Soluble Epoxide Hydrolase 2 Expression Is Elevated in Obese Humans and Decreased by Physical Activity


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Study Population and the Effects of Exercise
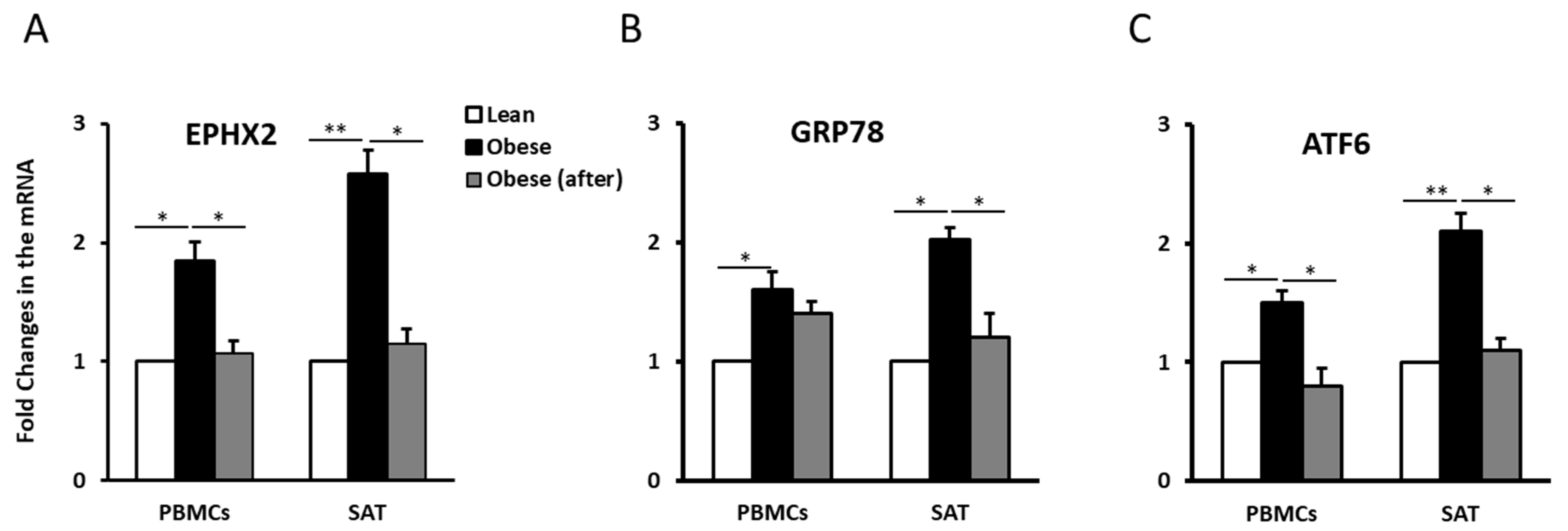
2.2. EPHX2 Expression Levels Are Elevated in the PBMCs and SAT of Obese Subjects and Decrease with Physical Activity
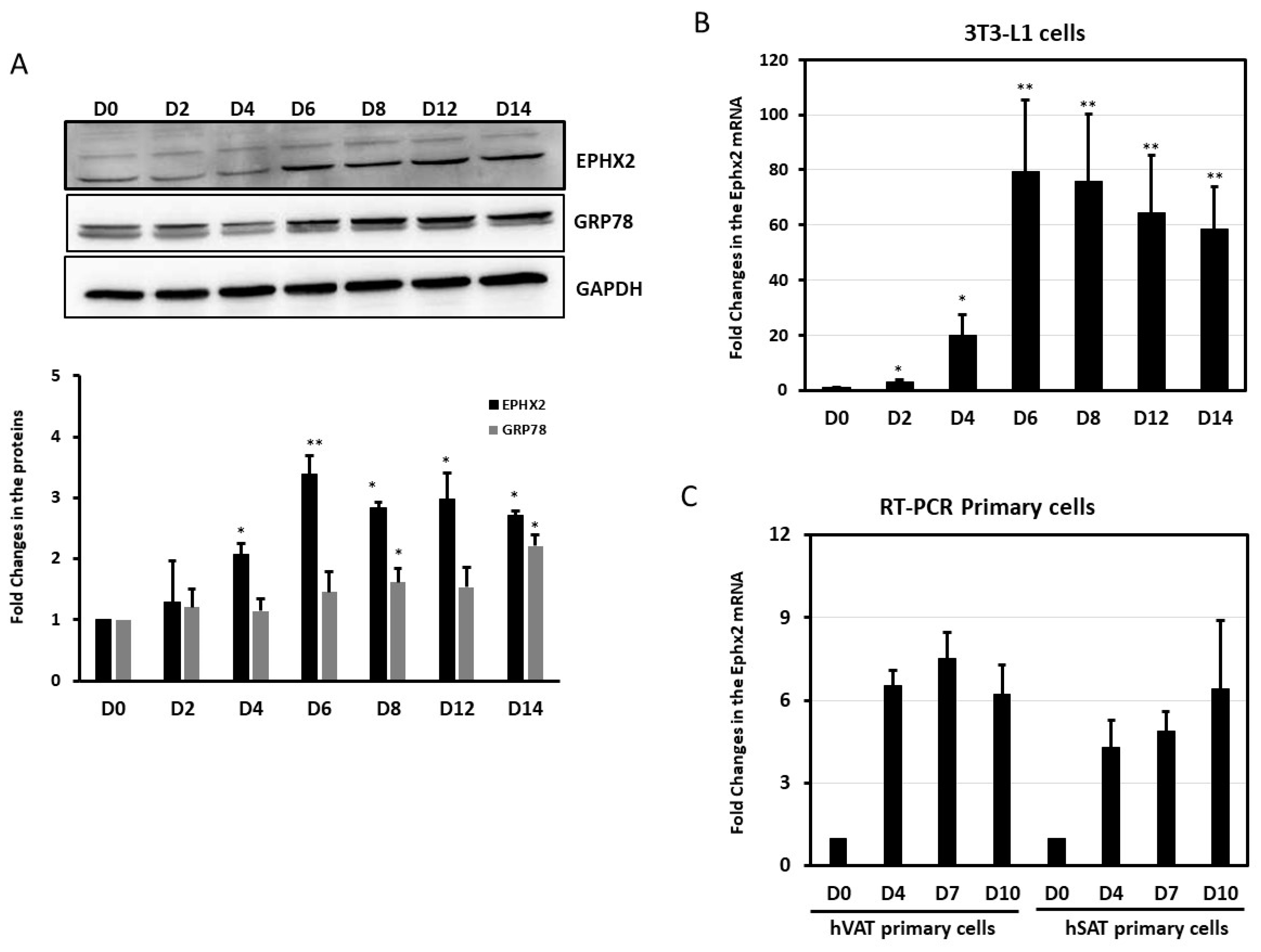
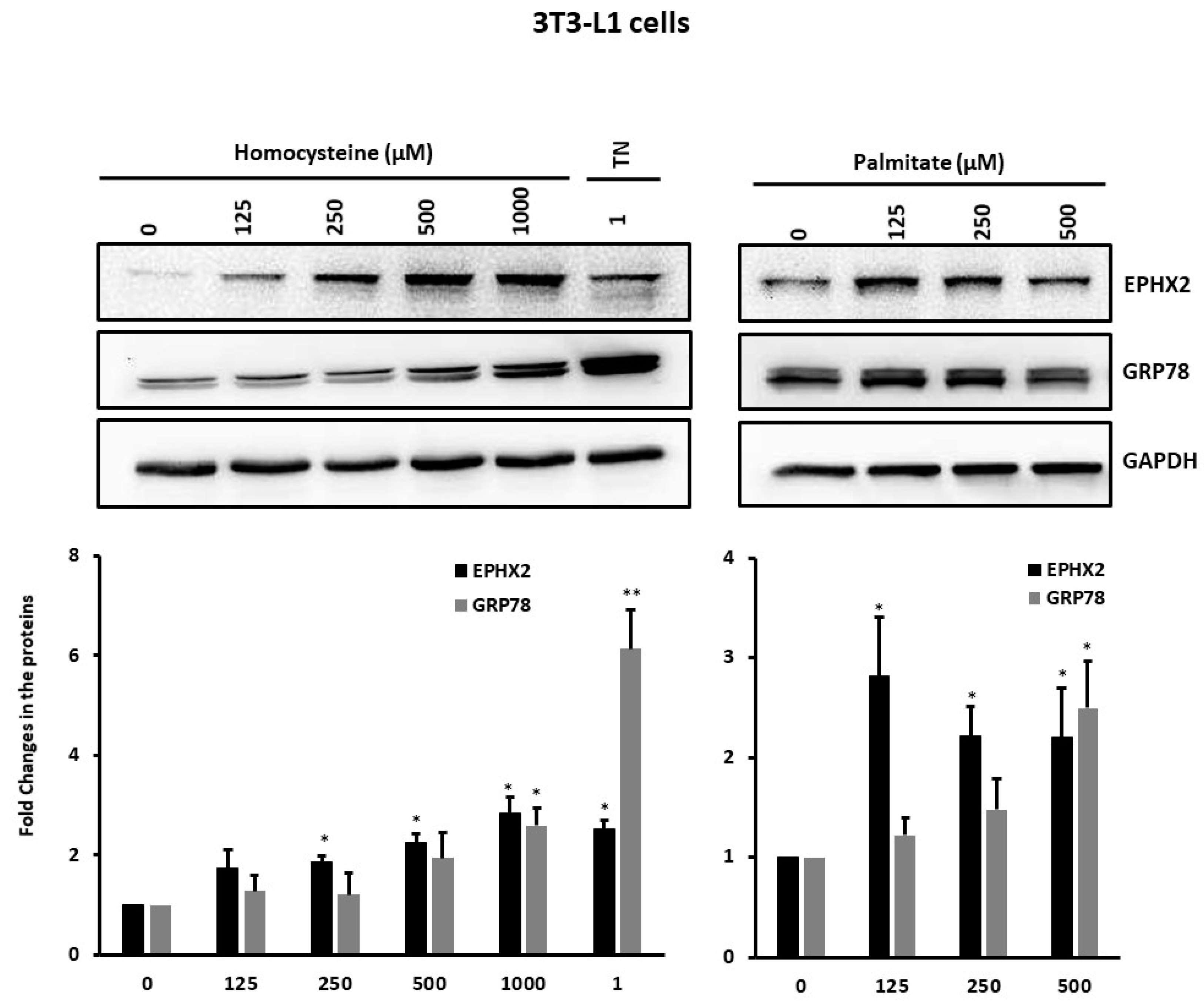
2.3. EPHX2 Expression Levels Increase during Preadipocyte Differentiation
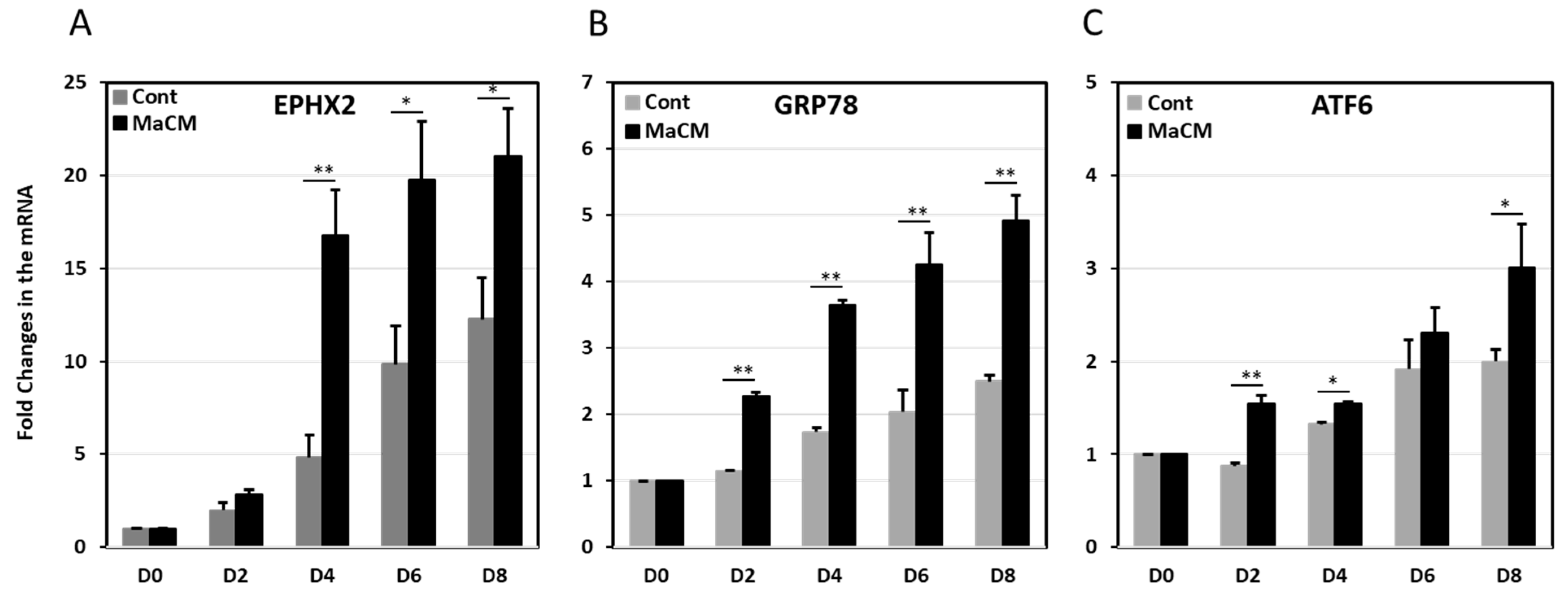
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Exercise Protocol and Anthropometric Measurements
4.3. Blood and Tissue Sampling
4.4. Blood Biochemistry
4.5. Immunohistochemical and Immunofluorescence Analyses
4.6. Cell Cultures
4.7. Quantitative Real-Time PCR
4.8. Western Blot Analysis
4.9. Oil Red O Staining
4.10. AdipoRed Assay
4.11. Statistical Analysis
4.12. Ethics Approval and Consent to Participate
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DBP | diastolic blood pressure |
| EPHX2 | epoxide hydrolase 2 |
| HbA1c | hemoglobin A1c; |
| HDL | high-density lipoprotein |
| HR | heart rate |
| hsCRP | high-sensitivity CRP |
| LDL | low-density lipoprotein; |
| PBF | percent body fat |
| SAT | subcutaneous adipose tissue |
| PBMC | peripheral blood mononuclear cell; |
| sEH | soluble epoxide hydrolase; |
| SBP | systolic blood pressure; |
| TGL | triglyceride |
| VO2,max | maximum oxygen consumption; |
| WC | waist circumference. |
References
- He, J.; Wang, C.; Zhu, Y.; Ai, D. Soluble epoxide hydrolase: A potential target for metabolic diseases. J. Diabetes 2016, 8, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, U.N. Arachidonic acid in health and disease with focus on hypertension and diabetes mellitus: A review. J. Adv. Res. 2018, 11, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.C.; Hammock, B.D. Discovery of inhibitors of soluble epoxide hydrolase: A target with multiple potential therapeutic indications. J. Med. Chem. 2012, 55, 1789–1808. [Google Scholar] [CrossRef] [Green Version]
- Luther, J.M.; Brown, N.J. Epoxyeicosatrienoic acids and glucose homeostasis in mice and men. Prostaglandins Other Lipid Mediat 2016, 125, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.R.; North, K.E.; Bray, M.S.; Fornage, M.; Seubert, J.M.; Newman, J.W.; Hammock, B.D.; Couper, D.J.; Heiss, G.; Zeldin, D.C. Genetic variation in soluble epoxide hydrolase (ephx2) and risk of coronary heart disease: The atherosclerosis risk in communities (aric) study. Hum. Mol Genet. 2006, 15, 1640–1649. [Google Scholar] [CrossRef]
- Hancock, A.M.; Witonsky, D.B.; Gordon, A.S.; Eshel, G.; Pritchard, J.K.; Coop, G.; Di Rienzo, A. Adaptations to climate in candidate genes for common metabolic disorders. PLoS Genet. 2008, 4, e32. [Google Scholar] [CrossRef]
- Yang, L.; Maki-Petaja, K.; Cheriyan, J.; McEniery, C.; Wilkinson, I.B. The role of epoxyeicosatrienoic acids in the cardiovascular system. Br. J. Clin. Pharmacol. 2015, 80, 28–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, C.E.; Shuey, M.M.; Milne, G.L.; Gilbert, K.; Hui, N.; Yu, C.; Luther, J.M.; Brown, N.J. Arg287gln variant of ephx2 and epoxyeicosatrienoic acids are associated with insulin sensitivity in humans. Prostaglandins Other Lipid Mediat 2014, 113-115, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtoshi, K.; Kaneto, H.; Node, K.; Nakamura, Y.; Shiraiwa, T.; Matsuhisa, M.; Yamasaki, Y. Association of soluble epoxide hydrolase gene polymorphism with insulin resistance in type 2 diabetic patients. Biochem. Biophys. Res. Commun. 2005, 331, 347–350. [Google Scholar] [CrossRef]
- Yang, J.; Oh, Y.T.; Wan, D.; Watanabe, R.M.; Hammock, B.D.; Youn, J.H. Postprandial effect to decrease soluble epoxide hydrolase activity: Roles of insulin and gut microbiota. J. Nutr. Biochem. 2017, 49, 8–14. [Google Scholar] [CrossRef]
- Jakicic, J.M.; Rogers, R.J.; Davis, K.K.; Collins, K.A. Role of physical activity and exercise in treating patients with overweight and obesity. Clin. Chem. 2018, 64, 99–107. [Google Scholar] [CrossRef]
- Neufer, P.D.; Bamman, M.M.; Muoio, D.M.; Bouchard, C.; Cooper, D.M.; Goodpaster, B.H.; Booth, F.W.; Kohrt, W.M.; Gerszten, R.E.; Mattson, M.P.; et al. Understanding the cellular and molecular mechanisms of physical activity-induced health benefits. Cell Metab. 2015, 22, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Pucci, B.; Villanova, L.; Sansone, L.; Pellegrini, L.; Tafani, M.; Carpi, A.; Fini, M.; Russo, M.A. Sirtuins: The molecular basis of beneficial effects of physical activity. Intern. Emerg Med. 2013, 8 (Supp. 1), S23–S25. [Google Scholar] [CrossRef]
- Giordano, R.M.; Newman, J.W.; Pedersen, T.L.; Ramos, M.I.; Stebbins, C.L. Effects of dynamic exercise on plasma arachidonic acid epoxides and diols in human volunteers. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 471–479. [Google Scholar] [CrossRef]
- Gollasch, B.; Dogan, I.; Rothe, M.; Gollasch, M.; Luft, F.C. Maximal exercise and plasma cytochrome p450 and lipoxygenase mediators: A lipidomics study. Physiol. Rep. 2019, 7, e14165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellsten, Y.; Nyberg, M.; Jensen, L.G.; Mortensen, S.P. Vasodilator interactions in skeletal muscle blood flow regulation. J. Physiol. 2012, 590, 6297–6305. [Google Scholar] [CrossRef] [PubMed]
- Khadir, A.; Kavalakatt, S.; Madhu, D.; Hammad, M.; Devarajan, S.; Tuomilehto, J.; Tiss, A. Fetuin-a levels are increased in the adipose tissue of diabetic obese humans but not in circulation. Lipids Health Dis. 2018, 17, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Nikolajczyk, B.S. Tissue immune cells fuel obesity-associated inflammation in adipose tissue and beyond. Front. Immunol. 2019, 10, 1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imig, J.D.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases. Nat. Rev. Drug Discov. 2009, 8, 794–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorthioir, A.; Guerrot, D.; Joannides, R.; Bellien, J. Diabetic cvd–soluble epoxide hydrolase as a target. Cardiovasc. Hematol. Agents Med. Chem. 2012, 10, 212–222. [Google Scholar] [CrossRef] [PubMed]
- De Taeye, B.M.; Morisseau, C.; Coyle, J.; Covington, J.W.; Luria, A.; Yang, J.; Murphy, S.B.; Friedman, D.B.; Hammock, B.B.; Vaughan, D.E. Expression and regulation of soluble epoxide hydrolase in adipose tissue. Obesity (Silver Spring) 2010, 18, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khadir, A.; Kavalakatt, S.; Abubaker, J.; Cherian, P.; Madhu, D.; Al-Khairi, I.; Abu-Farha, M.; Warsame, S.; Elkum, N.; Dehbi, M.; et al. Physical exercise alleviates er stress in obese humans through reduction in the expression and release of grp78 chaperone. Metabolism 2016, 65, 1409–1420. [Google Scholar] [CrossRef]
- Sharma, N.K.; Das, S.K.; Mondal, A.K.; Hackney, O.G.; Chu, W.S.; Kern, P.A.; Rasouli, N.; Spencer, H.J.; Yao-Borengasser, A.; Elbein, S.C. Endoplasmic reticulum stress markers are associated with obesity in nondiabetic subjects. J. Clin. Endocrinol. Metab. 2008, 93, 4532–4541. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Duan, X.; Homko, C.; Molina, E.J.; Song, W.; Perez, O.; Cheung, P.; Merali, S. Increase in endoplasmic reticulum stress-related proteins and genes in adipose tissue of obese, insulin-resistant individuals. Diabetes 2008, 57, 2438–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Ye, R.; Jung, D.Y.; Barron, E.; Friedline, R.H.; Benoit, V.M.; Hinton, D.R.; Kim, J.K.; Lee, A.S. Grp78 plays an essential role in adipogenesis and postnatal growth in mice. FASEB J. 2013, 27, 955–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, M.; Spinelli, R.; D’Esposito, V.; Zatterale, F.; Fiory, F.; Nigro, C.; Raciti, G.A.; Miele, C.; Formisano, P.; Beguinot, F.; et al. Pathologic endoplasmic reticulum stress induced by glucotoxic insults inhibits adipocyte differentiation and induces an inflammatory phenotype. Biochim. Biophys. Acta 2016, 1863, 1146–1156. [Google Scholar] [CrossRef]
- Gupta, S.; Deepti, A.; Deegan, S.; Lisbona, F.; Hetz, C.; Samali, A. Hsp72 protects cells from er stress-induced apoptosis via enhancement of ire1alpha-xbp1 signaling through a physical interaction. PLoS Biol. 2010, 8, e1000410. [Google Scholar] [CrossRef] [Green Version]
- Tiss, A.; Khadir, A.; Abubaker, J.; Abu-Farha, M.; Al-Khairi, I.; Cherian, P.; John, J.; Kavalakatt, S.; Warsame, S.; Al-Ghimlas, F.; et al. Immunohistochemical profiling of the heat shock response in obese non-diabetic subjects revealed impaired expression of heat shock proteins in the adipose tissue. Lipids Health Dis. 2014, 13, 106. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Vicario, C.; Alcaraz-Quiles, J.; Garcia-Alonso, V.; Rius, B.; Hwang, S.H.; Titos, E.; Lopategi, A.; Hammock, B.D.; Arroyo, V.; Claria, J. Inhibition of soluble epoxide hydrolase modulates inflammation and autophagy in obese adipose tissue and liver: Role for omega-3 epoxides. Proc. Natl. Acad. Sci. USA 2015, 112, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Zha, W.; Edin, M.L.; Vendrov, K.C.; Schuck, R.N.; Lih, F.B.; Jat, J.L.; Bradbury, J.A.; DeGraff, L.M.; Hua, K.; Tomer, K.B.; et al. Functional characterization of cytochrome p450-derived epoxyeicosatrienoic acids in adipogenesis and obesity. J. Lipid Res. 2014, 55, 2124–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, T.R.; Bettaieb, A.; Kodani, S.; Dong, H.; Myers, R.; Chiamvimonvat, N.; Haj, F.G.; Hammock, B.D. Inhibition of soluble epoxide hydrolase attenuates hepatic fibrosis and endoplasmic reticulum stress induced by carbon tetrachloride in mice. Toxicol. Appl. Pharmacol. 2015, 286, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettaieb, A.; Nagata, N.; AbouBechara, D.; Chahed, S.; Morisseau, C.; Hammock, B.D.; Haj, F.G. Soluble epoxide hydrolase deficiency or inhibition attenuates diet-induced endoplasmic reticulum stress in liver and adipose tissue. J. Biol. Chem. 2013, 288, 14189–14199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Yang, J.; Zhang, J.; Wang, Y.; Hwang, S.H.; Qi, W.; Wan, D.; Kim, D.; Sun, J.; Sanidad, K.Z.; et al. Lipidomic profiling reveals soluble epoxide hydrolase as a therapeutic target of obesity-induced colonic inflammation. Proc. Natl. Acad. Sci. USA 2018, 115, 5283–5288. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.; Sun, D. Sexually dimorphic regulation of eet synthesis and metabolism: Roles of estrogen. Front. Pharmacol. 2018, 9, 1222. [Google Scholar] [CrossRef]
- Cao, J.; Singh, S.P.; McClung, J.A.; Joseph, G.; Vanella, L.; Barbagallo, I.; Jiang, H.; Falck, J.R.; Arad, M.; Shapiro, J.I.; et al. Eet intervention on wnt1, nov, and ho-1 signaling prevents obesity-induced cardiomyopathy in obese mice. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H368–H380. [Google Scholar] [CrossRef] [Green Version]
- do Carmo, J.M.; da Silva, A.A.; Morgan, J.; Jim Wang, Y.X.; Munusamy, S.; Hall, J.E. Inhibition of soluble epoxide hydrolase reduces food intake and increases metabolic rate in obese mice. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 598–604. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Luo, F.; Zhang, X.; Chen, J.; Shen, L.; Zhu, Y.; Xu, D. Tppu enhanced exercise-induced epoxyeicosatrienoic acid concentrations to exert cardioprotection in mice after myocardial infarction. J. Cell Mol. Med. 2018, 22, 1489–1500. [Google Scholar] [CrossRef] [Green Version]
- Chadderdon, S.M.; Belcik, J.T.; Smith, E.; Pranger, L.; Kievit, P.; Grove, K.L.; Lindner, J.R. Activity restriction, impaired capillary function, and the development of insulin resistance in lean primates. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E607–E613. [Google Scholar] [CrossRef]
- Stanford, K.I.; Lynes, M.D.; Takahashi, H.; Baer, L.A.; Arts, P.J.; May, F.J.; Lehnig, A.C.; Middelbeek, R.J.W.; Richard, J.J.; So, K.; et al. 12,13-dihome: An exercise-induced lipokine that increases skeletal muscle fatty acid uptake. Cell Metab. 2018, 27, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Blaak, E.E.; van Aggel-Leijssen, D.P.; Wagenmakers, A.J.; Saris, W.H.; van Baak, M.A. Impaired oxidation of plasma-derived fatty acids in type 2 diabetic subjects during moderate-intensity exercise. Diabetes 2000, 49, 2102–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khadir, A.; Kavalakatt, S.; Cherian, P.; Warsame, S.; Abubaker, J.A.; Dehbi, M.; Tiss, A. Physical exercise enhanced heat shock protein 60 expression and attenuated inflammation in the adipose tissue of human diabetic obese. Front. Endocrinol. (Lausanne) 2018, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Kavalakatt, S.; Khadir, A.; Madhu, D.; Hammad, M.; Devarajan, S.; Abubaker, J.; Al-Mulla, F.; Tuomilehto, J.; Tiss, A. Urocortin 3 levels are impaired in overweight humans with and without type 2 diabetes and modulated by exercise. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khadir, A.; Tiss, A.; Abubaker, J.; Abu-Farha, M.; Al-Khairi, I.; Cherian, P.; John, J.; Kavalakatt, S.; Warsame, S.; Al-Madhoun, A.; et al. Map kinase phosphatase dusp1 is overexpressed in obese humans and modulated by physical exercise. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E71–E83. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Lean (n = 20) (Mean ± SD, n = 20) | Obese (n = 20) (Mean ± SD, n = 20) | (p-Value) |
|---|---|---|---|
| Anthropometric and physical characteristics | |||
| AGE (year) | 38.75 ± 9.60 | 40.13 ± 8.68 | 0.55 |
| Gender (M/F) | 8/12 | 10/10 | 0.37 |
| BMI (kg/m2) | 22.72 ± 2.09 | 34.83 ± 2.99 | <0.001 |
| Waist (cm) | 79.45 ± 15.96 | 108.52 ± 13.71 | <0.001 |
| Hip (cm) | 92.07 ± 14.98 | 118.15 ± 8.29 | <0.001 |
| PBF (%) | 27.50 ± 5.35 | 39.36 ± 5.12 | <0.001 |
| SBP (mmHg) | 113.00 ± 10.81 | 127.50 ± 11.89 | 0.01 |
| DBP (mmHg) | 76.43 ± 6.33 | 82.00 ± 10.14 | 0.13 |
| VO2, Max (mL/kg/min) | 21.63 ± 3.76 | 16.54 ± 4.83 | 0.03 |
| Metabolic markers | |||
| Cholesterol (mmol/L) | 5.22 ± 0.91 | 5.38 ± 1.10 | 0.74 |
| HDL (mmol/L) | 1.48 ± 0.53 | 1.19 ± 0.26 | 0.015 |
| LDL (mmol/L) | 3.23 ± 0.91 | 3.39 ± 0.98 | 0.28 |
| TG (mmol/L) | 0.91 ± 0.42 | 1.47 ± 0.83 | 0.008 |
| Glucose (mmol/L) | 5.0 ± 0.5 | 5.7 ± 0.7 | 0.013 |
| HbA1c (%) | 5.50 ± 0.44 | 5.93 ± 0.48 | 0.014 |
| Insulin (ng/mL) | 2.6 ± 1.3 | 4.2 ± 2.5 | 0.029 |
| HOMA-IR | 0.7 ± 0.5 | 1.1 ± 0.7 | 0.025 |
| Inflammatory markers | |||
| IL-6 (pg/mL) | 17.06 ± 2.9 | 17.40 ± 4.4 | 0.39 |
| TNF-a (pg/mL) | 123.52 ± 35 | 126.47 ± 43 | 0.94 |
| hsCRP (μg/mL) | 1.7 ± 1.2 | 5.3 ± 3.6 | <0.001 |
| Stress markers | |||
| GRP78 (μg/mL) | 0.925 ± 0.95 | 1.2 ± 0.75 | 0.012 |
| HSP72 (ng/mL) | 1.85± 0.53 | 1.62 ± 0.45 | 0.055 |
| Marker | Obese before (Mean ± SD, n = 20) | Obese after (Mean ± SD, n = 20) | p-Value |
|---|---|---|---|
| Anthropometric and physical characteristics | |||
| BMI (kg/m2) | 34.83 ± 2.99 | 34.08 ± 3.31 | 0.09 |
| Waist (cm) | 108.52 ± 13.71 | 105.17 ± 10.97 | 0.03 |
| Hip (cm) | 118.15 ± 8.29 | 116.57 ± 9.05 | 0.23 |
| PBF (%) | 39.36 ± 5.12 | 38.44 ± 5.09 | 0.02 |
| SBP (mmHg) | 127.50 ± 11.89 | 117.22 ± 8.20 | 0.01 |
| DBP (mmHg) | 82.00 ± 10.14 | 79.39 ± 4.95 | 0.08 |
| VO2,max (mL/kg/min) | 16.54 ± 4.83 | 18.94 ± 3.78 | 0.01 |
| Metabolic markers | |||
| Cholesterol (mmol/L) | 5.38 ± 1.10 | 5.37 ± 1.22 | 0.83 |
| HDL (mmol/L) | 1.19 ± 0.26 | 1.29 ± 0.49 | 0.11 |
| LDL (mmol/L) | 3.39 ± 0.98 | 3.47 ± 1.13 | 0.69 |
| TG (mmol/L) | 1.47 ± 0.83 | 1.28 ± 0.77 | 0.17 |
| Glucose (mmol/L) | 5.7 ± 0.7 | 5.5 ± 0.6 | 0.49 |
| HbA1c (%) | 5.93 ± 0.48 | 5.78 ± 0.42 | 0.28 |
| Insulin (ng/mL) | 4.2 ± 2.5 | 3.30 ± 1.11 | 0.01 |
| HOMA-IR | 1.1 ± 0.7 | 0.8 ± 0.2 | 0.04 |
| Inflammatory markers | |||
| IL-6 (pg/mL) | 17.4 ± 4.4 | 16.6 ± 6.5 | 0.87 |
| TNF-a (pg/mL) | 126.4 ± 43 | 123.8± 33.1 | 0.59 |
| hsCRP (μg/mL) | 5.3 ± 3.6 | 6.2 ± 4.1 | 0.43 |
| Stress markers | |||
| GRP78 (μg/mL) | 1.2 ± 0.75 | 0.85 ± 0.26 | 0.02 |
| HSP72 (ng/mL) | 1.62± 0.45 | 2.08 ± 0.59 | 0.35 |
| Markers | R-Value | p-Value |
|---|---|---|
| BMI (kg/m2) | 0.73 | 0.001 |
| Waist (cm) | 0.66 | 0.008 |
| Hip (cm) | 0.51 | 0.004 |
| PBF (%) | 0.54 | 0.002 |
| SBP (mmHg) | 0.49 | 0.006 |
| DBP (mmHg) | 0.30 | 0.110 |
| VO2, Max (mL/kg/min) | –0.44 | 0.016 |
| HSP72 | 0.43 | 0.020 |
| GRP78 | 0.52 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khadir, A.; Kavalakatt, S.; Madhu, D.; Cherian, P.; Al-Mulla, F.; Abubaker, J.; Tiss, A. Soluble Epoxide Hydrolase 2 Expression Is Elevated in Obese Humans and Decreased by Physical Activity. Int. J. Mol. Sci. 2020, 21, 2056. https://doi.org/10.3390/ijms21062056
Khadir A, Kavalakatt S, Madhu D, Cherian P, Al-Mulla F, Abubaker J, Tiss A. Soluble Epoxide Hydrolase 2 Expression Is Elevated in Obese Humans and Decreased by Physical Activity. International Journal of Molecular Sciences. 2020; 21(6):2056. https://doi.org/10.3390/ijms21062056
Chicago/Turabian StyleKhadir, Abdelkrim, Sina Kavalakatt, Dhanya Madhu, Preethi Cherian, Fahd Al-Mulla, Jehad Abubaker, and Ali Tiss. 2020. "Soluble Epoxide Hydrolase 2 Expression Is Elevated in Obese Humans and Decreased by Physical Activity" International Journal of Molecular Sciences 21, no. 6: 2056. https://doi.org/10.3390/ijms21062056
APA StyleKhadir, A., Kavalakatt, S., Madhu, D., Cherian, P., Al-Mulla, F., Abubaker, J., & Tiss, A. (2020). Soluble Epoxide Hydrolase 2 Expression Is Elevated in Obese Humans and Decreased by Physical Activity. International Journal of Molecular Sciences, 21(6), 2056. https://doi.org/10.3390/ijms21062056