Assessment of the Effect of Sorafenib on Omega-6 and Omega-3 Epoxyeicosanoid Formation in Patients with Hepatocellular Carcinoma

 ,
,
Abstract
:1. Introduction
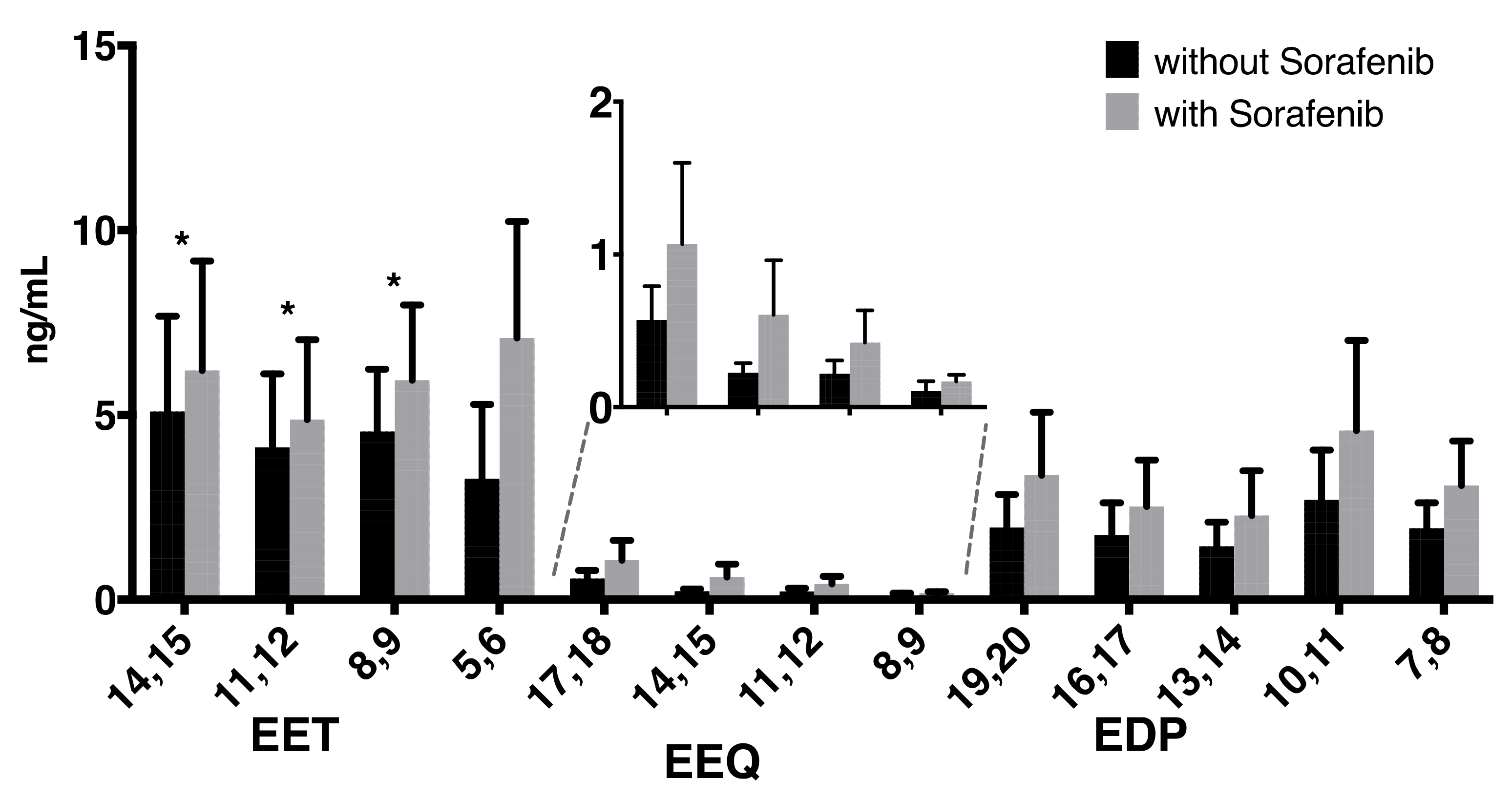
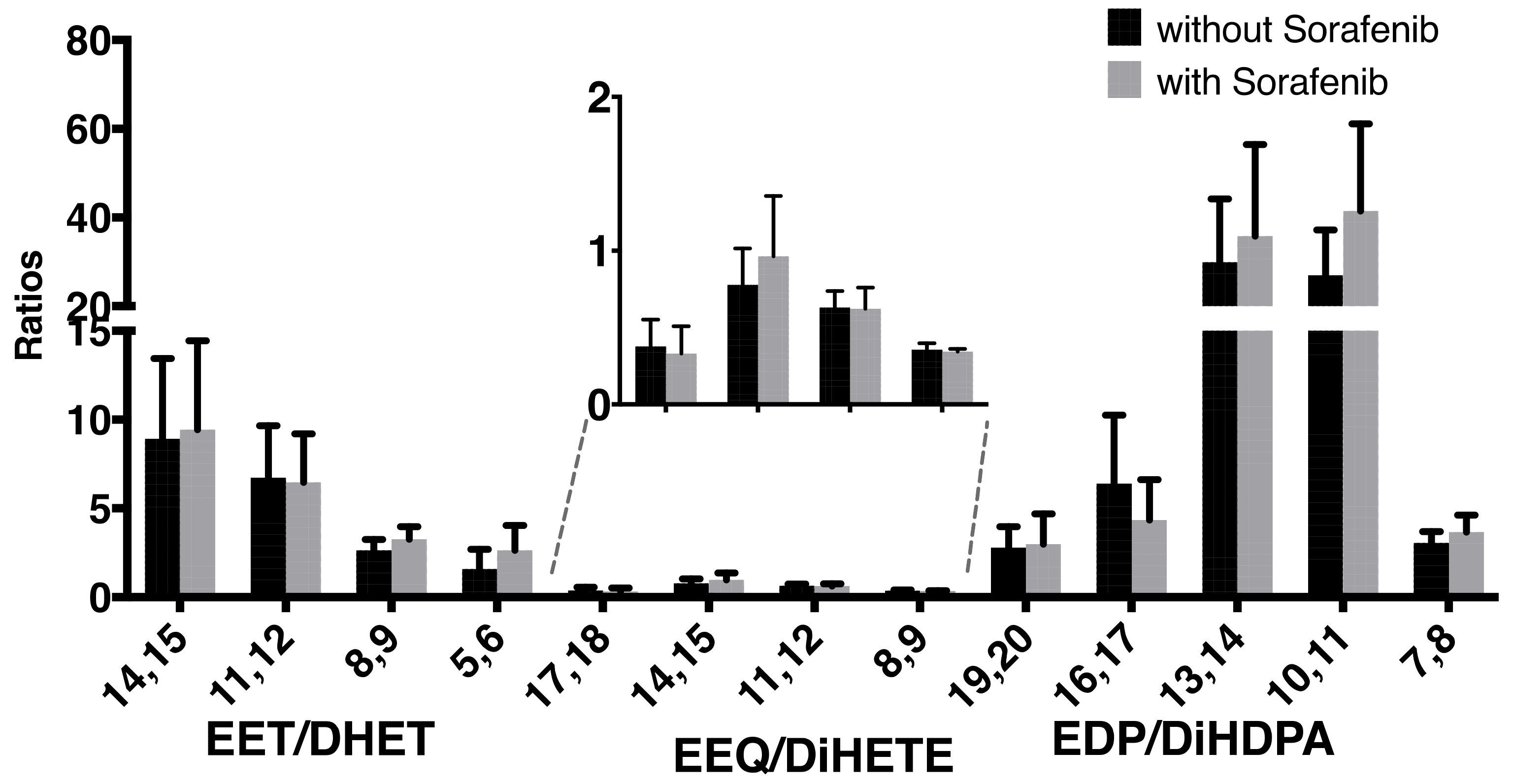
2. Results
3. Discussion
4. Materials and Methods
4.1. Patients and Blood Sampling
4.2. Sample Preparation and LC/ESI-MS/MS
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | arachidonic acid | |
| BHT | butylated hydroxytoluene | |
| CYP450 | cytochrome P450 | |
| DHA | docosahexaenoic acid | |
| DHET | dihydroxyeicosatrienoic acid | |
| DiHDPA | dihydroxydocosapentaenoic acid | |
| DiHETE | dihydroxyeicosatetraenoic acid | |
| EDP | epoxydocosapentaenoic acid | |
| EEQ | epoxyeicosatetraenoic acid | |
| EET | epoxyeicosatrienoic acid | |
| EGF | epidermal growth factor | |
| EPA | eicosapentaenoic acid | |
| FGF | fibroblast growth factor | |
| HCC | hepatocellular carcinoma | |
| LC-PUFA | long-chain polyunsaturated fatty acid | |
| n-3 PUFA | omega-3 polyunsaturated fatty acid | |
| NAFLD | non-alcoholic fatty liver disease | |
| NASH | non-alcoholic steatohepatitis | |
| PDGFR-ß | platelet-derived growth factor receptor ß | |
| sEH | soluble epoxide hydrolase | |
| SPE | solid phase extraction | |
| VEGF | vascular endothelial growth factor |
References
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet (Lond. Engl.) 2018, 391, 1301–1314. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Serag, H.B. Epidemiology of viral hepatitis and hepatocellular carcinoma. Gastroenterology 2012, 142, 1264–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, T.R.; Mandayam, S.; Jamal, M.M. Alcohol and hepatocellular carcinoma. Gastroenterology 2004, 127, S87–S96. [Google Scholar] [CrossRef] [PubMed]
- Sherman, M. Surveillance for hepatocellular carcinoma. Best Pract. Res. Clin. Gastroenterol. 2014, 28, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet (Lond. Engl.) 2012, 379, 1245–1255. [Google Scholar] [CrossRef]
- Wilhelm, S.M.; Carter, C.; Tang, L.; Wilkie, D.; McNabola, A.; Rong, H.; Chen, C.; Zhang, X.; Vincent, P.; McHugh, M.; et al. Bay 43-9006 exhibits broad spectrum oral antitumor activity and targets the raf/mek/erk pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004, 64, 7099–7109. [Google Scholar] [CrossRef] [Green Version]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Mody, K.; Abou-Alfa, G.K. Systemic therapy for advanced hepatocellular carcinoma in an evolving landscape. Curr. Treat. Options Oncol. 2019, 20, 3. [Google Scholar] [CrossRef]
- Zschabitz, S.; Grullich, C. Lenvantinib: A tyrosine kinase inhibitor of vegfr 1-3, fgfr 1-4, pdgfralpha, kit and ret. Recent Results Cancer Res 2018, 211, 187–198. [Google Scholar]
- Harding, J.J. Immune checkpoint blockade in advanced hepatocellular carcinoma: An update and critical review of ongoing clinical trials. Future Oncol. (Lond. Engl.) 2018, 14, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, K.T.; Lathia, C.; Frye, R.F.; Schuchter, L.; Redlinger, M.; Rosen, M.; O’Dwyer, P.J. Interaction of sorafenib and cytochrome p450 isoenzymes in patients with advanced melanoma: A phase i/ii pharmacokinetic interaction study. Cancer Chemother. Pharmacol. 2011, 68, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Park, S.H.; Morisseau, C.; Hwang, S.H.; Hammock, B.D.; Weiss, R.H. Sorafenib has soluble epoxide hydrolase inhibitory activity, which contributes to its effect profile in vivo. Mol. Cancer Ther. 2009, 8, 2193–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Kodani, S.; Hammock, B.D. Stabilized epoxygenated fatty acids regulate inflammation, pain, angiogenesis and cancer. Prog. Lipid Res. 2014, 53, 108–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Yang, J.; Zhang, J.; Wang, Y.; Hwang, S.H.; Qi, W.; Wan, D.; Kim, D.; Sun, J.; Sanidad, K.Z.; et al. Lipidomic profiling reveals soluble epoxide hydrolase as a therapeutic target of obesity-induced colonic inflammation. Proc. Natl. Acad. Sci. USA 2018, 115, 5283–5288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in our understanding of oxylipins derived from dietary pufas. Adv. Nutr. (Bethesda Md.) 2015, 6, 513–540. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases. Nat. Rev. Drug Discov. 2009, 8, 794–805. [Google Scholar] [CrossRef] [Green Version]
- Iyer, A.; Kauter, K.; Alam, M.A.; Hwang, S.H.; Morisseau, C.; Hammock, B.D.; Brown, L. Pharmacological inhibition of soluble epoxide hydrolase ameliorates diet-induced metabolic syndrome in rats. Exp. Diabetes Res. 2012, 2012, 758614. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.H.; Zheng, L.; Gui, L.; Lin, J.Y.; Zhu, Y.M.; Deng, W.S.; Luo, M. Soluble epoxide hydrolase inhibition with t-tucb alleviates liver fibrosis and portal pressure in carbon tetrachloride-induced cirrhosis in rats. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 118–125. [Google Scholar] [CrossRef]
- Yao, L.; Cao, B.; Cheng, Q.; Cai, W.; Ye, C.; Liang, J.; Liu, W.; Tan, L.; Yan, M.; Li, B.; et al. Inhibition of soluble epoxide hydrolase ameliorates hyperhomocysteinemia-induced hepatic steatosis by enhancing beta-oxidation of fatty acid in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G527–G538. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Wang, H.; Xiao, F.; Ning, Q. Epoxyeicosatrienoic acids alleviate methionine-choline-deficient diet-induced non-alcoholic steatohepatitis in mice. Scand. J. Immunol. 2019, 90, e12791. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vicario, C.; Alcaraz-Quiles, J.; Garcia-Alonso, V.; Rius, B.; Hwang, S.H.; Titos, E.; Lopategi, A.; Hammock, B.D.; Arroyo, V.; Claria, J. Inhibition of soluble epoxide hydrolase modulates inflammation and autophagy in obese adipose tissue and liver: Role for omega-3 epoxides. Proc. Natl. Acad. Sci. USA 2015, 112, 536–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panigrahy, D.; Edin, M.L.; Lee, C.R.; Huang, S.; Bielenberg, D.R.; Butterfield, C.E.; Barnes, C.M.; Mammoto, A.; Mammoto, T.; Luria, A.; et al. Epoxyeicosanoids stimulate multiorgan metastasis and tumor dormancy escape in mice. J. Clin. Investig. 2012, 122, 178–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Panigrahy, D.; Mahakian, L.M.; Yang, J.; Liu, J.Y.; Stephen Lee, K.S.; Wettersten, H.I.; Ulu, A.; Hu, X.; Tam, S.; et al. Epoxy metabolites of docosahexaenoic acid (dha) inhibit angiogenesis, tumor growth, and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 6530–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filppula, A.M.; Neuvonen, P.J.; Backman, J.T. In vitro assessment of time-dependent inhibitory effects on cyp2c8 and cyp3a activity by fourteen protein kinase inhibitors. Drug Metab. Dispos. 2014, 42, 1202–1209. [Google Scholar] [CrossRef] [Green Version]
- Morisseau, C.; Hammock, B.D. Impact of soluble epoxide hydrolase and epoxyeicosanoids on human health. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 37–58. [Google Scholar] [CrossRef] [Green Version]
- Node, K.; Huo, Y.; Ruan, X.; Yang, B.; Spiecker, M.; Ley, K.; Zeldin, D.C.; Liao, J.K. Anti-inflammatory properties of cytochrome p450 epoxygenase-derived eicosanoids. Science (N. Y.) 1999, 285, 1276–1279. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.; Chen, S.; You, B.; Sun, J. Activation of sphingosine kinase-1 mediates induction of endothelial cell proliferation and angiogenesis by epoxyeicosatrienoic acids. Cardiovasc. Res. 2008, 78, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Pozzi, A.; Macias-Perez, I.; Abair, T.; Wei, S.; Su, Y.; Zent, R.; Falck, J.R.; Capdevila, J.H. Characterization of 5,6- and 8,9-epoxyeicosatrienoic acids (5,6- and 8,9-eet) as potent in vivo angiogenic lipids. J. Biol. Chem. 2005, 280, 27138–27146. [Google Scholar] [CrossRef] [Green Version]
- Arnold, C.; Konkel, A.; Fischer, R.; Schunck, W.H. Cytochrome p450-dependent metabolism of omega-6 and omega-3 long-chain polyunsaturated fatty acids. Pharmacol. Rep. 2010, 62, 536–547. [Google Scholar] [CrossRef]
- Arnold, C.; Markovic, M.; Blossey, K.; Wallukat, G.; Fischer, R.; Dechend, R.; Konkel, A.; von Schacky, C.; Luft, F.C.; Muller, D.N.; et al. Arachidonic acid-metabolizing cytochrome p450 enzymes are targets of {omega}-3 fatty acids. J. Biol. Chem. 2010, 285, 32720–32733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Aspelund, A.; Alitalo, K. Lymphangiogenic factors, mechanisms, and applications. J. Clin. Investig. 2014, 124, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Nithipatikom, K.; Brody, D.M.; Tang, A.T.; Manthati, V.L.; Falck, J.R.; Williams, C.L.; Campbell, W.B. Inhibition of carcinoma cell motility by epoxyeicosatrienoic acid (eet) antagonists. Cancer Sci. 2010, 101, 2629–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, R.; Sun, L.; Liao, J.; Li, H.; You, X.; Xu, D.; Yang, J.; Hwang, S.H.; Jones, R.D.; Hammock, B.; et al. Inhibition of pancreatic carcinoma growth through enhancing omega-3 epoxy polyunsaturated fatty acid profile by inhibition of soluble epoxide hydrolase. Anticancer Res. 2019, 39, 3651–3660. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profile in man primarily via the cyp-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffitts, J.; Saunders, D.; Tesiram, Y.A.; Reid, G.E.; Salih, A.; Liu, S.; Lydic, T.A.; Busik, J.V.; Kang, J.X.; Towner, R.A. Non-mammalian fat-1 gene prevents neoplasia when introduced to a mouse hepatocarcinogenesis model: Omega-3 fatty acids prevent liver neoplasia. Biochim. Et Biophys. Acta 2010, 1801, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.; Han, C.; Dai, Y.; Shen, M.; Wu, T. Omega-3 polyunsaturated fatty acids inhibit hepatocellular carcinoma cell growth through blocking beta-catenin and cyclooxygenase-2. Mol. Cancer Ther. 2009, 8, 3046–3055. [Google Scholar] [CrossRef] [Green Version]
- Weylandt, K.H.; Krause, L.F.; Gomolka, B.; Chiu, C.Y.; Bilal, S.; Nadolny, A.; Waechter, S.F.; Fischer, A.; Rothe, M.; Kang, J.X. Suppressed liver tumorigenesis in fat-1 mice with elevated omega-3 fatty acids is associated with increased omega-3 derived lipid mediators and reduced tnf-alpha. Carcinogenesis 2011, 32, 897–903. [Google Scholar] [CrossRef] [Green Version]
- Inoue-Yamauchi, A.; Itagaki, H.; Oda, H. Eicosapentaenoic acid attenuates obesity-related hepatocellular carcinogenesis. Carcinogenesis 2018, 39, 28–35. [Google Scholar] [CrossRef]
- Gao, M.; Sun, K.; Guo, M.; Gao, H.; Liu, K.; Yang, C.; Li, S.; Liu, N. Fish consumption and n-3 polyunsaturated fatty acids, and risk of hepatocellular carcinoma: Systematic review and meta-analysis. Cancer Causes Control 2015, 26, 367–376. [Google Scholar] [CrossRef]
- Koh, W.P.; Dan, Y.Y.; Goh, G.B.; Jin, A.; Wang, R.; Yuan, J.M. Dietary fatty acids and risk of hepatocellular carcinoma in the singapore chinese health study. Liver Int. Off. J. Int. Assoc. Study Liver 2016, 36, 893–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, N.; Inoue, M.; Iwasaki, M.; Sasazuki, S.; Shimazu, T.; Yamaji, T.; Takachi, R.; Tanaka, Y.; Mizokami, M.; Tsugane, S. Consumption of n-3 fatty acids and fish reduces risk of hepatocellular carcinoma. Gastroenterology 2012, 142, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Reynolds, L.; Mulik, R.S.; Kim, S.Y.; Van Treuren, T.; Nguyen, L.H.; Zhu, H.; Corbin, I.R. Hepatic arterial infusion of low-density lipoprotein docosahexaenoic acid nanoparticles selectively disrupts redox balance in hepatoma cells and reduces growth of orthotopic liver tumors in rats. Gastroenterology 2016, 150, 488–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Patient | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Age | 64 | 71 | 48 | 67 | 61 | 73 |
| m/f | m | f | m | m | m | m |
| HCC treatments before sorafenib administration | ||||||
| Afterloading | + | - | - | - | - | - |
| Surgical resection | + | - | + | + | - | - |
| Transplantation | - | - | - | - | - | - |
| Transarterial chemoembolization | - | + | - | - | - | - |
| Underlying liver disease | ||||||
| Alcoholic liver disease | - | - | - | - | + | - |
| Hepatitis C | + | + | - | - | - | - |
| Hepatitis B | - | - | + | - | - | - |
| NAFLD/NASH | - | - | - | + | - | + |
| Without Sorafenib | With Sorafenib | ||||
|---|---|---|---|---|---|
| ng/ml | SEM | ng/ml | SEM | ||
| 14,15-EET | 5.09 | 2.58 | 6.21 | 2.96 | * |
| 11,12-EET | 4.11 | 2.00 | 4.87 | 2.17 | * |
| 8,9-EET | 4.55 | 1.68 | 5.94 | 2.04 | * |
| 5,6-EET | 3.28 | 1.83 | 7.08 | 3.15 | |
| 14,15-DHET | 0.53 | 0.06 | 0.66 | 0.12 | |
| 11,12-DHET | 0.53 | 0.05 | 0.73 | 0.23 | |
| 8,9-DHET | 1.51 | 0.26 | 1.71 | 0.37 | |
| 5,6-DHET | 2.51 | 0.56 | 2.96 | 0.71 | |
| 17,18-EEQ | 0.57 | 0.22 | 1.07 | 0.53 | |
| 14,15-EEQ | 0.23 | 0.06 | 0.61 | 0.36 | |
| 11,12-EEQ | 0.22 | 0.08 | 0.42 | 0.21 | |
| 8,9-EEQ | 0.11 | 0.05 | 0.17 | 0.03 | |
| 17,18-DiHETE | 1.66 | 0.21 | 3.32 | 0.61 | * |
| 14,15-DiHETE | 0.30 | 0.06 | 0.54 | 0.11 | |
| 11,12-DiHETE | 0.18 | 0.04 | 0.71 | 0.23 | |
| 8,9-DiHETE | 0.66 | 0.18 | 1.39 | 0.62 | |
| 19,20-EDP | 1.95 | 0.90 | 3.37 | 1.71 | |
| 16,17-EDP | 1.75 | 0.87 | 2.52 | 1.26 | |
| 13,14-EDP | 1.44 | 0.65 | 2.28 | 1.21 | |
| 10,11-EDP | 2.70 | 1.35 | 4.57 | 2.44 | |
| 7,8-EDP | 1.93 | 0.68 | 3.09 | 1.21 | |
| 19,20-DiHDPA | 0.66 | 0.09 | 1.28 | 0.27 | |
| 16,17-DiHDPA | 0.33 | 0.03 | 0.68 | 0.16 | |
| 13,14-DiHDPA | 0.05 | 0.00 | 0.07 | 0.02 | |
| 10,11-DiHDPA | 0.09 | 0.01 | 0.09 | 0.02 | |
| 7,8-DiHDPA | 0.62 | 0.14 | 0.76 | 0.14 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leineweber, C.G.; Pietzner, A.; Zhang, I.W.; Blessin, U.B.; Rothe, M.; Schott, E.; Schebb, N.H.; Weylandt, K.H. Assessment of the Effect of Sorafenib on Omega-6 and Omega-3 Epoxyeicosanoid Formation in Patients with Hepatocellular Carcinoma. Int. J. Mol. Sci. 2020, 21, 1875. https://doi.org/10.3390/ijms21051875
Leineweber CG, Pietzner A, Zhang IW, Blessin UB, Rothe M, Schott E, Schebb NH, Weylandt KH. Assessment of the Effect of Sorafenib on Omega-6 and Omega-3 Epoxyeicosanoid Formation in Patients with Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2020; 21(5):1875. https://doi.org/10.3390/ijms21051875
Chicago/Turabian StyleLeineweber, Can G., Anne Pietzner, Ingrid W. Zhang, Usha B. Blessin, Michael Rothe, Eckart Schott, Nils H. Schebb, and Karsten H. Weylandt. 2020. "Assessment of the Effect of Sorafenib on Omega-6 and Omega-3 Epoxyeicosanoid Formation in Patients with Hepatocellular Carcinoma" International Journal of Molecular Sciences 21, no. 5: 1875. https://doi.org/10.3390/ijms21051875
APA StyleLeineweber, C. G., Pietzner, A., Zhang, I. W., Blessin, U. B., Rothe, M., Schott, E., Schebb, N. H., & Weylandt, K. H. (2020). Assessment of the Effect of Sorafenib on Omega-6 and Omega-3 Epoxyeicosanoid Formation in Patients with Hepatocellular Carcinoma. International Journal of Molecular Sciences, 21(5), 1875. https://doi.org/10.3390/ijms21051875