Adrenoceptor Expression during Intervertebral Disc Degeneration

 , ,
, ,
Abstract
:
1. Introduction
2. Results
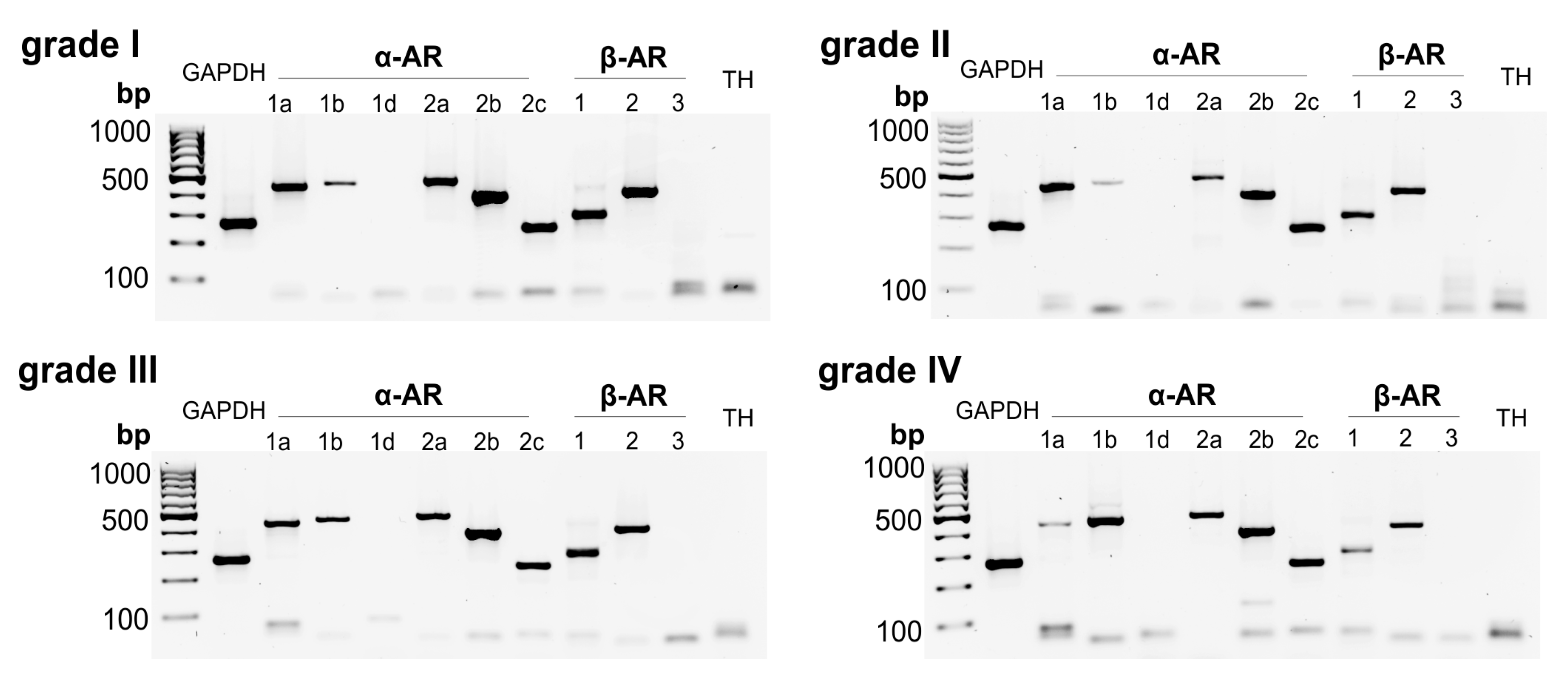
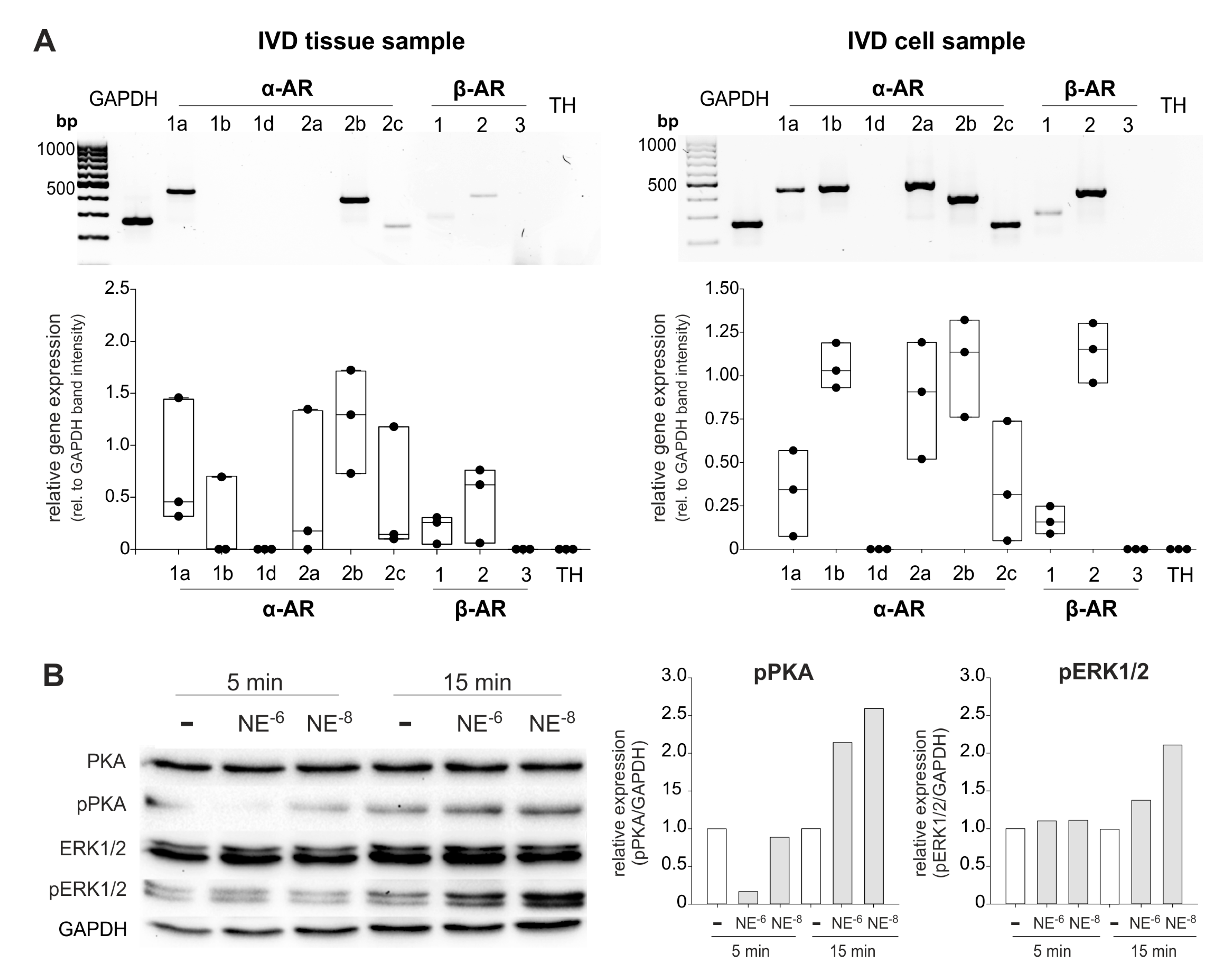
2.1. AR and TH Gene Expression in Human IVD Tissue Samples
2.2. Correlation between AR and TH Gene Expression and the Degree of IVD Degeneration
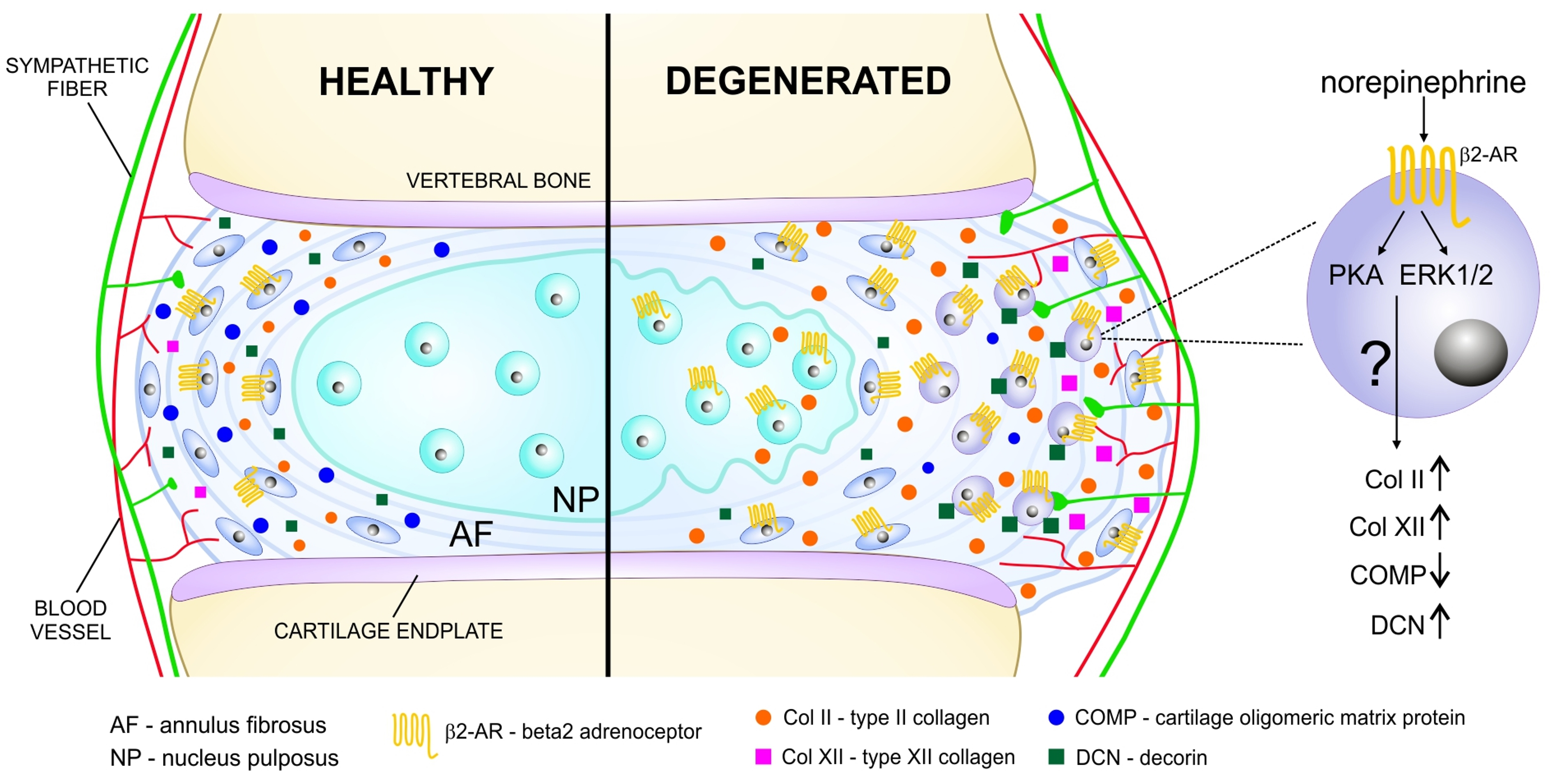
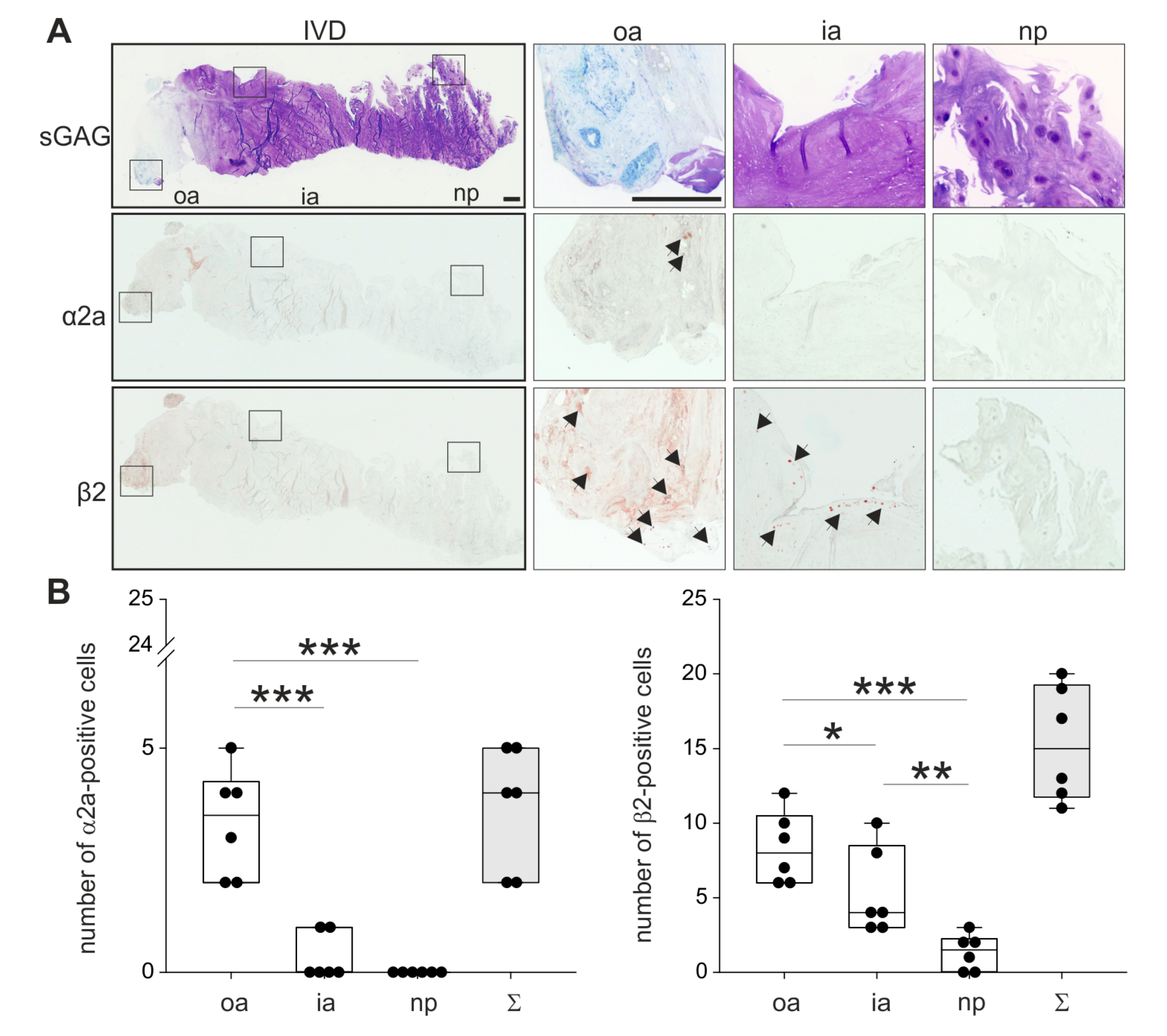
2.3. Localization of ARs in Human Degenerated IVD Tissue
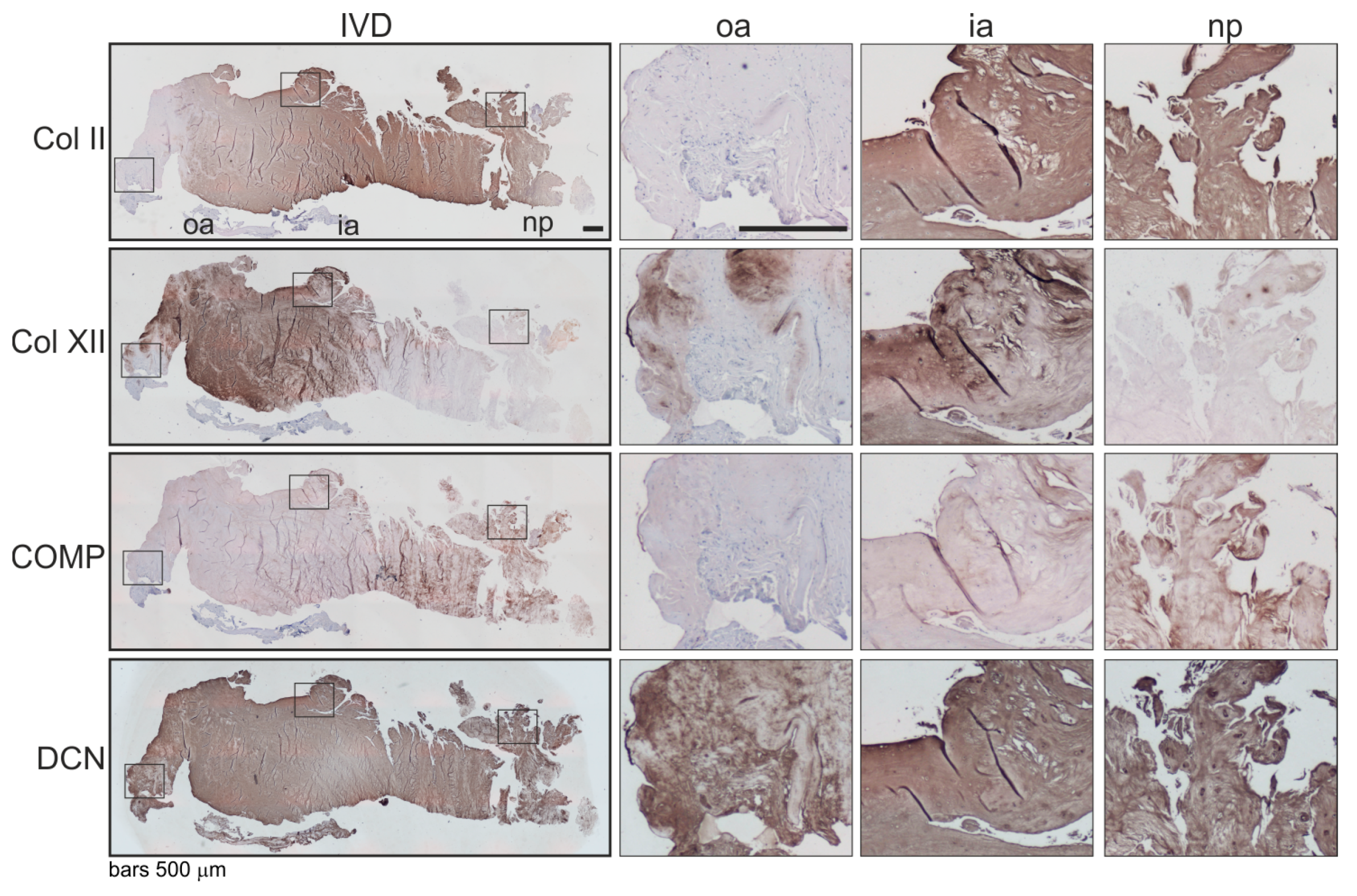
2.4. Associations of β2-AR Expression with Changes in ECM Expression in Human IVD Tissue
2.5. AR Protein Expression in Murine IVD Degeneration Models and Associations of ar Expression with Changes in ECM Expression in Murine IVD Tissue
2.6. AR and TH Gene Expression in Isolated Human IVD Cells and IVD Cell Response to NE
3. Discussion
4. Materials and Methods
4.1. Human IVD Tissue
4.2. Murine IVD Samples
4.3. Adrenoceptor Gene Expression Analysis
4.4. Immunohistological Stainings
4.5. β2-AR Western Blot
4.6. IVD Cell Isolation and Stimulation with NE
4.7. NE-Dependent Signal Transduction
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | Annulus fibrosus |
| AR | Adrenoceptor |
| cDNA | Complementary desoxy ribonucleic acid |
| Col II | Type II collagen |
| Col VI | Type VI collagen |
| Col XII | Type XII collagen |
| COMP | Cartilage oligomeric matrix protein |
| DCN | Decorin |
| DMEM/F12 | Dulbecco’s modified eagle’s medium and Ham’s F-12 Medium |
| DMMB | 1,9-Dimethyl-methylene blue |
| ECM | Extracellular matrix |
| ERK | Extracellular signal-regulated kinases |
| FBS | Fetal bovine serum |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| HT29 | Human colon cancer cell line |
| IL | Interleukin |
| IVD | Intervertebral disc |
| IVDD | Intervertebral disc degeneration |
| mRNA | Messenger ribonucleic acid |
| NE | Norepinephrine |
| NP | Nucleus pulposus |
| OA | Osteoarthritis |
| P/S | Penicillin streptomycin |
| PCR | Polymerase chain reaction |
| pERK | Phosphorylated extracellular signal-regulated kinases |
| PKA | Protein kinase A |
| pPKA | Phosphorylated protein kinase A |
| RT-PCR | Reverse-transcriptase PCR |
| Saf-O | Safranin-O/fast green staining |
| sGAG | Sulphated glycosaminoglycans |
| TH | Tyrosine hydroxylase |
| WT | Wildtype mice |
References
- Taher, F.; Essig, D.; Lebl, D.R.; Hughes, A.P.; Sama, A.A.; Cammisa, F.P.; Girardi, F.P. Lumbar degenerative disc disease: Current and future concepts of diagnosis and management. Adv. Orthop. 2012, 2012, 970752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Burden of Disease Study 2013 Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990-2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 386, 743–800. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.J.; Nerurkar, N.L.; Choi, K.S.; Harfe, B.D.; Elliott, D.M. Degeneration and regeneration of the intervertebral disc: Lessons from development. Dis. Models Mech. 2011, 4, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustenburg, C.M.E.; Emanuel, K.S.; Peeters, M.; Lems, W.F.; Vergroesen, P.A.; Smit, T.H. Osteoarthritis and intervertebral disc degeneration: Quite different, quite similar. Jor Spine 2018, 1, e1033. [Google Scholar] [CrossRef] [PubMed]
- de Kunder, S.L.; Rijkers, K.; Caelers, I.; de Bie, R.A.; Koehler, P.J.; van Santbrink, H. Lumbar Interbody Fusion: A Historical Overview and a Future Perspective. Spine 2018, 43, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewski, K.A.; Saganiak, K.; Gladysz, T.; Walocha, J.A. The biology behind the human intervertebral disc and its endplates. Folia Morphol. 2015, 74, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Kerr, G.J.; Veras, M.A.; Kim, M.K.; Seguin, C.A. Decoding the intervertebral disc: Unravelling the complexities of cell phenotypes and pathways associated with degeneration and mechanotransduction. Semin. Cell Dev. Biol. 2017, 62, 94–103. [Google Scholar] [CrossRef]
- Kepler, C.K.; Ponnappan, R.K.; Tannoury, C.A.; Risbud, M.V.; Anderson, D.G. The molecular basis of intervertebral disc degeneration. Spine J. 2013, 13, 318–330. [Google Scholar] [CrossRef]
- Janeczko, L.; Janeczko, M.; Chrzanowski, R.; Zielinski, G. The role of polymorphisms of genes encoding collagen IX and XI in lumbar disc disease. Neurol. I Neurochir. Pol. 2014, 48, 60–62. [Google Scholar] [CrossRef]
- Martirosyan, N.L.; Patel, A.A.; Carotenuto, A.; Kalani, M.Y.; Belykh, E.; Walker, C.T.; Preul, M.C.; Theodore, N. Genetic Alterations in Intervertebral Disc Disease. Front. Surg. 2016, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Grässel, S.G. The role of peripheral nerve fibers and their neurotransmitters in cartilage and bone physiology and pathophysiology. Arthritis Res. 2014, 16, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagatsu, T.; Levitt, M.; Udenfriend, S. TYROSINE HYDROXYLASE. THE INITIAL STEP IN NOREPINEPHRINE BIOSYNTHESIS. J. Biol. Chem. 1964, 239, 2910–2917. [Google Scholar] [PubMed]
- Rosenbaum, D.M.; Rasmussen, S.G.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Cosamalon, J.; del Valle, M.E.; Calavia, M.G.; Garcia-Suarez, O.; Lopez-Muniz, A.; Otero, J.; Vega, J.A. Intervertebral disc, sensory nerves and neurotrophins: Who is who in discogenic pain? J. Anat. 2010, 217, 1–15. [Google Scholar] [CrossRef]
- Pertovaara, A. The noradrenergic pain regulation system: A potential target for pain therapy. Eur. J. Pharmacol. 2013, 716, 2–7. [Google Scholar] [CrossRef]
- Binch, A.L.; Cole, A.A.; Breakwell, L.M.; Michael, A.L.; Chiverton, N.; Creemers, L.B.; Cross, A.K.; Le Maitre, C.L. Nerves are more abundant than blood vessels in the degenerate human intervertebral disc. Arthritis Res 2015, 17, 370. [Google Scholar] [CrossRef] [Green Version]
- Barczewska, M.; Juranek, J.; Wojtkiewicz, J. Origins and Neurochemical Characteristics of Porcine Intervertebral Disc Sympathetic Innervation: A Preliminary Report. J. Mol. Neurosci. 2017, 63, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Jenei-Lanzl, Z.; Grassel, S.; Pongratz, G.; Kees, F.; Miosge, N.; Angele, P.; Straub, R.H. Norepinephrine inhibition of mesenchymal stem cell and chondrogenic progenitor cell chondrogenesis and acceleration of chondrogenic hypertrophy. Arthritis Rheumatol. 2014, 66, 2472–2481. [Google Scholar] [CrossRef] [Green Version]
- Gotz, W.; Barnert, S.; Bertagnoli, R.; Miosge, N.; Kresse, H.; Herken, R. Immunohistochemical localization of the small proteoglycans decorin and biglycan in human intervertebral discs. Cell Tissue Res. 1997, 289, 185–190. [Google Scholar] [CrossRef]
- Ishii, Y.; Thomas, A.O.; Guo, X.E.; Hung, C.T.; Chen, F.H. Localization and distribution of cartilage oligomeric matrix protein in the rat intervertebral disc. Spine 2006, 31, 1539–1546. [Google Scholar] [CrossRef]
- Melrose, J.; Ghosh, P.; Taylor, T.K. A comparative analysis of the differential spatial and temporal distributions of the large (aggrecan, versican) and small (decorin, biglycan, fibromodulin) proteoglycans of the intervertebral disc. J. Anat. 2001, 198, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Klawitter, M.; Hakozaki, M.; Kobayashi, H.; Krupkova, O.; Quero, L.; Ospelt, C.; Gay, S.; Hausmann, O.; Liebscher, T.; Meier, U.; et al. Expression and regulation of toll-like receptors (TLRs) in human intervertebral disc cells. Eur. Spine J. 2014, 23, 1878–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xiong, C.; Kudelko, M.; Li, Y.; Wang, C.; Wong, Y.L.; Tam, V.; Rai, M.F.; Cheverud, J.; Lawson, H.A.; et al. Early onset of disc degeneration in SM/J mice is associated with changes in ion transport systems and fibrotic events. Matrix Biol. 2018, 70, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, J.; Schafer, N.; Bauer, R.; Jenei-Lanzl, Z.; Springorum, R.H.; Grassel, S. Norepinephrine modulates osteoarthritic chondrocyte metabolism and inflammatory responses. Osteoarthr. Cartil. 2016, 24, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.R.; Sakai, D.; Nakai, T.; Toyama, K.; Mochida, J.; Alini, M.; Grad, S. A phenotypic comparison of intervertebral disc and articular cartilage cells in the rat. Eur. Spine J. 2007, 16, 2174–2185. [Google Scholar] [CrossRef] [Green Version]
- Hadcock, J.R.; Malbon, C.C. Down-regulation of beta-adrenergic receptors: Agonist-induced reduction in receptor mRNA levels. Proc. Natl. Acad. Sci. USA 1988, 85, 5021–5025. [Google Scholar] [CrossRef] [Green Version]
- Heck, D.A.; Bylund, D.B. Mechanism of down-regulation of alpha-2 adrenergic receptor subtypes. J. Pharmacol. Exp. Ther. 1997, 282, 1219–1227. [Google Scholar]
- Wikberg, J.E.; Akers, M.; Caron, M.G.; Hagen, P.O. Norepinephrine-induced down regulation of alpha 1 adrenergic receptors in cultured rabbit aorta smooth muscle cells. Life Sci. 1983, 33, 1409–1417. [Google Scholar] [CrossRef]
- Neidlinger-Wilke, C.; Galbusera, F.; Pratsinis, H.; Mavrogonatou, E.; Mietsch, A.; Kletsas, D.; Wilke, H.J. Mechanical loading of the intervertebral disc: From the macroscopic to the cellular level. Eur. Spine J. 2014, 23 (Suppl. 3), S333–S343. [Google Scholar] [CrossRef]
- Vergroesen, P.P.; Kingma, I.; Emanuel, K.S.; Hoogendoorn, R.J.; Welting, T.J.; van Royen, B.J.; van Dieen, J.H.; Smit, T.H. Mechanics and biology in intervertebral disc degeneration: A vicious circle. Osteoarthr. Cartil. 2015, 23, 1057–1070. [Google Scholar] [CrossRef] [Green Version]
- Mederos y Schnitzler, M.; Storch, U.; Meibers, S.; Nurwakagari, P.; Breit, A.; Essin, K.; Gollasch, M.; Gudermann, T. Gq-coupled receptors as mechanosensors mediating myogenic vasoconstriction. Embo J. 2008, 27, 3092–3103. [Google Scholar] [CrossRef] [PubMed]
- Storch, U.; Mederos y Schnitzler, M.; Gudermann, T. G protein-mediated stretch reception. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1241–H1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldeira, J.; Santa, C.; Osorio, H.; Molinos, M.; Manadas, B.; Goncalves, R.; Barbosa, M. Matrisome Profiling During Intervertebral Disc Development And Ageing. Sci. Rep. 2017, 7, 11629. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.W.; Ahmed, N.; Parreno, J.; Lunstrum, G.P.; Gross, A.E.; Diamandis, E.P.; Kandel, R.A. Collagen type XII and versican are present in the early stages of cartilage tissue formation by both redifferentating passaged and primary chondrocytes. Tissue Eng. Part A 2015, 21, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.E.; Keene, D.R.; Tufa, S.F.; Lunstrum, G.P.; Morris, N.P. Developmental distribution of collagen type XII in cartilage: Association with articular cartilage and the growth plate. J. Bone Miner. Res. 2001, 16, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A. Osteoarthritis year 2012 in review: Biomarkers. Osteoarthr. Cartil. 2012, 20, 1451–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto, G.; Soininen, A.; Ylinen, P.; Sandelin, J.; Konttinen, Y.T.; Nordstrom, D.C.; Eklund, K.K. Soluble biglycan: A potential mediator of cartilage degradation in osteoarthritis. Arthritis Res. 2015, 17, 379. [Google Scholar] [CrossRef] [Green Version]
- Zhen, E.Y.; Brittain, I.J.; Laska, D.A.; Mitchell, P.G.; Sumer, E.U.; Karsdal, M.A.; Duffin, K.L. Characterization of metalloprotease cleavage products of human articular cartilage. Arthritis Rheum. 2008, 58, 2420–2431. [Google Scholar] [CrossRef]
- Guo, D.; Kassiri, Z.; Basu, R.; Chow, F.L.; Kandalam, V.; Damilano, F.; Liang, W.; Izumo, S.; Hirsch, E.; Penninger, J.M.; et al. Loss of PI3Kgamma enhances cAMP-dependent MMP remodeling of the myocardial N-cadherin adhesion complexes and extracellular matrix in response to early biomechanical stress. Circ. Res. 2010, 107, 1275–1289. [Google Scholar] [CrossRef]
- Speichert, S.; Molotkov, N.; El Bagdadi, K.; Meurer, A.; Zaucke, F.; Jenei-Lanzl, Z. Role of Norepinephrine in IL-1beta-Induced Chondrocyte Dedifferentiation under Physioxia. Int. J. Mol. Sci. 2019, 20. [Google Scholar]
- Schubert, A.K.; Smink, J.J.; Pumberger, M.; Putzier, M.; Sittinger, M.; Ringe, J. Standardisation of basal medium for reproducible culture of human annulus fibrosus and nucleus pulposus cells. J. Orthop. Surg. Res. 2018, 13, 209. [Google Scholar] [CrossRef] [PubMed]
- Jiao, K.; Zeng, G.; Niu, L.N.; Yang, H.X.; Ren, G.T.; Xu, X.Y.; Li, F.F.; Tay, F.R.; Wang, M.Q. Activation of alpha2A-adrenergic signal transduction in chondrocytes promotes degenerative remodelling of temporomandibular joint. Sci. Rep. 2016, 6, 30085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Bagdadi, K.; Zaucke, F.; Meurer, A.; Straub, R.H.; Jenei-Lanzl, Z. Norepinephrine Inhibits Synovial Adipose Stem Cell Chondrogenesis via alpha2a-Adrenoceptor-Mediated ERK1/2 Activation. Int. J. Mol. Sci. 2019, 20, 3127. [Google Scholar] [CrossRef] [Green Version]
- Tiaden, A.N.; Klawitter, M.; Lux, V.; Mirsaidi, A.; Bahrenberg, G.; Glanz, S.; Quero, L.; Liebscher, T.; Wuertz, K.; Ehrmann, M.; et al. Detrimental role for human high temperature requirement serine protease A1 (HTRA1) in the pathogenesis of intervertebral disc (IVD) degeneration. J. Biol. Chem. 2012, 287, 21335–21345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, P.; Zwolanek, D.; Keene, D.R.; Schulz, J.N.; Blumbach, K.; Heinegard, D.; Zaucke, F.; Paulsson, M.; Krieg, T.; Koch, M.; et al. Collagen XII and XIV, new partners of cartilage oligomeric matrix protein in the skin extracellular matrix suprastructure. J. Biol. Chem. 2012, 287, 22549–22559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitznagel, L.; Nitsche, D.P.; Paulsson, M.; Maurer, P.; Zaucke, F. Characterization of a pseudoachondroplasia-associated mutation (His587-->Arg) in the C-terminal, collagen-binding domain of cartilage oligomeric matrix protein (COMP). Biochem. J. 2004, 377, 479–487. [Google Scholar] [CrossRef] [Green Version]
- 47. Mayorca-Guiliani Willacy, O.; Madsen, C.D.; Rafaeva, M.; Heumüller, S.E.; Bock, F.; Sengle, G.; Koch, M.; Imhof, T.; Zaucke, F.; Wagener, R.; et al. Decellularization and Antibody Staining of Mouse Tissues to Map Native Extracellular Matrix Structures in 3D. Nature Protocols. 2019, 14, 3395–3425. [Google Scholar] [CrossRef]
- Tang, X.; Richardson, W.J.; Fitch, R.D.; Brown, C.R.; Isaacs, R.E.; Chen, J. A new non-enzymatic method for isolating human intervertebral disc cells preserves the phenotype of nucleus pulposus cells. Cytotechnology 2014, 66, 979–986. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AR Subtype | Total (n = 43) | Grade I (n = 5) | Grade II (n = 9) | Grade III (n = 21) | Grade IV (n = 8) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | n | % | n | % | |
| α1a | 42 | 97.7% | 5 | 100% | 8 | 88.9% | 21 | 100% | 8 | 100% |
| α1b | 20 | 46.5% | 2 | 40% | 5 | 55.6% | 8 | 38.1% | 5 | 62.5% |
| α2a | 39 | 90.7% | 4 | 80% | 7 | 77.8% | 21 | 100% | 7 | 87.5% |
| α2b | 42 | 97.7% | 4 | 80% | 9 | 100% | 21 | 100% | 8 | 100% |
| α2c | 42 | 97.7% | 5 | 100% | 9 | 100% | 20 | 95.2% | 8 | 100% |
| β1 | 43 | 100% | 5 | 100% | 9 | 100% | 21 | 100% | 8 | 100% |
| β2 | 43 | 100% | 5 | 100% | 9 | 100% | 21 | 100% | 8 | 100% |
| Patient Characteristics | Number (%)/Mean Age ± SEM |
|---|---|
| total (number/age) | 43 (100%)/66.93 ± 1.83 |
| female (number/age) | 33 (76.74%)/66.64 ± 2.21 |
| male (number/age) | 10 (23.26%)/67.9 ± 2.9 |
| Gene Symbol | NCBI Reference | Foward (5′−3’) | Reverse (5´−3´) |
|---|---|---|---|
| GAPDH | NM_001289745.2 | CTCCTGTTCGACAGTCAGCC | TTCCCGTTCTCAGCCTTGAC |
| ADRA1A | NM_000680.3 | CCATGCTCCAGCCAAGAGTT | TCCTGTCCTAGACTTCCTCCC |
| ADRA1B | NM_000679.3 | GTCCACCGTCATCTCCATCG | GAACAAGGAGCCAAGCGGTAG |
| ADRA1D | NM_000678.3 | TGACTTTCCGCGATCTCCTG | TTACCTGCCACGGCCATAAG |
| ADRA2A | NM_000681.3 | TGGTCATCGGAGTGTTCGTG | GCCCACTAGGAAGATGGCTC |
| ADRA2B | NM_000682.6 | GACATTTCACCGGCAACACC | GGGACTGAGAACCAGGAAGC |
| ADRA2C | NM000683.3 | CGATGTGCTGTTTTGCACCT | GGATGTACCAGGTCTCGTCG |
| ADRB1 | NM_000684.2 | TAGCAGGTGAACTCGAAGCC | ATCTTCCACTCCGGTCCTCT |
| ADRB2 | NM_000024.5 | CAGAGCCTGCTGACCAAGAA | GCCTAACGTCTTGAGGGCTT |
| ADRB3 | NM_000025.2 | GCCAATTCTGCCTTCAACCC | GCCAGAGGTTTTCCACAGGT |
| TH | NM_000360.3 | CAGGCAGAGGCCATCATGT | GTGGTCCAAGTCCAGGTCAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kupka, J.; Kohler, A.; El Bagdadi, K.; Bostelmann, R.; Brenneis, M.; Fleege, C.; Chan, D.; Zaucke, F.; Meurer, A.; Rickert, M.; et al. Adrenoceptor Expression during Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2020, 21, 2085. https://doi.org/10.3390/ijms21062085
Kupka J, Kohler A, El Bagdadi K, Bostelmann R, Brenneis M, Fleege C, Chan D, Zaucke F, Meurer A, Rickert M, et al. Adrenoceptor Expression during Intervertebral Disc Degeneration. International Journal of Molecular Sciences. 2020; 21(6):2085. https://doi.org/10.3390/ijms21062085
Chicago/Turabian StyleKupka, Johannes, Annika Kohler, Karima El Bagdadi, Richard Bostelmann, Marco Brenneis, Christoph Fleege, Danny Chan, Frank Zaucke, Andrea Meurer, Marcus Rickert, and et al. 2020. "Adrenoceptor Expression during Intervertebral Disc Degeneration" International Journal of Molecular Sciences 21, no. 6: 2085. https://doi.org/10.3390/ijms21062085
APA StyleKupka, J., Kohler, A., El Bagdadi, K., Bostelmann, R., Brenneis, M., Fleege, C., Chan, D., Zaucke, F., Meurer, A., Rickert, M., & Jenei-Lanzl, Z. (2020). Adrenoceptor Expression during Intervertebral Disc Degeneration. International Journal of Molecular Sciences, 21(6), 2085. https://doi.org/10.3390/ijms21062085