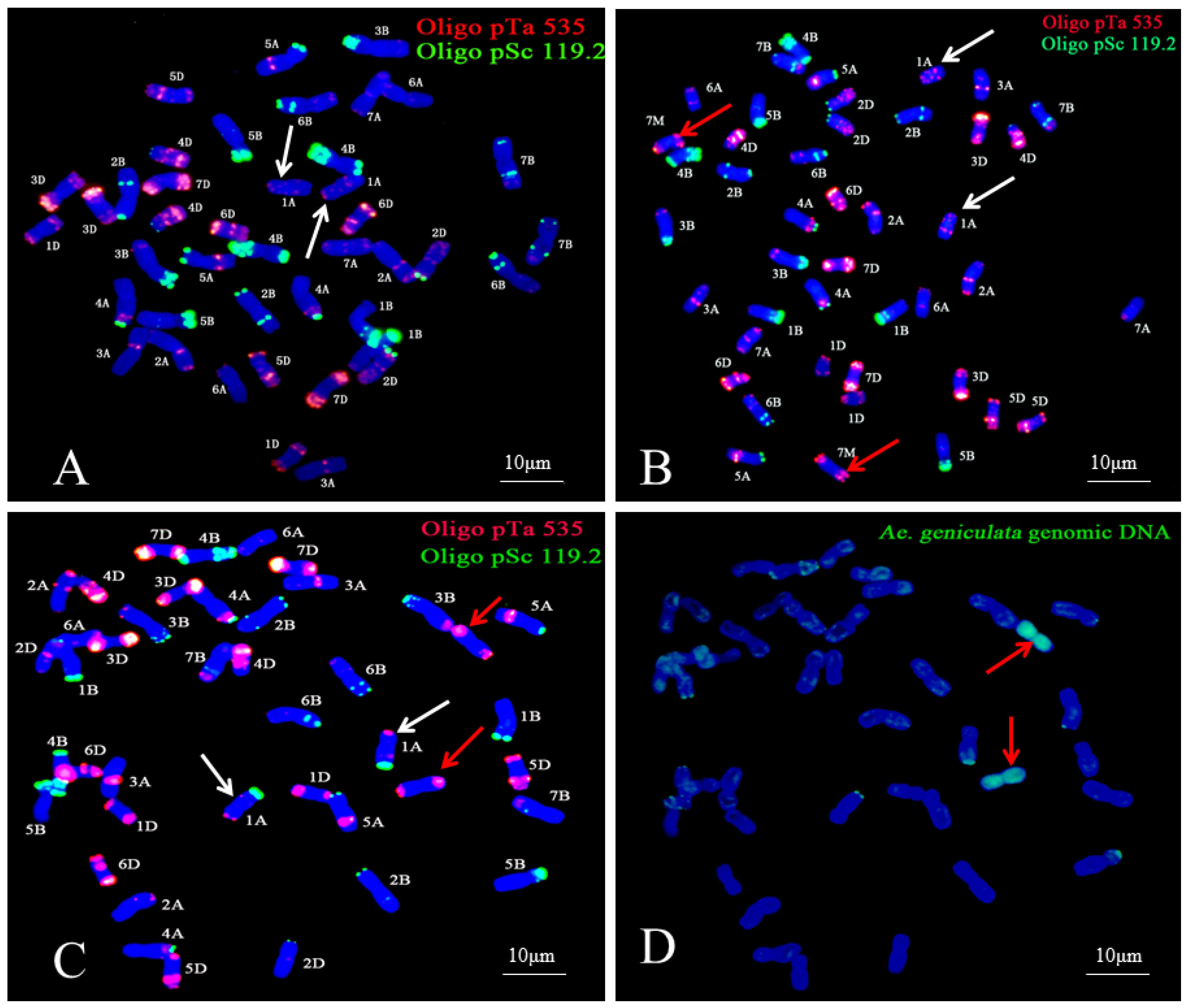
2.4. FISH and Sequential FISH–GISH Analysis
2.5. Evaluation of Powdery Mildew Resistance and Agronomic Traits
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cytological Observation
4.3. GISH, FISH and Sequential FISH–GISH
4.4. EST–STS and PLUG Markers Analysis
4.5. Powdery Mildew Resistance and Agronomic Trait Evaluation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ullah, K.N.; Li, N.; Shen, T.; Wang, P.S.; Tang, W.B.; Ma, S.W.; Zhang, Z.M.; Jia, H.Y.; Kong, Z.X.; Ma, Z.Q. Fine mapping of powdery mildew resistance gene Pm4e in bread wheat (Triticum aestivum L.). Planta 2018, 248, 1319–1328. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Z.Y.; Hu, Y.K.; Duan, X.Y.; Xin, Z.Y. Identification and molecular mapping of a resistance gene to powdery mildew from the synthetic wheat line M53. J. Appl. Genet. 2011, 52, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Benavente, E.; Fernández-Calvín, B.; Orellana, J. Relationship between the levels of wheat-rye metaphase I chromosomal pairing and recombination revealed by GISH. Chromosoma 1996, 105, 92–96. [Google Scholar] [CrossRef]
- Wang, Y.J.; Quan, W.; Peng, N.N.; Wang, C.Y.; Yang, X.F.; Liu, X.L.; Zhang, H.; Chen, C.H.; Ji, W.Q. Molecular cytogenetic identification of a wheat-Aegilops geniculata Roth 7Mg disomic addition line with powdery mildew resistance. Mol. Breed. 2016, 36, 40. [Google Scholar] [CrossRef]
- Zhu, C.; Wang, Y.Z.; Chen, C.H.; Wang, C.Y.; Zhang, A.C.; Peng, N.N.; Wang, Y.J.; Zhang, H.; Liu, X.L.; Ji, W.Q. Molecular cytogenetic identification of a wheat-Thinopyrum ponticum substitution line with stripe rust resistance. Genome 2017, 60, 860–867. [Google Scholar] [CrossRef]
- Chen, P.D.; Qi, L.L.; Zhou, B.; Zhang, S.Z.; Liu, D.J. Development and molecular cytogenetic analysis of wheat—Haynaldia villosa 6VS/6AL translocation lines specifying resistance to powdery mildew. Theoret. Appl. Genet. 1995, 91, 1125–1128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.Q.; Sun, B.X.; Chen, J.; Cao, A.Z.; Xing, L.P.; Feng, L.G.; Lan, C.X.; Chen, P.D. Pm55, a developmental-stage and tissue-specific powdery mildew resistance gene introgressed from Dasypyrum villosum into common wheat. Theor. Appl. Genet. 2016, 10, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.G.; Luo, H.Y.; Chang, Z.J.; Zhang, H.Y.; Zhang, M.; Ren, Z.L. Characterization and chromosomal location of Pm40 in common wheat: A new gene for resistance to powdery mildew derived from Elytrigia intermedium. Theor. Appl. Genet. 2009, 118, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Devos, K.M.; Chao, S.; Miller, T.E.; Reader, S.M.; Gale, M.D. RFLP-based maps of the homoeologous group-6 chromosomes of wheat and their application in the tagging of Pm12, a powdery mildew resistance gene transferred from Aegilops speltoides to wheat. Theor. Appl. Genet. 1996, 92, 559–565. [Google Scholar] [CrossRef]
- Hsam, S.L.K.; Lapochkina, I.F.; Zeller, F.J. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.). Gene Pm32 in a wheat-Aegilops speltoides translocation line. Euphytica 2003, 133, 367–370. [Google Scholar] [CrossRef]
- Petersen, S.; Lyerly, J.H.; Worthington, M.L.; Parks, W.R.; Cowger, C.; Marshall, D.S.; Brown-Guedira, G.; Murphy, J.P. Mapping of powdery mildew resistance gene Pm53 introgressed from Aegilops speltoides into soft red winter wheat. Theor. Appl. Genet. 2015, 128, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Ceoloni, C.; Signore, G.D.; Ercoli, L.; Donini, P. Locating the alien chromatin segment in common wheat-Aegilops longissima mildew resistant transfers. Hereditas 1992, 116, 239–245. [Google Scholar] [CrossRef]
- Liu, W.X.; Koo, D.-H.; Xia, Q.; Li, C.X.; Bai, F.Q.; Song, Y.L.; Friebe, B.; Gill, B.S. Homoeologous recombination-based transfer and molecular cytogenetic mapping of powdery mildew-resistant gene Pm57 from Aegilops searsii into wheat. Theor. Appl. Genet. 2017, 130, 841–848. [Google Scholar] [CrossRef] [PubMed]
- He, R.L.; Chang, Z.J.; Yang, Z.J.; Yuan, Z.Y.; Zhan, H.X.; Zhang, X.J. Inheritance and mapping of powdery mildew resistance gene Pm43 introgressed from Thinopyrum intermedium into wheat. Theor. Appl. Genet. 2009, 118, 1173–1180. [Google Scholar] [CrossRef]
- Zaharieva, M.; Monneveux, P.; Henry, M.; Rivoal, R.; Valkoun, J.; Nachit, M.M. Evaluation of a collection of wild wheat relative Aegilops geniculata Roth and identification of potential sources for useful traits. Euphytica 2001, 119, 33–38. [Google Scholar] [CrossRef]
- Gill, B.S.; Sharma, H.C.; Raupp, W.J.; Browder, L.E.; Hatchett, J.H.; Harvey, T.L.; Moseman, J.G.; Waines, J.W. Evaluation of Aegilops species for resistance to powdery mildew, wheat leaf rust, Hessian fly, and greenbug. Plant Dis. 1985, 69, 314–316. [Google Scholar] [CrossRef]
- Friebe, B.R.; Jiang, J.; Raupp, W.J.; McIntosh, R.A.; Gill, B.S. Characterization of wheat-alien translocations conferring resistance to diseases and pests: Current status. Euphytica 1996, 91, 59–87. [Google Scholar] [CrossRef]
- Ohta, S. Diverse morphological and cytogenetic variation and differentiation of the two subspecies in Aegilops geniculata Roth, a wild relative of wheat. Genet. Resour. Crop Evol. 2017, 64, 2009–2020. [Google Scholar] [CrossRef]
- Rawat, N.; Tiwari, V.K.; Singh, N.; Randhawa, G.S.; Singh, K.; Chhuneja, P.; Dhaliwal, H.S. Evaluation and utilization of Aegilops and wild Triticum species for enhancing iron and zinc content in wheat. Genet. Resour. Crop Evol. 2009, 56, 53–64. [Google Scholar] [CrossRef]
- Siddiqui, K.A.; Yosufzai, M.N. Natural and indused variation for endomorphic traits in the tribe Triticeae. In Proceedings of the 7th International Wheat Genetics Symposium, Cambridge, UK, 13–19 July 1988; pp. 139–144. [Google Scholar]
- Zhang, X.Y.; Wang, R.R.C.; Dong, Y.S. RAPD polymorphisms in Aegilops geniculata Roth (Ae. ovata auct. non L.). Genet. Resour. Crop Evol. 1996, 43, 429–433. [Google Scholar] [CrossRef]
- Friebe, B.R.; Tuleen, N.A.; Gill, B.S. Development and identification of a complete set of Triticum aestivum-Aegilops geniculata chromosome addition lines. Genome 1999, 42, 374–380. [Google Scholar] [CrossRef]
- Kuraparthy, V.; Chhuneja, P.; Dhaliwal, H.S.; Kaur, S.; Bowden, R.L.; Gill, B.S. Characterization and mapping of cryptic alien introgression from Aegilops geniculata with new leaf rust and stripe rust resistance genes Lr57 and Yr40 in wheat. Theor. Appl. Genet. 2007, 114, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Zeller, F.; Kong, L.; Hartl, L.; Mohler, V.; Hsam, S.L.K. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.) Gene Pm29 in line Pova. Euphytica 2002, 123, 187–194. [Google Scholar] [CrossRef]
- Tang, Z.X.; Yang, Z.J.; Fu, S.L. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef]
- Lukaszewski, A.J.; Porter, D.R.; Baker, C.A.; Rybka, K.; Lapinski, B. Attempts to transfer Russian wheat aphid resistance from a rye chromosome in Russian triticales to wheat. Crop Sci. 2001, 41, 1743–1749. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhuang, L.F.; Sun, L.; Feng, Y.G.; Pei, Z.Y.; Qi, Z.J. Allocation of a powdery mildew resistance locus to the chromosome arm 6RL of Secale cereale L. cv. ‘Jingzhouheimai’. Euphytica 2010, 176, 157–166. [Google Scholar] [CrossRef]
- Zhuang, L.F.; Liu, P.; Liu, Z.Q.; Chen, T.T.; Wu, N.; Sun, L.; Qi, Z.J. Multiple structural aberrations and physical mapping of rye chromosome 2R introgressed into wheat. Mol. Breed. 2015, 35, 133. [Google Scholar] [CrossRef]
- Fu, S.L.; Yang, M.Y.; Ren, Z.L.; Yan, B.; Tang, Z.X. Abnormal mitosis induced by wheat-rye 1R monosomic addition lines. Genome 2014, 57, 21–28. [Google Scholar] [CrossRef]
- Fu, S.L.; Chen, L.; Wang, Y.Y.; Li, M.; Yang, Z.J.; Qiu, L.; Yan, B.J.; Ren, Z.L.; Tang, Z.X. Oligonucleotide probes for ND-FISH analysis to identify rye and wheat chromosomes. Sci. Rep. 2015, 215, 10552. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, M.; Sepsi, A.; Tyankova, N.; Molnár-Láng, M. Molecular cytogenetic characterization of two high protein wheat-Thinopyrum intermedium partial amphiploids. J. Appl. Genet. 2011, 52, 269–277. [Google Scholar] [CrossRef]
- Yang, X.F.; Wang, C.Y.; Li, X.; Chen, C.H.; Tian, Z.R.; Wang, Y.J.; Ji, W.Q. Development and Molecular Cytogenetic Identification of a Novel Wheat-Leymus mollis Lm#7Ns (7D) Disomic Substitution Line with Stripe Rust Resistance. PLoS ONE 2015, 10, e0140227. [Google Scholar] [CrossRef]
- Hu, L.J.; Li, G.R.; Zeng, Z.X.; Chang, Z.J.; Liu, C.; Yang, Z.J. Molecular characterization of a wheat-Thinopyrum ponticum partial amphiploid and its derived substitution line for resistance to stripe rust. J. Appl. Genet. 2011, 52, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guo, X.X.; Wang, C.Y.; Ji, W.Q. Spontaneous and divergent hexaploid Triticales derived from common wheat × Rye by complete elimination of D-genome chromosomes. PLoS ONE 2015, 10, e0120421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Lv, Z.L.; Niu, Z.X.; Li, B.; Li, H.W.; Xu, S.S.; Han, F.P.; Li, Z.S. Molecular cytogenetic characterization and stem rust resistance of five wheat-Thinopyrum ponticum partial amphiploids. J. Genet. Genom. 2014, 41, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Linc, G.; Molnár-Láng, M. Fluorescence in situ hybridization polymorphism using two repetitive DNA clones in different cultivars of wheat. Plant Breed. 2003, 122, 396–400. [Google Scholar] [CrossRef]
- Kwiatek, M.; Wiśniewska, H.; Apolinarska, B. Cytogenetic analysis of Aegilops chromosomes, potentially usable in triticale (X Triticosecale Witt.) breeding. J. Appl. Genet. 2013, 54, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.T.; Lei, Y.H.; Zhang, H.T.; Song, D.H.; Liu, X.; Cao, Z.L.; Chu, C.G.; Zhuang, L.F.; Qi, Z.J. Frequent variations in tandem repeats pSc200 and pSc119.2 cause rapid chromosome evolution of open-pollinated rye. Mol. Breed. 2019, 39, 133. [Google Scholar] [CrossRef]
- Frederic, J.R.; Bauer, P.J. Ecology and Physiology of Yield Determination; Food Products Press: Binghamtom, NY, USA, 2000; pp. 45–65. [Google Scholar]
- Yen, C.; Zheng, Y.L.; Yang, J.L. An ideotype for high yield breeding in theory and practice. In Proceedings of the 8th International Wheat Genetics Symposium, Beijing, China, 20–25 July 1993; Agricultural Sci-Tech. Press: Beijing, China, 1995; pp. 1113–1117. [Google Scholar]
- Wu, J.; Yang, X.M.; Wang, H.; Li, H.J.; Li, L.H.; Li, X.Q.; Liu, W.H. The introgression of chromosome 6P specifying for increased numbers of florets and kernels from Agropyron cristatum into wheat. Theor. Appl. Genet. 2006, 114, 13–20. [Google Scholar] [CrossRef]
- Li, Z.S.; Rong, S.; Cheng, S.Y.; Zhong, G.C.; Mu, S.M. Wheat Wide Hybridization; Chinese Scientific Press: Beijing, China, 1985. [Google Scholar]
- An, D.G.; Ma, P.T.; Zheng, Q.; Fu, S.L.; Li, L.H.; Han, F.P.; Han, G.H.; Wang, J.; Xu, Y.F.; Jin, Y.L.; et al. Development and molecular cytogenetic identification of a new wheat-rye 4R chromosome disomic addition line with resistances to powdery mildew, stripe rust and sharp eyespot. Theor. Appl. Genet. 2019, 132, 257–272. [Google Scholar] [CrossRef]
- Han, F.P.; Lamb, J.C.; Birchler, J.A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef] [Green Version]
- Pei, Y.R.; Cui, Y.; Zhang, Y.P.; Wang, H.G.; Bao, Y.G.; Li, X.F. Molecular cytogenetic identification of three rust-resistant wheat-Thinopyrum ponticum partial amphiploids. Mol. Cytogenet. 2018, 11, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, G.; Nakamura, T.; Ashida, T.; Saito, M.; Nasuda, S.; Endo, T.R.; Wu, J.Z.; Matsumoto, T. Localization of anchor loci representing five hundred annotated rice genes to wheat chromosomes using PLUG markers. Theor. Appl. Genet. 2009, 118, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Sheng, B. Grades of resistance to powdery mildew classified by different phenotypes of response in the seeding stage of wheat. Plant Prot. 1988, 1, 49. [Google Scholar]
- Wang, Y.J.; Wang, C.Y.; Quan, W.; Jia, X.J.; Fu, Y.; Zhang, H.; Liu, X.L.; Chen, C.H.; Ji, W.Q. Identification and mapping of PmSE5785, a new recessive powdery mildew resistance locus, in synthetic hexaploid wheat. Euphytica 2016, 207, 619–626. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Type | Primer (5’-3’) | Location | Geltype/Restrictionenzyme | Tm °C/t (h) |
|---|---|---|---|---|---|
| BE637663 | EST-SSR | F: ACTGTTGCTTCGCTCCAAGT R: GTTCCATTTCCGATGTGCTC | 7AL 7BL 7DL | 8% non-denaturing polyacrylamide gel/- | 60/- |
| TNAC1868 | PLUG | F: CTCCGCCTTCATCGGAAA R: CCGTTCTGCTTCAGGATCTC | 7AL 7BL 7DL | 8% non-denaturing polyacrylamide gel/- | 60/- |
| TNAC1782 | PLUG | F: TCACTGAACAGCCTAGACATGG R: ATTCGCAGACCGCATCTATC | 7AS 7BS 7DS | 2% agarose gel/TaqI/HaeIII | 60/2 or 37/2 |
| TNAC1829 | PLUG | F: GCCACTTCCTCCCTCCTC R: GTCGGTCCTCCAGTATCAGC | 7AL 7BL 7DL | 2% agarose gel/TaqI | 60/2 |
| TNAC1845 | PLUG | F: AATGAACAGCTTGCTTTCTGC R: CAGATGCTCTGGATTTCATGG | 7AL 7BL 7DL | 2% agarose gel/TaqI | 60/2 |
| TNAC1888 | PLUG | F: AGGGATGTGTTGGAGCTGTTA R: CACAGTGACCTTCTGCTCCTT | 7AL 7BL 7DL | 2% agarose gel/TaqI | 60/2 |
| TNAC1929 | PLUG | F: GCACCAGAAGGTTCAGTAGCA R: ATCTGTCAGCAGGGCACACT | 7AS 7BS 7DS | 2% agarose gel/TaqI | 60/2 |
| TNAC1941 | PLUG | F: AATGATCCTGACAAGGTGCAG R: GTAGCGATGGCATCCAGAGA | 7AS 7BS 7DS | 2% agarose gel/TaqI | 60/2 |
| Materials | Tillering | Plant Height (cm) | Spike Length (cm) | Spikelets/Spike | Kernels/Spikelet | Kernels/Spike | Thousand Kenel Weight (g) | Awnedness |
|---|---|---|---|---|---|---|---|---|
| CS | 13 ± 4 | 130 ± 5 | 9.0 ± 0.3 | 20 ± 2 | 4 ± 1 | 40 ± 4 | 30 ± 0.5 | awnless |
| SY159 | 75 ± 5 | 65 ± 5 | 2.8 ± 0.4 | 3 ± 1 | 3 ± 1 | 10 ± 2 | 18 ± 2.0 | Long awn |
| W16998 | 15 ± 5 | 120 ± 5 | 11.0 ± 0.5 ** | 23 ± 2 | 5 ± 2 | 50 ± 5 ** | 40 ± 1.0 ** | Long awn |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Long, D.; Wang, Y.; Wang, C.; Liu, X.; Zhang, H.; Tian, Z.; Chen, C.; Ji, W. Characterization and Evaluation of Resistance to Powdery Mildew of Wheat–Aegilops geniculata Roth 7Mg (7A) Alien Disomic Substitution Line W16998. Int. J. Mol. Sci. 2020, 21, 1861. https://doi.org/10.3390/ijms21051861
Wang Y, Long D, Wang Y, Wang C, Liu X, Zhang H, Tian Z, Chen C, Ji W. Characterization and Evaluation of Resistance to Powdery Mildew of Wheat–Aegilops geniculata Roth 7Mg (7A) Alien Disomic Substitution Line W16998. International Journal of Molecular Sciences. 2020; 21(5):1861. https://doi.org/10.3390/ijms21051861
Chicago/Turabian StyleWang, Yajuan, Deyu Long, Yanzhen Wang, Changyou Wang, Xinlun Liu, Hong Zhang, Zengrong Tian, Chunhuan Chen, and Wanquan Ji. 2020. "Characterization and Evaluation of Resistance to Powdery Mildew of Wheat–Aegilops geniculata Roth 7Mg (7A) Alien Disomic Substitution Line W16998" International Journal of Molecular Sciences 21, no. 5: 1861. https://doi.org/10.3390/ijms21051861
APA StyleWang, Y., Long, D., Wang, Y., Wang, C., Liu, X., Zhang, H., Tian, Z., Chen, C., & Ji, W. (2020). Characterization and Evaluation of Resistance to Powdery Mildew of Wheat–Aegilops geniculata Roth 7Mg (7A) Alien Disomic Substitution Line W16998. International Journal of Molecular Sciences, 21(5), 1861. https://doi.org/10.3390/ijms21051861