Exposure to Dichlorodiphenyldichloroethylene (DDE) and Metallothionein Levels in Rats Fed with Normocaloric or High-Fat Diet: A Review



Abstract
:1. Multifunctional Roles of Metallothioneins
2. Metallothioneins and DDT Metabolites
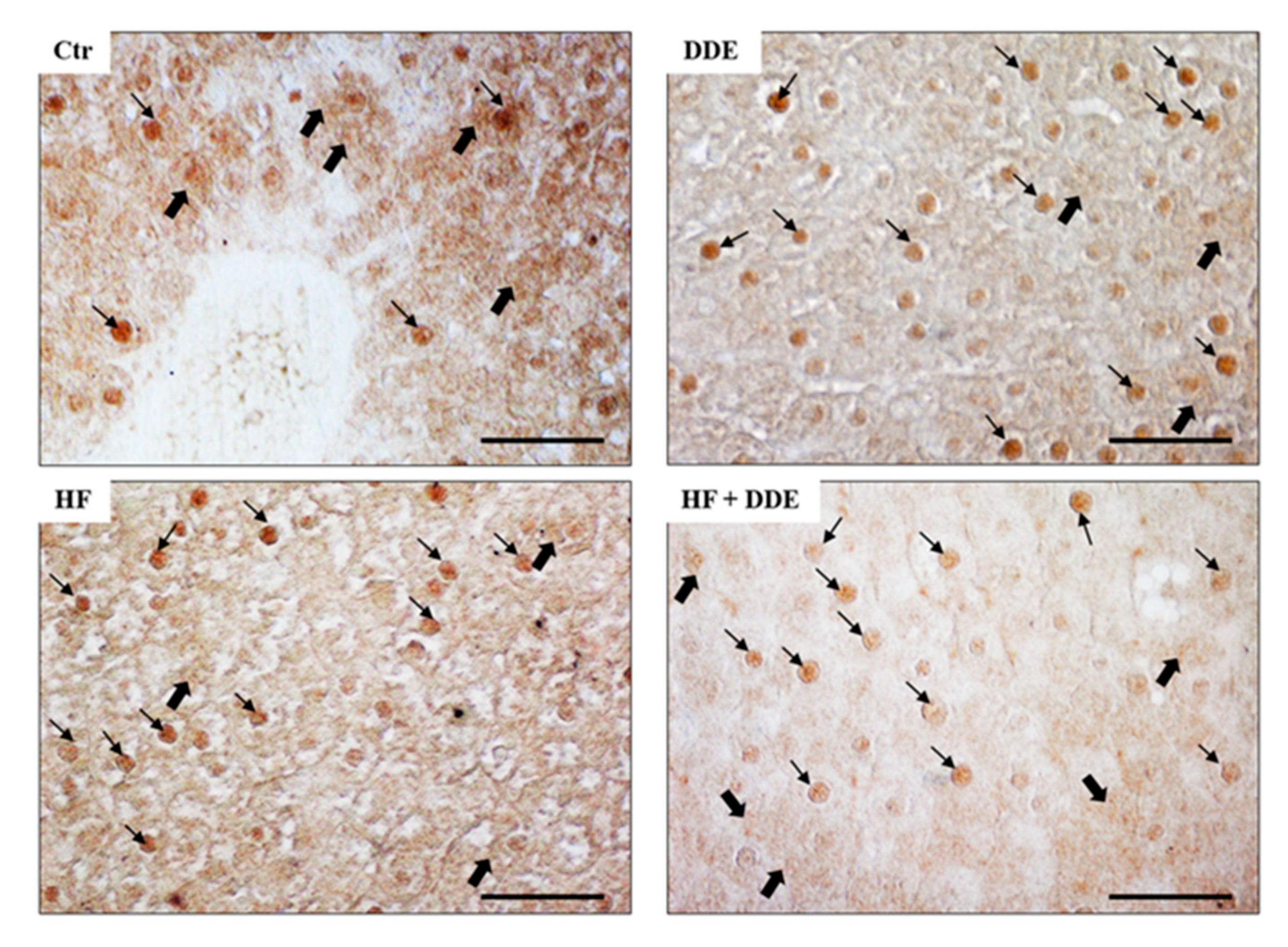
3. Effect of DDE on Expression, Synthesis, and Localization of MTs in Rat Tissues
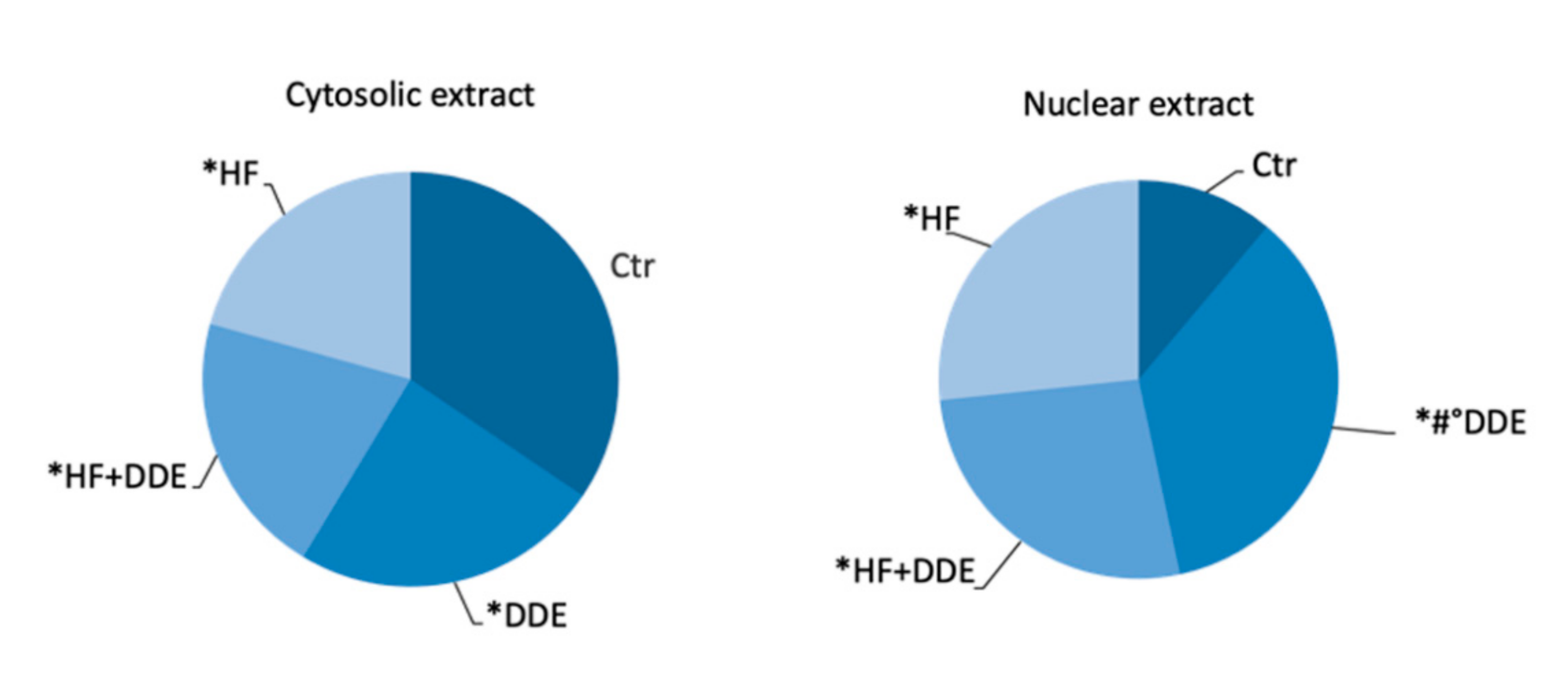
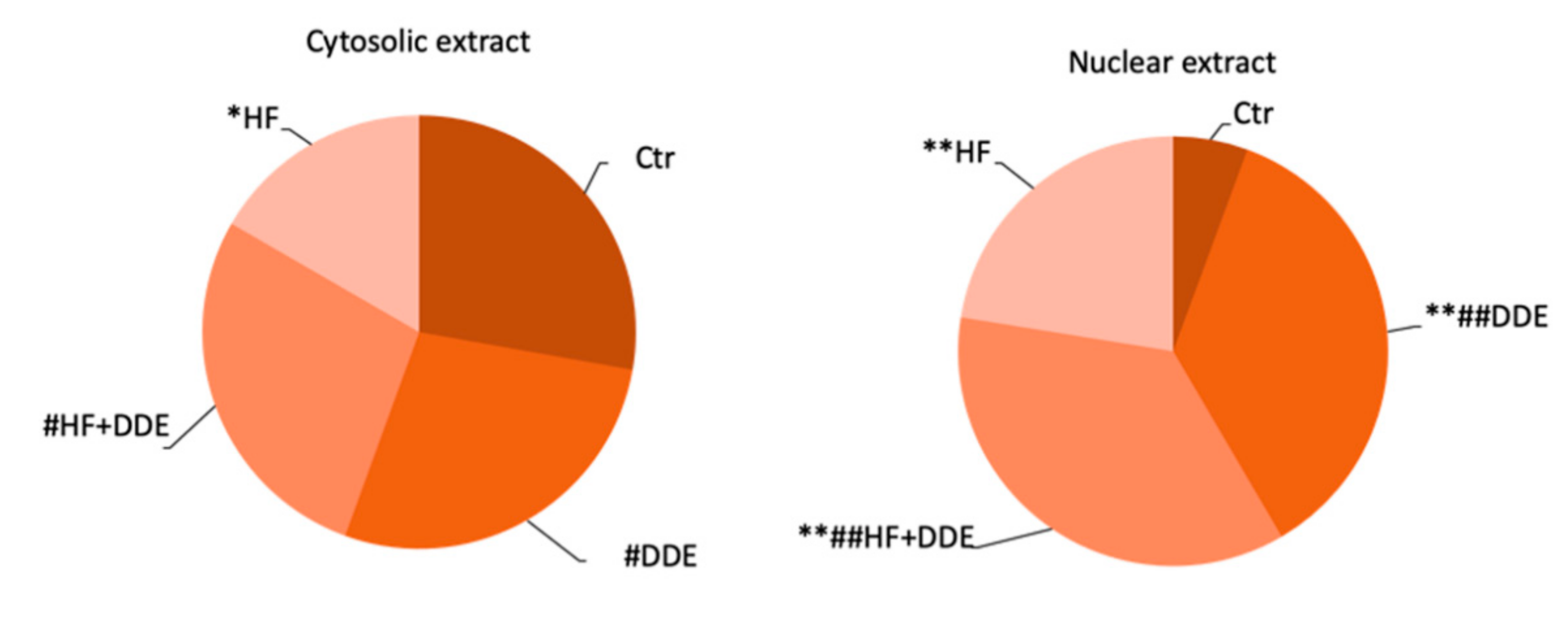
3.1. Liver
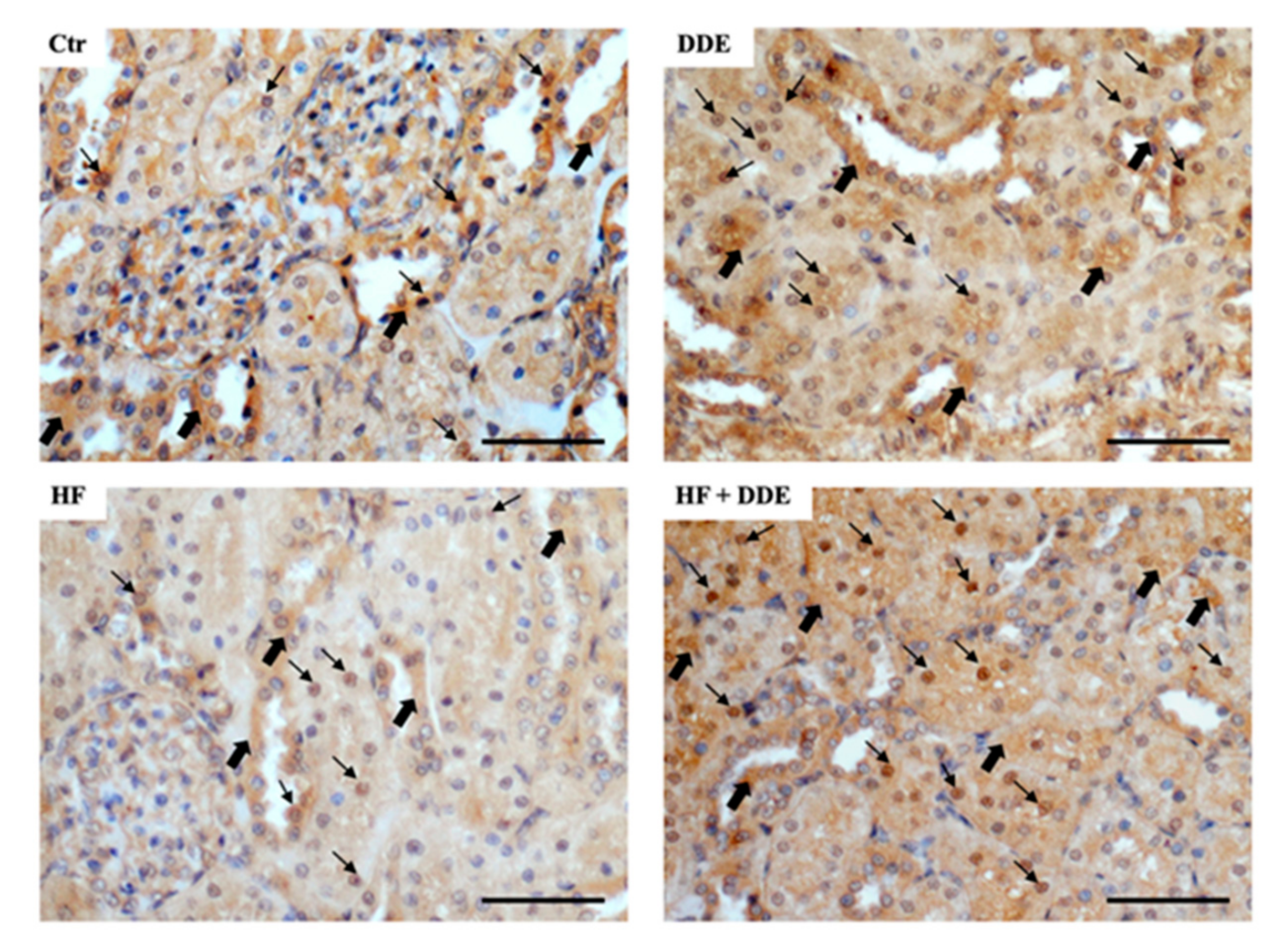
3.2. Kidneys
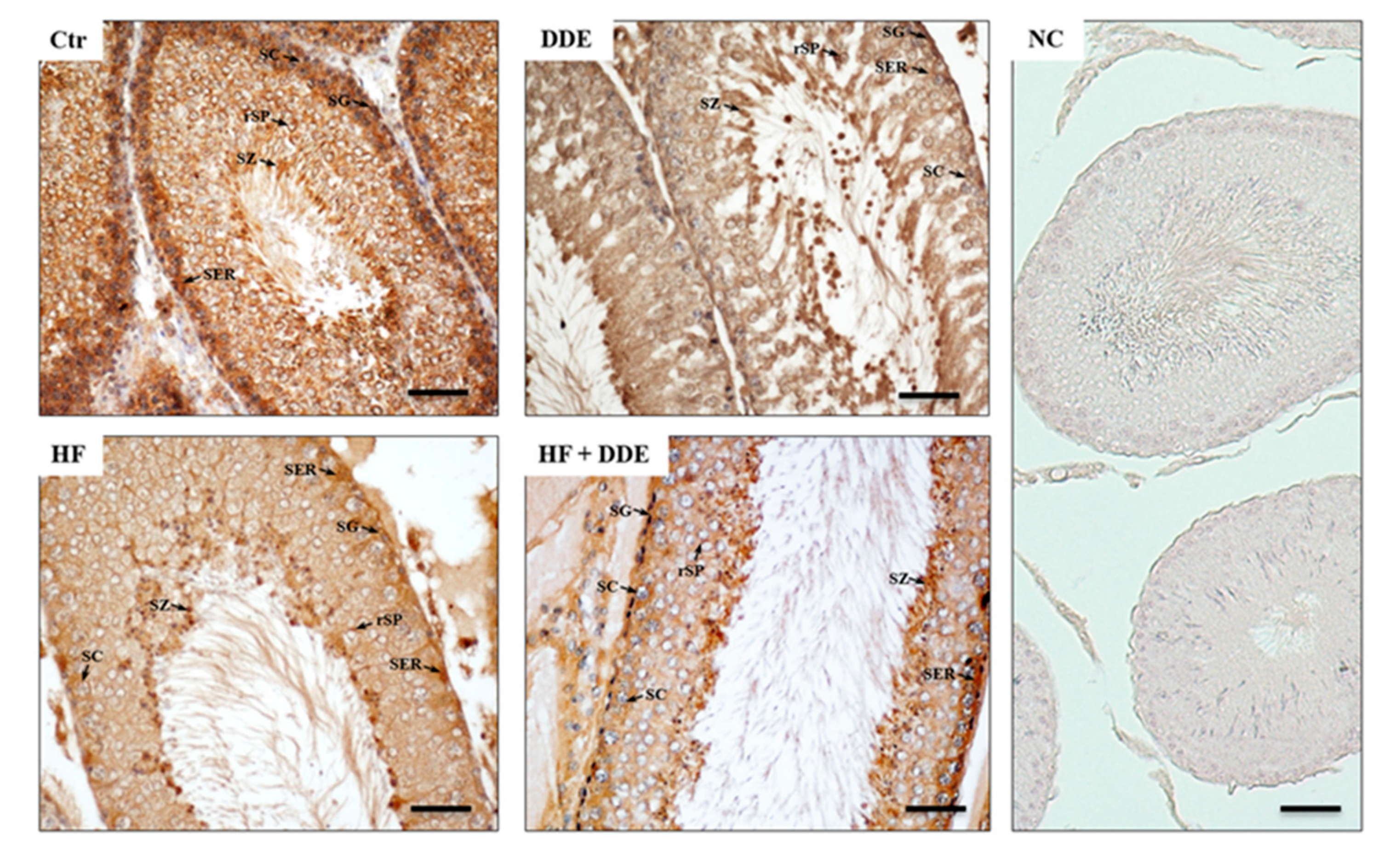
3.3. Testes
4. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Kagi, J.H.R.; Vallee, B.L. Metallothionein: A cadmium and zinc-containign protein from equine renal cortex. II. Physico-chemical properties. J. Biol. Chem. 1961, 236, 2435–2442. [Google Scholar] [PubMed]
- Webb, M. Toxicological significance of metallothionein. Exp. Suppl. 1987, 52, 109–134. [Google Scholar]
- Romero-Isart, N.; Vasák, M. Advances in the structure and chemistry of metallothioneins. J. Inorg. Biochem. 2002, 88, 388–396. [Google Scholar] [CrossRef]
- Isani, G.; Carpenè, E. Metallothioneins, unconventional proteins from unconventional animals: A long journey from nematodes to mammals. Biomolecules 2014, 4, 435–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziller, A.; Fraissinet-Tachet, L. Metallothionein diversity and distribution in the tree of life: A multifunctional protein. Metallomics 2018, 10, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef]
- Andrews, G.K. Regulation of metallothionein gene expression. Prog. Food Nutr. Sci. 1990, 14, 193–258. [Google Scholar]
- Scudiero, R.; Carginale, V.; Capasso, C.; Riggio, M.; Filosa, S.; Parisi, E. Structural and functional analysis of metal regulatory elements in the promoter region of genes encoding metallothionein isoforms in the Antarctic fish Chionodraco hamatus (icefish). Gene 2001, 274, 199–208. [Google Scholar] [CrossRef]
- Moleirinho, A.; Carneiro, J.; Matthiesen, R.; Silva, R.M.; Amorim, A.; Azevedo, L. Gains, losses and changes of function after gene duplication: Study of the metallothionein family. PLoS ONE 2011, 6, e18487. [Google Scholar] [CrossRef] [Green Version]
- Palmiter, R.D.; Findley, S.D.; Whitmore, T.E.; Durnam, D.D. MT-III, a brain-specific member of the metallothionein gene family. Proc. Natl. Acad. Sci. USA 1992, 89, 6333–6337. [Google Scholar] [CrossRef] [Green Version]
- Quaife, C.J.; Findley, S.D.; Erickson, J.C.; Froelick, G.J.; Kelly, E.J.; Zambrowicz, B.P.; Palmiter, R.D. Induction of a new metallothionein isoform (MT-IV) occurs during differentiation of stratified squamous epithelia. Biochemistry 1994, 33, 7250–7259. [Google Scholar] [CrossRef] [PubMed]
- Valls, M.; Bofill, R.; Gonzales-Duarte, R.; Gonzales-Duarte, P.; Capdevila, M.; Atrian, S. A new insight into metallothionein (MT) classification and evolution. The in vivo and in vitro metal binding features of Homarus americanus recombinant MT. J. Biol. Chem. 2001, 276, 32835–32843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios, O.; Atrian, S.; Capdevila, M. Zn- and Cu-thioneins: A functional classification for metallothioneins? J. Biol. Inorg. Chem. 2011, 16, 991–1009. [Google Scholar] [CrossRef]
- Artells, E.; Palacios, O.; Capdevila, M.; Atrian, S. Mammalian MT1 and MT2 metallothioneins differ in their metal binding abilities. Metallomics 2013, 5, 1397–1410. [Google Scholar] [CrossRef] [PubMed]
- Comes, G.; Fernandez-Gayol, O.; Molinero, A.; Giralt, M.; Capdevila, M.; Atrian, S.; Hidalgo, J. Mouse metallothionein-1 and metallothionein-2 are not biologically interchangeable in an animal model of multiple sclerosis, EAE. Metallomics 2019, 11, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Thirumoorthy, N.; Manisenthil Kumar, K.T.; Shyam Sundar, A.; Panayappan, L.; Chatterjee, M. Metallothionein: An overview. World J. Gastroenterol. 2007, 13, 993–996. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Koh, J.Y. Roles of zinc and metallothionein-3 in oxidative stress-induced lysosomal dysfunction, cell death, and autophagy in neurons and astrocytes. Mol. Brain 2010, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Namdarghanbari, M.; Wobig, W.; Krezoski, S.; Tabatabai, N.M.; Petering, D.H. Mammalian metallothionein in toxicology, cancer, and cancer chemotherapy. J. Biol. Inorg. Chem. 2011, 16, 1087–1101. [Google Scholar] [CrossRef]
- Werynska, B.; Pula, B.; Kobierzycki, C.; Dziegiel, P.; Podhorska-Okolow, M. Metallothioneins in the lung cancer. Folia Histochem. Cytobiol. 2015, 53, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dziegiel, P.; Pula, B.; Kobierzycki, C.; Stasiolek, M.; Podhorska-Okolow, M. Metallothioneins in normal and cancer cells. Adv. Anat. Embryol. Cell Biol. 2016, 218, 1–117. [Google Scholar]
- Juárez-Rebollar, D.; Rios, C.; Nava-Ruíz, C.; Méndez-Armenta, M. Metallothionein in brain disorders. Oxid. Med. Cell. Longev. 2017. [Google Scholar] [CrossRef] [Green Version]
- Chubatsu, L.S.; Meneghini, R. Metallothionein protects DNA from oxidative damage. Biochem. J. 1993, 291, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Chiaverini, N.; De Ley, M. Protective effect of metallothionein on oxidative stress-induced DNA damage. Free Radic. Res. 2010, 44, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Trinchella, F.; Riggio, M.; Filosa, S.; Volpe, M.G.; Parisi, E.; Scudiero, R. Cadmium distribution and metallothionein expression in lizard tissues following acute and chronic cadmium intoxication. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 144, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Cretì, P.; Trinchella, F.; Scudiero, R. Heavy metals bioaccumulation and metallothionein content in tissues of the sea bream Sparus aurata from three different fish farming systems. Environ. Monit. Assess. 2010, 165, 321–329. [Google Scholar] [CrossRef]
- Scudiero, R.; Cretì, P.; Trinchella, F.; Esposito, M.G. Evaluation of cadmium, lead and metallothionein contents in the tissues of mussels (Mytilus galloprovincialis) from the Campania coast (Italy): Levels and seasonal trends. C. R. Biol. 2014, 337, 451–458. [Google Scholar] [CrossRef]
- Capaldo, A.; Gay, F.; Scudiero, R.; Trinchella, F.; Caputo, I.; Lepretti, M.; Marabotti, A.; Esposito, C.; Laforgia, V. Histological changes, apoptosis and metallothionein levels in Triturus carnifex (Amphibia, Urodela) exposed to environmental cadmium concentrations. Aquat. Toxicol. 2016, 173, 63–73. [Google Scholar] [CrossRef]
- Verderame, M.; Limatola, E.; Scudiero, R. Metallothionein expression and synthesis in the testis of the lizard Podarcis sicula under natural conditions and following estrogenic exposure. Eur. J. Histochem. 2017, 61, 90–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scudiero, R.; Verderame, M.; Motta, C.M.; Simoniello, P. Unravelling the role of metallothionein on development, reproduction and detoxification in the wall lizard Podarcis sicula. Int. J. Mol. Sci. 2017, 18, 1569. [Google Scholar] [CrossRef] [Green Version]
- Erdoğan, O.; Ceyhun, S.B.; Ekinci, D.; Aksakal, E. Impact of deltamethrin exposure on mRNA expression levels of metallothionein A, B and cytochrome P450 1A in rainbow trout muscles. Gene 2011, 484, 13–17. [Google Scholar] [CrossRef]
- Mosleh, Y.Y.; Paris-Palacios, S.; Arnoult, F.; Couderchet, M.; Biagianti-Risbourg, S.; Vernet, G. Metallothionein induction in aquatic oligochaete Tubifex tubifex exposed to herbicide isoproturon. Environ. Toxicol. 2004, 19, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Won, J.H.; Ahn, K.H.; Back, M.J.; Fu, Z.; Jang, J.M.; Ha, H.C.; Jang, Y.J.; Kim, D.K. Paraquat reduces natural killer cell activity via metallothionein induction. J. Immunotoxicol. 2015, 12, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verderame, M.; Scudiero, R. How glyphosate impairs liver condition in the field lizard Podarcis siculus (Rafinesque-Schmaltz, 1810): Histological and molecular evidence. Biomed. Res. Int. 2019, 4746283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deichmann, W.B. The debate on DDT. Arch. Toxicol. 1972, 29, 1–27. [Google Scholar] [CrossRef]
- Nájera, J.A.; González-Silva, M.; Alonso, P.L. Some lessons for the future from the Global Malaria Eradication Programme (1955–1969). PLoS Med. 2011, 8, e1000412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrema, E.J.; Rubino, F.M.; Brambilla, G.; Moretto, A.; Tsatsakis , A.M.; Colosio, C. Persistent organochlorinated pesticides and mechanisms of their toxicity. Toxicology 2013, 307, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Yohannes, Y.B.; Ikenaka, Y.; Ito, G.; Nakayama, S.M.M.; Mizukawa, H.; Wepener, V.; Smit, N.J.; Van Vuren, J.H.J.; Ishizuka, M. Assessment of DDT contamination in house rat as a possible bioindicator in DDT-sprayed areas from Ethiopia and South Africa. Environ. Sci. Pollut. Res. Int. 2017, 24, 23763–23770. [Google Scholar] [CrossRef]
- Turgut, C.; Atatanir, L.; Mazmanci, B.; Mazmanci, M.A.; Henkelmann, B.; Schramm, K.W. The occurrence and environmental effect of persistent organic pollutants (POPs) in Taurus Mountains soils. Environ. Sci. Pollut. Res. Int. 2012, 19, 325–334. [Google Scholar] [CrossRef]
- Aislabie, J.M.; Richards, N.K.; Boul, H.L. Microbial degradation of DDT and its residues—A review. N. Z. J. Agric. Res. 1997, 40, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Jaga, K.; Dharmani, C. Global surveillance of DDT and DDE levels in human tissues. Int. J. Occup. Med. Environ. Health 2003, 16, 7–20. [Google Scholar]
- Tebourbi, O.; Driss, M.R.; Sakly, M.; Rhouma, K.B. Metabolism of DDT in different tissues of young rats. J. Environ. Sci. Health B 2006, 41, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, T.P.; Crispo Smith, S.; Park, J.S.; Petreas, M.X.; Rappaport, S.M.; Metayer, C. Concentrations of persistent organic pollutants in California children’s whole blood and residential dust. Environ. Sci. Technol. 2015, 49, 9331–9340. [Google Scholar] [CrossRef] [PubMed]
- Dowling, V.; Hoarau, P.C.; Romeo, M.; O’Halloran, J.; van Pelt, F.; O’Brien, N.; Sheehan, D. Protein carbonylation and heat shock response in Ruditapes decussatus following p,p’-dichlorodiphenyldichloroethylene (DDE) exposure: A proteomic approach reveals that DDE causes oxidative stress. Aquat. Toxicol. 2006, 77, 11–18. [Google Scholar] [CrossRef]
- Mota, P.C.; Cordeiro, M.; Pereira, S.P.; Oliveira, P.J.; Moreno, A.J.; Ramalho-Santos, J. Differential effects of p,p’-DDE on testis and liver mitochondria: Implications for reproductive toxicology. Reprod. Toxicol. 2011, 31, 80–85. [Google Scholar] [CrossRef]
- Marouani, N.; Hallegue, D.; Sakly, M.; Benkhalifa, M.; Ben Rhouma, K.; Tebourbi, O. p,p’-DDT induces testicular oxidative stress-induced apoptosis in adult rats. Reprod. Biol. Endocrinol. 2017, 15, 40. [Google Scholar] [CrossRef] [Green Version]
- Marouani, N.; Hallegue, D.; Sakly, M.; Benkhalifa, M.; Ben Rhouma, K.; Tebourbi, O. Involvment of oxidative stress in the mechanism of p,p’-DDT-induced nephrotoxicity in adult rats. Gen. Physiol. Biophys. 2017, 36, 309–320. [Google Scholar] [CrossRef]
- Migliaccio, V.; Scudiero, R.; Sica, R.; Lionetti, L.; Putti, R. Oxidative stress and mitochondrial uncoupling protein 2 expression in hepatic steatosis induced by exposure to xenobiotic DDE and high fat diet in male Wistar rats. PLoS ONE 2019, 14, e0215955. [Google Scholar] [CrossRef]
- Migliaccio, V.; Di Gregorio, I.; Putti, R.; Lionetti, L. Mitochondrial involvement in the adaptive response to chronic exposure to environmental pollutants and high-fat feeding in a rat liver and testis. Cells 2019, 8, 834. [Google Scholar] [CrossRef] [Green Version]
- Ben Miled, H.; Barka, Z.B.; Hallègue, D.; Lahbib, K.; Ladjimi, M.; Tlili, M.; Sakly, M.; Rhouma, K.B.; Ksouri, R.; Tebourbi, O. Hepatoprotective activity of Rhus oxyacantha root cortex extract against DDT-induced liver injury in rats. Biomed. Pharmacother. 2017, 90, 203–215. [Google Scholar] [CrossRef]
- Sabolić, I.; Breljak, D.; Skarica, M.; Herak-Kramberger, C.M. Role of metallothionein in cadmium traffic and toxicity in kidneys and other mammalian organs. Biometals 2010, 23, 897–926. [Google Scholar] [CrossRef]
- Quan, C.; Shi, Y.; Wang, C.; Wang, C.; Yang, K. p,p’-DDE damages spermatogenesis via phospholipid hydroperoxide glutathione peroxidase depletion and mitochondria apoptosis pathway. Environ. Toxicol. 2016, 31, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Alhama, J.; Fuentes-Almagro, C.A.; Abril, N.; Michán, C. Alterations in oxidative responses and post-translational modification caused by p,p’-DDE in Mus spretus testes reveal Cys oxidation status in proteins related to cell-redox homeostasis and male fertility. Sci. Total Environ. 2018, 636, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Makita, Y.; Omura, M.; Tanaka, A.; Kiyohara, C. Effects of concurrent exposure to tributyltin and 1,1-dichloro-2,2bis(p-chlorophenyl)ethylene(p,p’-DDE) on immature male wistar rats. Basic Clin. Pharmacol. Toxicol. 2005, 97, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, R.; Cigliano, L.; Verderame, M. Age-related changes of metallothionein 1/2 and metallothionein 3 expression in rat brain. C. R. Biol. 2017, 340, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, V.; Lionetti, L.; Putti, R.; Sica, R.; Scudiero, R. Combined effects of DDE and hyperlipidic diet on metallothionein expression and synthesis in rat tissues. Environ. Toxicol. 2019, 34, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, V.; Sica, R.; Scudiero, R.; Simoniello, P.; Putti, R.; Lionetti, L. Physiological adaptation to simultaneous chronic exposure to high-fat diet and dichlorodipheniletylhene (DDE) in wistar rat testis. Cells 2019, 8, 443. [Google Scholar] [CrossRef] [Green Version]
- Vukovic, V.; Pheng, S.R.; Stewart, A.; Vik, C.H.; Hedley, D.W. Protection from radiation-induced DNA single-strand breaks by induction of nuclear metallothionein. Int. J. Radiat. Biol. 2000, 76, 757–762. [Google Scholar] [CrossRef]
- Verderame, M.; Migliaccio, V.; Scudiero, R. Role of estrogen receptors, P450 aromatase, PCNA and p53 in high-fat-induced impairment of spermatogenesis in rats. CR Biol. 2018, 341, 371–379. [Google Scholar] [CrossRef]
- Ryan, J.A.; Hightower, L.E. Stress proteins as molecular biomarkers for environmental toxicology. EXS 1996, 77, 411–424. [Google Scholar]
- Pedrini-Martha, V.; Niederwanger, M.; Kopp, R.; Schnegg, R.; Dallinger, R. Physiological, diurnal and stress-related variability of cadmium-metallothionein gene expression in land snails. PLoS ONE 2016, 11, e0150442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolide, A.D.; Zawdie, B.; Alemayehu, T.; Tadesse, S. Association of trace metal elements with lipid profiles in type 2 diabetes mellitus patients: A cross sectional study. BMC Endocr. Disord. 2017, 17, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherian, M.G. The significance of the nuclear and cytoplasmic localization of metallothionein in human liver and tumor cells. Environ. Health Perspect. 1994, 102, 131–135. [Google Scholar]
- Cherian, M.G.; Lau, J.C.; Apostolova, M.D.; Cai, L. The nuclear-cytoplasmic presence of Metallothionein in cells during differentiation and development. In Metallothionein IV; Klaasen, C., Ed.; Birkhaüser: Basel, Switzerland, 1999; pp. 291–294. [Google Scholar]
- Cherian, M.G.; Apostolova, M.D. Nuclear localization of metallothionein during cell proliferation and differentiation. Cell. Mol. Biol. (Noisy-le-Grand) 2000, 46, 347–356. [Google Scholar]
- Ogra, Y.; Suzuki, K.T. Nuclear trafficking of metallothionein: Possible mechanisms and current knowledge. Cell. Mol. Biol. (Noisy-le-Grand) 2000, 46, 357–365. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metallothionein Gene Expression | HFD | HFD + p,p-DDE | p,p-DDE | p,p-DDT | |
|---|---|---|---|---|---|
| Migliaccio et al., 2019. [56] | LIVER | ↓ | NTD ↓ | NTD ↓ | |
| TESTES | ↓ | NTD ↓ | NTD ↓ | ||
| KIDNEYS | ↓ | NTD ↑ | NTD ↑ | ||
| Marouani et al., 2017. [45] | TESTES | HD ↓ | |||
| Metallothionein Protein Levels | HFD | HFD + p,p-DDE | p,p-DDE | p,p-DDT | |
| Migliaccio et al., 2019. [56] | LIVER
| ↓ ↓ ↑ | NTD ↓ ↓ ↑ | NTD ↓ ↓ ↑ | |
| Migliaccio et al., 2019. [57] | TESTES
| ↓ | NTD ↓ | NTD ↓ | |
| Migliaccio et al., 2019. [56] | KIDNEYS
| ↓ - ↑ | NTD - - ↑ | NTD - - ↑ | |
| Marouani et al., 2017. [46] | KIDNEYS
| - | HD ↓ | ||
| Ben Miled et al., 2017. [49] | LIVER
| - | HD ↑ | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migliaccio, V.; Lionetti, L.; Putti, R.; Scudiero, R. Exposure to Dichlorodiphenyldichloroethylene (DDE) and Metallothionein Levels in Rats Fed with Normocaloric or High-Fat Diet: A Review. Int. J. Mol. Sci. 2020, 21, 1903. https://doi.org/10.3390/ijms21051903
Migliaccio V, Lionetti L, Putti R, Scudiero R. Exposure to Dichlorodiphenyldichloroethylene (DDE) and Metallothionein Levels in Rats Fed with Normocaloric or High-Fat Diet: A Review. International Journal of Molecular Sciences. 2020; 21(5):1903. https://doi.org/10.3390/ijms21051903
Chicago/Turabian StyleMigliaccio, Vincenzo, Lillà Lionetti, Rosalba Putti, and Rosaria Scudiero. 2020. "Exposure to Dichlorodiphenyldichloroethylene (DDE) and Metallothionein Levels in Rats Fed with Normocaloric or High-Fat Diet: A Review" International Journal of Molecular Sciences 21, no. 5: 1903. https://doi.org/10.3390/ijms21051903
APA StyleMigliaccio, V., Lionetti, L., Putti, R., & Scudiero, R. (2020). Exposure to Dichlorodiphenyldichloroethylene (DDE) and Metallothionein Levels in Rats Fed with Normocaloric or High-Fat Diet: A Review. International Journal of Molecular Sciences, 21(5), 1903. https://doi.org/10.3390/ijms21051903