Comparative Mitogenome Analysis of the Genus Trifolium Reveals Independent Gene Fission of ccmFn and Intracellular Gene Transfers in Fabaceae

Abstract
:1. Introduction
2. Results
2.1. Mitogenome Features of Four Trifolium Species
2.2. Repeat Composition of Organelle Genomes in Trifolium
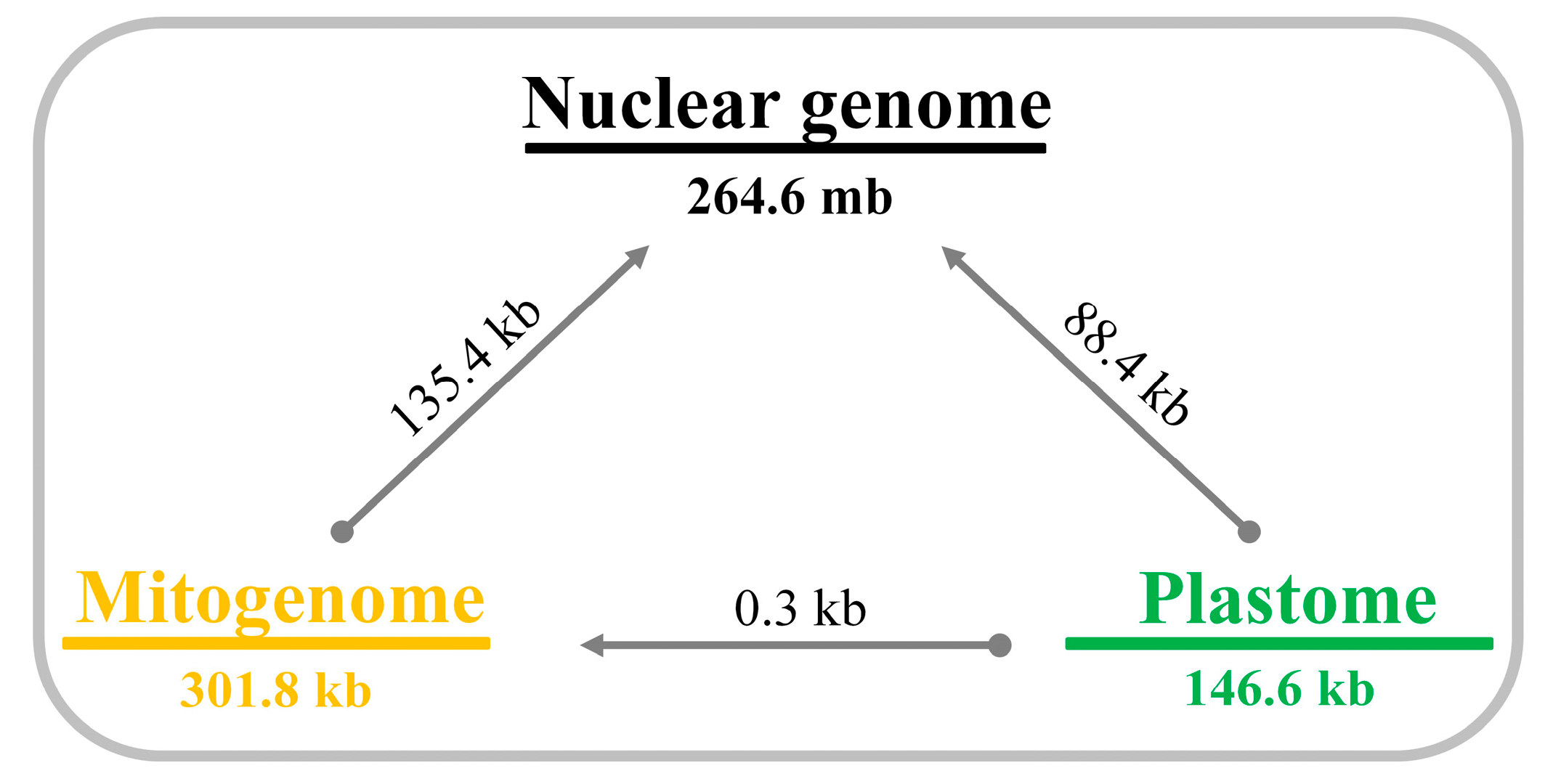
2.3. Intracellular Gene Transfer (IGT) in Trifolium
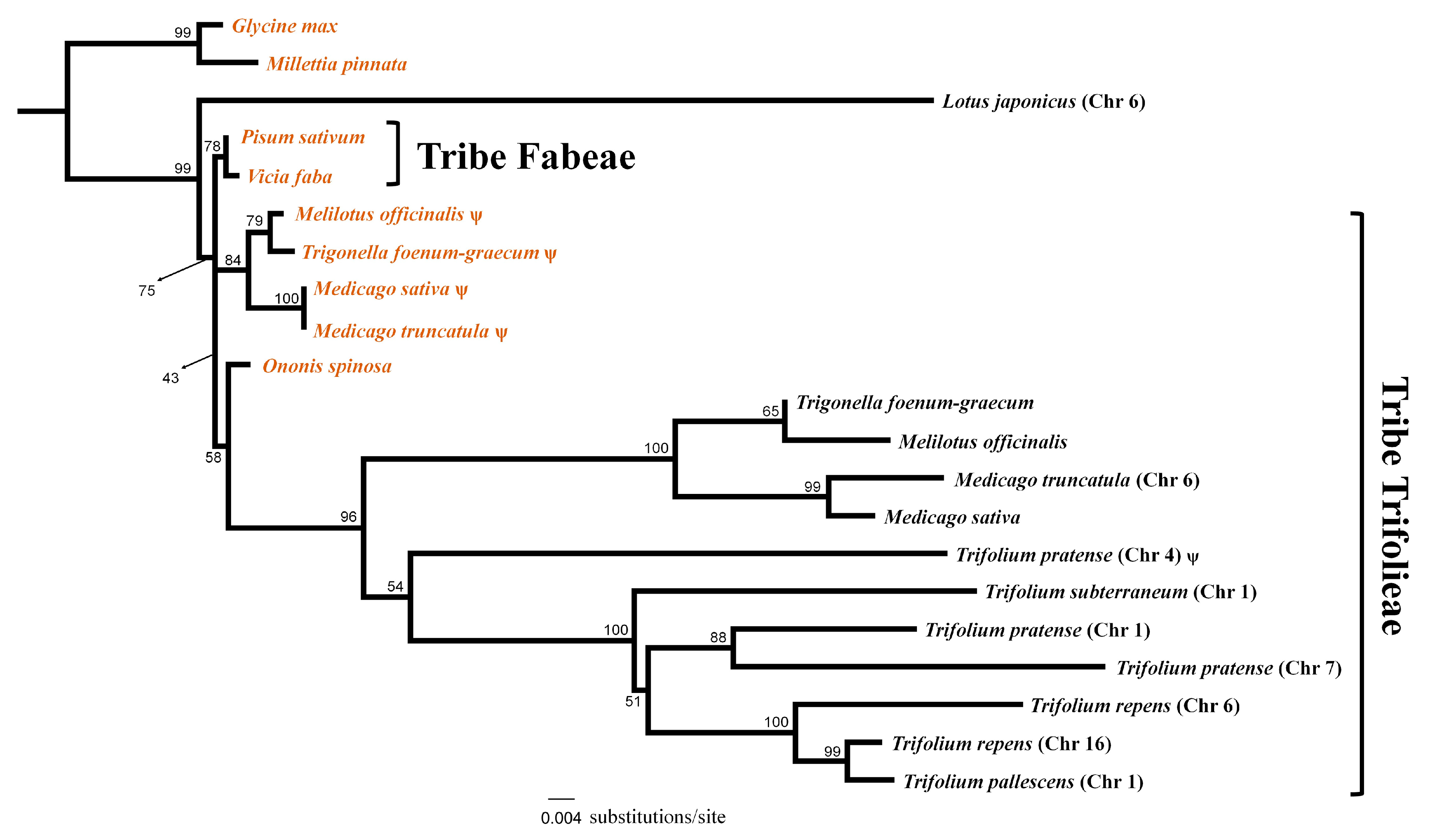
2.4. Multiple Functional Transfers of Mitochondrial rps1 in Papilionoideae
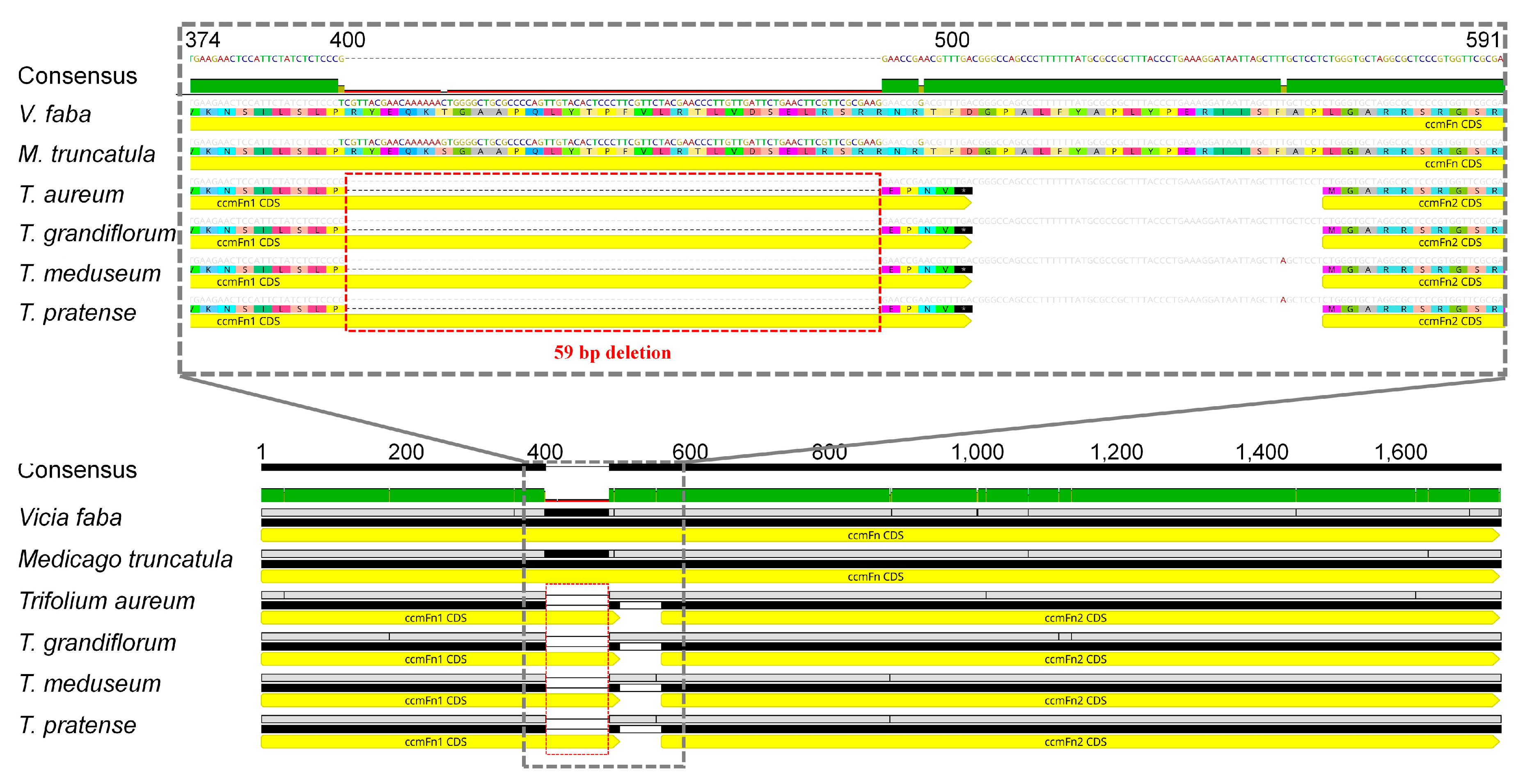
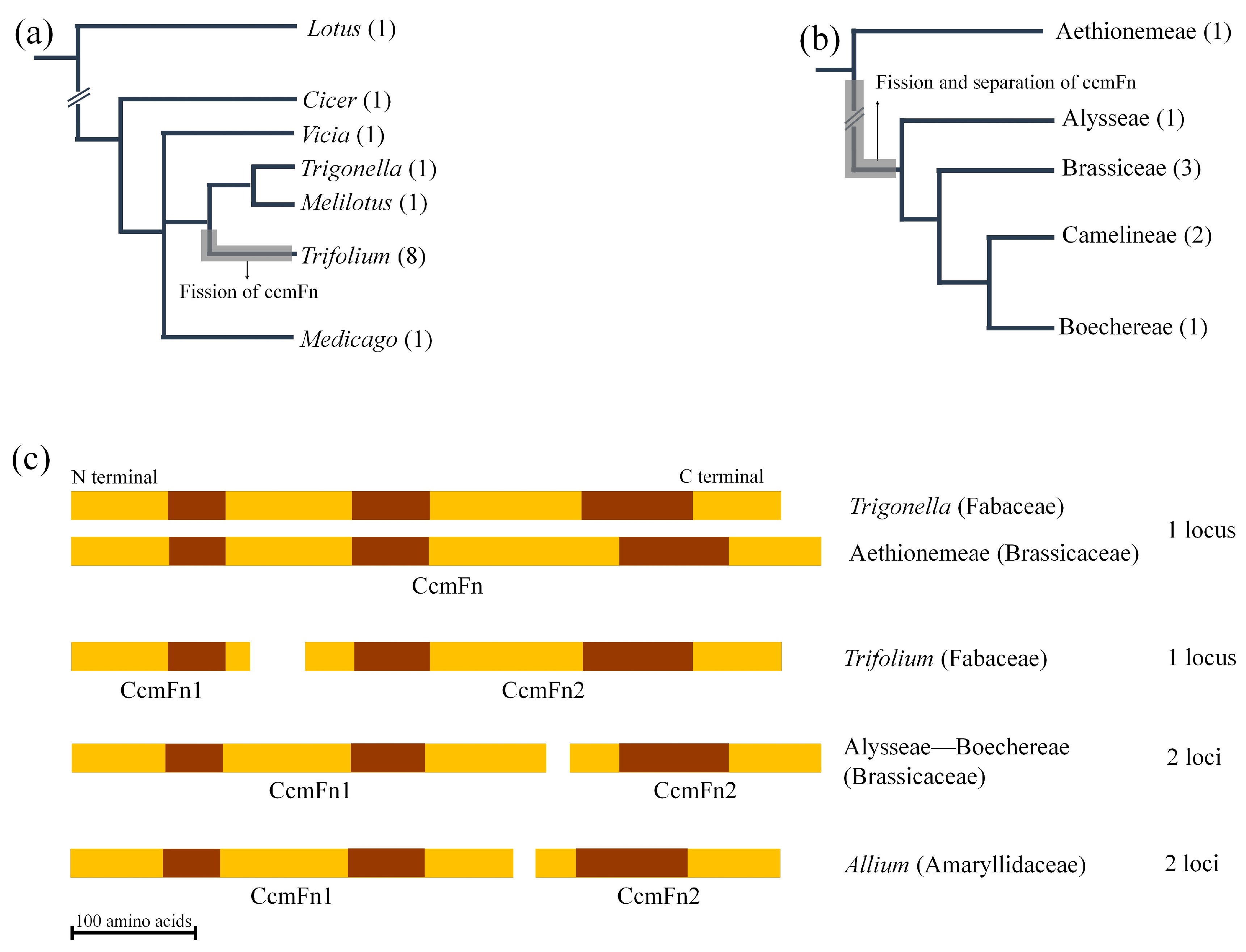
2.5. Fission of ccmF in Land Plants
3. Discussion
3.1. Contrasting Evolutionary Trajectories of Trifolium Organelle Genomes
3.2. Multiple Functional Transfers of the Mitochondrial rps1 Gene to the Nucleus in Papilionoideae
3.3. Shared DNA Among Genomes of Trifolium
3.4. Multiple Fissions of ccmF in Land Plants and a Novel Event in Trifolium
4. Materials and Methods
4.1. Assembly of Trifolium Mitogenomes
4.2. Annotation and Genome Content Comparison of Mitogenomes
4.3. Completion of the Trifolium Pratense Plastome
4.4. Repeat Estimation in Organelle Genomes
4.5. Shared DNA among Different Genomic Compartments
4.6. Investigation on Status of rps1 in Nuclear and Mitochondrial Genome
4.7. Investigation of ccmF Fissions in Selected Land Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| IGT | Intracellular gene transfer |
| NUMT | Nuclear mitochondrial DNA sequences |
| NUPT | Nuclear plastid DNA sequences |
| IR | Inverted repeat |
| IRLC | Inverted repeat lacking clade |
| ORF | Open reading frame |
| NGS | Next-generation sequencing |
| DNA-RRR | DNA replication, recombination and repair |
| ML | Maximum likelihood |
| DSB | Double strand break |
| NCBI | National Center for Biotechnology Information |
References
- Jansen, R.K.; Ruhlman, T.A. Plastid Genomes of Seed Plants. In Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 103–126. [Google Scholar]
- Ruhlman, T.A.; Jansen, R.K. The plastid genomes of flowering plants. In Chloroplast Biotechnology: Methods and Protocols; Maliga, P., Ed.; Spring: New York, NY, USA, 2014; pp. 3–38. [Google Scholar]
- Knoop, V. The mitochondrial DNA of land plants: Peculiarities in phylogenetic perspective. Curr. Genet. 2004, 46, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Wynn, E.L.; Christensen, A.C. Repeats of unusual size in plant mitochondrial genomes: Identification, incidence and evolution. G3 Genes Genomes Genet. 2019, 9, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, K.L.; Qiu, Y.-L.; Stoutemyer, M.; Palmer, J.D. Punctuated evolution of mitochondrial gene content: High and variable rates of mitochondrial gene loss and transfer to the nucleus during angiosperm evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 9905–9912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, T.; Mikami, T. Organization and variation of angiosperm mitochondrial genome. Physiol. Plant. 2007, 129, 6–13. [Google Scholar] [CrossRef]
- Gualberto, J.M.; Newton, K.J. Plant mitochondrial genomes: Dynamics and mechanisms of mutation. Annu. Rev. Plant Biol. 2017, 68, 225–252. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-S.; Schwarz, E.N.; Ruhlman, T.A.; Khiyami, M.A.; Sabir, J.S.; Hajarah, N.H.; Sabir, M.J.; Rabah, S.O.; Jansen, R.K. Fluctuations in Fabaceae mitochondrial genome size and content are both ancient and recent. BMC Plant Biol. 2019, 19, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, K.H.; Li, W.-H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [Green Version]
- Drouin, G.; Daoud, H.; Xia, J. Relative rates of synonymous substitutions in the mitochondrial, chloroplast and nuclear genomes of seed plants. Mol. Phylogenet. Evol. 2008, 49, 827–831. [Google Scholar] [CrossRef]
- Selosse, M.-A.; Albert, B.; Godelle, B. Reducing the genome size of organelles favours gene transfer to the nucleus. Trends Ecol. Evol. 2001, 16, 135–141. [Google Scholar] [CrossRef]
- Lopez, J.V.; Yuhki, N.; Masuda, R.; Modi, W.; O’Brien, S.J. Numt, a recent transfer and tandem amplification of mitochondrial DNA to the nuclear genome of the domestic cat. J. Mol. Evol. 1994, 39, 174–190. [Google Scholar]
- Timmis, J.N.; Ayliffe, M.A.; Huang, C.Y.; Martin, W. Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 2004, 5, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.O.; Palmer, J.D. Horizontal gene transfer in plants. J. Exp. Bot. 2006, 58, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonen, L. Mitochondrial genes leave home. New Phytol. 2006, 172, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Grünheit, N.; Ahmadinejad, N.; Timmis, J.N.; Martin, W. Mutational decay and age of chloroplast and mitochondrial genomes transferred recently to angiosperm nuclear chromosomes. Plant Physiol. 2005, 138, 1723–1733. [Google Scholar] [CrossRef] [Green Version]
- Adams, K.L.; Song, K.; Roessler, P.G.; Nugent, J.M.; Doyle, J.L.; Doyle, J.J.; Palmer, J.D. Intracellular gene transfer in action: Dual transcription and multiple silencings of nuclear and mitochondrial cox2 genes in legumes. Proc. Natl. Acad. Sci. USA 1999, 96, 13863–13868. [Google Scholar] [CrossRef] [Green Version]
- Laroche, J.; Li, P.; Maggia, L.; Bousquet, J. Molecular evolution of angiosperm mitochondrial introns and exons. Proc. Natl. Acad. Sci. USA 1997, 94, 5722–5727. [Google Scholar] [CrossRef] [Green Version]
- Adams, K.L.; Daley, D.O.; Qiu, Y.-L.; Whelan, J.; Palmer, J.D. Repeated, recent and diverse transfers of a mitochondrial gene to the nucleus in flowering plants. Nature 2000, 408, 354–357. [Google Scholar] [CrossRef]
- Adams, K.L.; Ong, H.C.; Palmer, J.D. Mitochondrial gene transfer in pieces: Fission of the ribosomal protein gene rpl2 and partial or complete gene transfer to the nucleus. Mol. Biol. Evol. 2001, 18, 2289–2297. [Google Scholar] [CrossRef] [Green Version]
- Szafranski, P. Intercompartmental piecewise gene transfer. Genes 2017, 8, 260. [Google Scholar] [CrossRef] [Green Version]
- Oda, K.; Yamato, K.; Ohta, E.; Nakamura, Y.; Takemura, M.; Nozato, N.; Akashi, K.; Kanegae, T.; Ogura, Y.; Kohchi, T. Gene organization deduced from the complete sequence of liverwort Marchantia polymorpha mitochondrial DNA: A primitive form of plant mitochondrial genome. J. Mol. Biol. 1992, 223, 1–7. [Google Scholar] [CrossRef]
- Handa, H.; Bonnard, G.R.; Grienenberger, J.-M. The rapeseed mitochondrial gene encoding a homologue of the bacterial protein Ccl1 is divided into two independently transcribed reading frames. Mol. Gen. Genet. 1996, 252, 292–302. [Google Scholar] [CrossRef]
- Unseld, M.; Marienfeld, J.R.; Brandt, P.; Brennicke, A. The mitochondrial genome of Arabidopsis thaliana contains 57 genes in 366,924 nucleotides. Nat. Genet. 1997, 15, 57–61. [Google Scholar] [CrossRef]
- Kim, B.; Kim, K.; Yang, T.-J.; Kim, S. Completion of the mitochondrial genome sequence of onion (Allium cepa L.) containing the CMS-S male-sterile cytoplasm and identification of an independent event of the ccmFN gene split. Curr. Genet. 2016, 62, 873–885. [Google Scholar] [CrossRef]
- Thöny-Meyer, L.; Fischer, F.; Künzler, P.; Ritz, D.; Hennecke, H. Escherichia coli genes required for cytochrome c maturation. J. Bacteriol. 1995, 177, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Giegé, P.; Grienenberger, J.; Bonnard, G. Cytochrome c biogenesis in mitochondria. Mitochondrion 2008, 8, 61–73. [Google Scholar] [CrossRef]
- LPWG, Legume Phylogeny Working Group. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny. Taxon 2017, 66, 44–77. [Google Scholar] [CrossRef] [Green Version]
- Lewis, G.P.; Schrire, B.; Mackinder, B.; Lock, M. Legumes of the World; Royal Botanic Gardens: Kew, UK, 2005. [Google Scholar]
- Wojciechowski, M.F.; Sanderson, M.J.; Steele, K.P.; Liston, A. Molecular phylogeny of the “temperate herbaceous tribes” of papilionoid legumes: A supertree approach. In Advances in Legume Systematics; Herendeen, P., Bruneau, A., Eds.; Royal Botanic Gardens: Kew, UK, 2000; Volume 9, pp. 277–298. [Google Scholar]
- Sabir, J.; Schwarz, E.; Ellison, N.; Zhang, J.; Baeshen, N.A.; Mutwakil, M.; Jansen, R.; Ruhlman, T. Evolutionary and biotechnology implications of plastid genome variation in the inverted-repeat-lacking clade of legumes. Plant Biotechnol. J. 2014, 12, 743–754. [Google Scholar] [CrossRef]
- Magee, A.M.; Aspinall, S.; Rice, D.W.; Cusack, B.P.; Sémon, M.; Perry, A.S.; Stefanović, S.; Milbourne, D.; Barth, S.; Palmer, J.D. Localized hypermutation and associated gene losses in legume chloroplast genomes. Genome Res. 2010, 20, 1700–1710. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Guisinger, M.; Kim, H.-G.; Ruck, E.; Blazier, J.C.; McMurtry, V.; Kuehl, J.V.; Boore, J.; Jansen, R.K. Extensive reorganization of the plastid genome of Trifolium subterraneum (Fabaceae) is associated with numerous repeated sequences and novel DNA insertions. J. Mol. Evol. 2008, 67, 696–704. [Google Scholar] [CrossRef]
- Sveinsson, S.; Cronk, Q. Evolutionary origin of highly repetitive plastid genomes within the clover genus (Trifolium). BMC Evol. Biol. 2014, 14, 228. [Google Scholar] [CrossRef] [Green Version]
- Milligan, B.G.; Hampton, J.N.; Palmer, J.D. Dispersed repeats and structural reorganization in subclover chloroplast DNA. Mol. Biol. Evol. 1989, 6, 355–368. [Google Scholar]
- Choi, I.S.; Jansen, R.; Ruhlman, T. Lost and Found: Return of the inverted repeat in the legume clade defined by its absence. Genome Biol. Evol. 2019, 11, 1321–1333. [Google Scholar] [CrossRef] [Green Version]
- Negruk, V. Mitochondrial genome sequence of the legume Vicia faba. Front. Plant Sci. 2013, 4, 128. [Google Scholar] [CrossRef] [Green Version]
- Bi, C.; Wang, X.; Xu, Y.; Wei, S.; Shi, Y.; Dai, X.; Yin, T.; Ye, N. The complete mitochondrial genome of Medicago truncatula. Mitochondrial DNA B 2016, 1, 122–123. [Google Scholar] [CrossRef] [Green Version]
- Hazle, T.; Bonen, L. Status of genes encoding the mitochondrial S1 ribosomal protein in closely-related legumes. Gene 2007, 405, 108–116. [Google Scholar] [CrossRef]
- Kazakoff, S.H.; Imelfort, M.; Edwards, D.; Koehorst, J.; Biswas, B.; Batley, J.; Scott, P.T.; Gresshoff, P.M. Capturing the biofuel wellhead and powerhouse: The chloroplast and mitochondrial genomes of the leguminous feedstock tree Pongamia pinnata. PLoS ONE 2012, 7, e51687. [Google Scholar] [CrossRef]
- Lock, J. Trifolieae. In Legumes of the World; Lewis, G.P., Schrire, B., Mackinder, B., Lock, M., Eds.; Royal Botanic Gardens: Kew, UK, 2005; pp. 499–504. [Google Scholar]
- Ellison, N.W.; Liston, A.; Steiner, J.J.; Williams, W.M.; Taylor, N.L. Molecular phylogenetics of the clover genus (Trifolium—Leguminosae). Mol. Phylogenet. Evol. 2006, 39, 688–705. [Google Scholar] [CrossRef]
- De Vega, J.J.; Ayling, S.; Hegarty, M.; Kudrna, D.; Goicoechea, J.L.; Ergon, Å.; Rognli, O.A.; Jones, C.; Swain, M.; Geurts, R. Red clover (Trifolium pratense L.) draft genome provides a platform for trait improvement. Sci. Rep. 2015, 5, 17394. [Google Scholar] [CrossRef]
- Hirakawa, H.; Kaur, P.; Shirasawa, K.; Nichols, P.; Nagano, S.; Appels, R.; Erskine, W.; Isobe, S.N. Draft genome sequence of subterranean clover, a reference for genus Trifolium. Sci. Rep. 2016, 6, 30358. [Google Scholar] [CrossRef] [Green Version]
- Dudchenko, O.; Pham, M.; Lui, C.; Batra, S.S.; Hoeger, M.; Nyquist, S.K.; Durand, N.C.; Shamim, M.S.; Machol, I.; Erskine, W. Hi-C yields chromosome-length scaffolds for a legume genome, Trifolium subterraneum. bioRxiv 2018, 473553. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, A.G.; Moraga, R.; Tausen, M.; Gupta, V.; Bilton, T.P.; Campbell, M.A.; Ashby, R.; Nagy, I.; Khan, A.; Larking, A. Breaking free: The genomics of allopolyploidy-facilitated niche expansion in white clover. Plant Cell 2019, 31, 1466–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-H.; Sun, R.; Hu, Y.; Zeng, L.; Zhang, N.; Cai, L.; Zhang, Q.; Koch, M.A.; Al-Shehbaz, I.; Edger, P.P. Resolution of Brassicaceae phylogeny using nuclear genes uncovers nested radiations and supports convergent morphological evolution. Mol. Biol. Evol. 2015, 33, 394–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, A.C. Plant mitochondrial genome evolution can be explained by DNA repair mechanisms. Genome Biol. Evol. 2013, 5, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.C. Genes and junk in plant mitochondria—Repair mechanisms and selection. Genome Biol. Evol. 2014, 6, 1448–1453. [Google Scholar] [CrossRef] [Green Version]
- Christensen, A.C. Mitochondrial DNA repair and genome evolution. In Annual Plant Reviews; Logan, D.C., Ed.; Wiley-Blackwell: New York, NY, USA, 2018; Volume 50, pp. 11–32. [Google Scholar]
- Zhang, J.; Ruhlman, T.A.; Sabir, J.S.; Blazier, J.C.; Weng, M.-L.; Park, S.; Jansen, R.K. Coevolution between nuclear-encoded DNA replication, recombination, and repair genes and plastid genome complexity. Genome Biol. Evol. 2016, 8, 622–634. [Google Scholar] [CrossRef] [Green Version]
- Bergsten, J. A review of long-branch attraction. Cladistics 2005, 21, 163–193. [Google Scholar] [CrossRef]
- Ong, H.C.; Palmer, J.D. Pervasive survival of expressed mitochondrial rps14 pseudogenes in grasses and their relatives for 80 million years following three functional transfers to the nucleus. BMC Evol. Biol. 2006, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Noutsos, C.; Richly, E.; Leister, D. Generation and evolutionary fate of insertions of organelle DNA in the nuclear genomes of flowering plants. Genome Res. 2005, 15, 616–628. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Kaul, S.; Rounsley, S.; Shea, T.P.; Benito, M.-I.; Town, C.D.; Fujii, C.Y.; Mason, T.; Bowman, C.L.; Barnstead, M. Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana. Nature 1999, 402, 761–768. [Google Scholar] [CrossRef]
- Stupar, R.M.; Lilly, J.W.; Town, C.D.; Cheng, Z.; Kaul, S.; Buell, C.R.; Jiang, J. Complex mtDNA constitutes an approximate 620-kb insertion on Arabidopsis thaliana chromosome 2: Implication of potential sequencing errors caused by large-unit repeats. Proc. Natl. Acad. Sci. USA 2001, 98, 5099–5103. [Google Scholar] [CrossRef] [Green Version]
- Cousins, G.; Woodfield, D. Effect of inbreeding on growth of white clover. In Proceedings of the 13th Australasian Plant Breeding Conference, Christchurch, New Zealand, 18–21 April 2006; pp. 568–572. [Google Scholar]
- Venkatesh, B.; Dandona, N.; Brenner, S. Fugu genome does not contain mitochondrial pseudogenes. Genomics 2006, 87, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Xing, Y.; Mao, X. The little brown bat nuclear genome contains an entire mitochondrial genome: Real or artifact? Gene 2017, 629, 64–67. [Google Scholar] [CrossRef]
- Rayapuram, N.; Hagenmuller, J.; Grienenberger, J.M.; Bonnard, G.; Giegé, P. The three mitochondrial encoded CcmF proteins form a complex that interacts with CCMH and c-type apocytochromes in Arabidopsis. J. Biol. Chem. 2008, 283, 25200–25208. [Google Scholar] [CrossRef] [Green Version]
- Kisiel, K.; Miwa, H.; Odrzykoski, I.J. Taxonomic identification of chloroplast genome of Marchantia polymorpha using DNA barcode sequences. In Proceedings of the Fourth International Barcode of Life Conference, Adelaide, Australia, 28 November–3 December 2011. [Google Scholar]
- Grewe, F.; Edger, P.P.; Keren, I.; Sultan, L.; Pires, J.C.; Ostersetzer-Biran, O.; Mower, J.P. Comparative analysis of 11 Brassicales mitochondrial genomes and the mitochondrial transcriptome of Brassica oleracea. Mitochondrion 2014, 19, 135–143. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq–versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Grewe, F.; Fan, W.; Young, G.J.; Knoop, V.; Palmer, J.D.; Mower, J.P. Ginkgo and Welwitschia mitogenomes reveal extreme contrasts in gymnosperm mitochondrial evolution. Mol. Biol. Evol. 2016, 33, 1448–1460. [Google Scholar] [CrossRef] [Green Version]
- Darzentas, N. Circoletto: Visualizing sequence similarity with Circos. Bioinformatics 2010, 26, 2620–2621. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T.; Jenkins, J.; Shu, S.; Ishizaki, K.; Yamaoka, S.; Nishihama, R.; Nakamura, Y.; Berger, F. Insights into land plant evolution garnered from the Marchantia polymorpha genome. Cell 2017, 171, 287–304. [Google Scholar] [CrossRef]
- Marks, R.A.; Smith, J.J.; Cronk, Q.; Grassa, C.J.; McLetchie, D.N. Genome of the tropical plant Marchantia inflexa: Implications for sex chromosome evolution and dehydration tolerance. Sci. Rep. 2019, 9, 8722. [Google Scholar] [CrossRef]
- Kwon, W.; Kim, Y.; Park, J. The complete chloroplast genome sequence of Dumortiera hirsuta (Sw.) Nees (Marchantiophyta, Dumortieraceae). Mitochondrial DNA B 2019, 4, 318–319. [Google Scholar] [CrossRef] [Green Version]
- Myszczyński, K.; Ślipiko, M.; Sawicki, J. Potential of transcript editing across mitogenomes of early land plants shows novel and familiar trends. Int. J. Mol. Sci. 2019, 20, 2963. [Google Scholar] [CrossRef] [Green Version]
- Pagni, M.; Ioannidis, V.; Cerutti, L.; Zahn-Zabal, M.; Jongeneel, C.V.; Hau, J.; Martin, O.; Kuznetsov, D.; Falquet, L. MyHits: Improvements to an interactive resource for analyzing protein sequences. Nucleic Acids Res. 2007, 35, W433–W437. [Google Scholar] [CrossRef]
- Sigrist, C.J.; Cerutti, L.; De Castro, E.; Langendijk-Genevaux, P.S.; Bulliard, V.; Bairoch, A.; Hulo, N. PROSITE, a protein domain database for functional characterization and annotation. Nucleic Acids Res. 2009, 38, D161–D166. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Subgenus | Raw Reads | Mitogenome Coverage | Mitogenome Length (bp) | GC (%) | NCBI Accession |
|---|---|---|---|---|---|---|
| T. aureum | Chronosemium | 71,346,514 | 645 | 294,911 | 44.9 | MT039392 |
| T. grandiflorum | Chronosemium | 48,390,678 | 197 | 347,723 | 45.1 | MT039391 |
| T. meduseum | Trifolium | 68,712,286 | 207 | 348,724 | 45.0 | MT039390 |
| T. pratense | Trifolium | 47,909,108 | 212 | 301,823 | 45.2 | MT039389 |
| Species | Subgenus | Section | Mitogenome | Plastome | ||||
|---|---|---|---|---|---|---|---|---|
| Size (bp) | Repeat (%) | NCBI Accession | Size (bp) | Repeat (%) | NCBI Accession | |||
| T. aureum | Chronosemium | Chronosemium | 294,911 | 8.6 | MT039392 | 126,970 | 5.2 | NC_024035 |
| T. grandiflorum | Chronosemium | Chronosemium | 347,723 | 8.4 | MT039391 | 125,628 | 4.7 | NC_024034 |
| T. boissieri | Chronosemium | Chronosemium | Not applicable | 125,740 | 5.1 | NC_025743 | ||
| T. strictum | Trifolium | Paramesus | Not applicable | 125,834 | 4.4 | NC_025745 | ||
| T. glanduliferum | Trifolium | Paramesus | Not applicable | 126,149 | 4.8 | NC_025744 | ||
| T. lupinaster | Trifolium | Lupinaster | Not applicable | 135,049 | 10.9 | KJ788287 | ||
| T. subterraneum | Trifolium | Trichocephalum | Not applicable | 144,763 | 19.7 | NC_011828 | ||
| T. meduseum | Trifolium | Trichocephalum | 348,724 | 8.5 | MT039390 | 142,595 | 19.5 | NC_024166 |
| T. pratense | Trifolium | Trifolium | 301,823 | 6.6 | MT039389 | 146,573 | 20.7 | MT039393 |
| T. hybridum | Trifolium | Vesicastrum | Not applicable | 134,831 | 13.1 | KJ788286 | ||
| T. semipilosum | Trifolium | Vesicastrum | Not applicable | 138,194 | 15.8 | KJ788291 | ||
| T. repens | Trifolium | Trifoliastrum | Not applicable | 132,120 | 10.7 | NC_024036 | ||
| T. occidentale | Trifolium | Trifoliastrum | Not applicable | 133,806 | 11.1 | KJ788289 | ||
| Comparison | Number | Average Identity | GC (%) | Length (bp) | |||
|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Median | ||||
| Nuclear vs. Mitochondria | 1830 | 95.5 | 45.8 | 33 | 3950 | 121.8 | 93.5 |
| Nuclear vs. Plastid | 1086 | 95.7 | 35.1 | 34 | 2027 | 144.2 | 118.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, I.-S.; Ruhlman, T.A.; Jansen, R.K. Comparative Mitogenome Analysis of the Genus Trifolium Reveals Independent Gene Fission of ccmFn and Intracellular Gene Transfers in Fabaceae. Int. J. Mol. Sci. 2020, 21, 1959. https://doi.org/10.3390/ijms21061959
Choi I-S, Ruhlman TA, Jansen RK. Comparative Mitogenome Analysis of the Genus Trifolium Reveals Independent Gene Fission of ccmFn and Intracellular Gene Transfers in Fabaceae. International Journal of Molecular Sciences. 2020; 21(6):1959. https://doi.org/10.3390/ijms21061959
Chicago/Turabian StyleChoi, In-Su, Tracey A. Ruhlman, and Robert K. Jansen. 2020. "Comparative Mitogenome Analysis of the Genus Trifolium Reveals Independent Gene Fission of ccmFn and Intracellular Gene Transfers in Fabaceae" International Journal of Molecular Sciences 21, no. 6: 1959. https://doi.org/10.3390/ijms21061959
APA StyleChoi, I. -S., Ruhlman, T. A., & Jansen, R. K. (2020). Comparative Mitogenome Analysis of the Genus Trifolium Reveals Independent Gene Fission of ccmFn and Intracellular Gene Transfers in Fabaceae. International Journal of Molecular Sciences, 21(6), 1959. https://doi.org/10.3390/ijms21061959