Drug Conjugation Induced Modulation of Structural and Membrane Interaction Features of Cationic Cell-Permeable Peptides

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Peptide Synthesis and Characterization
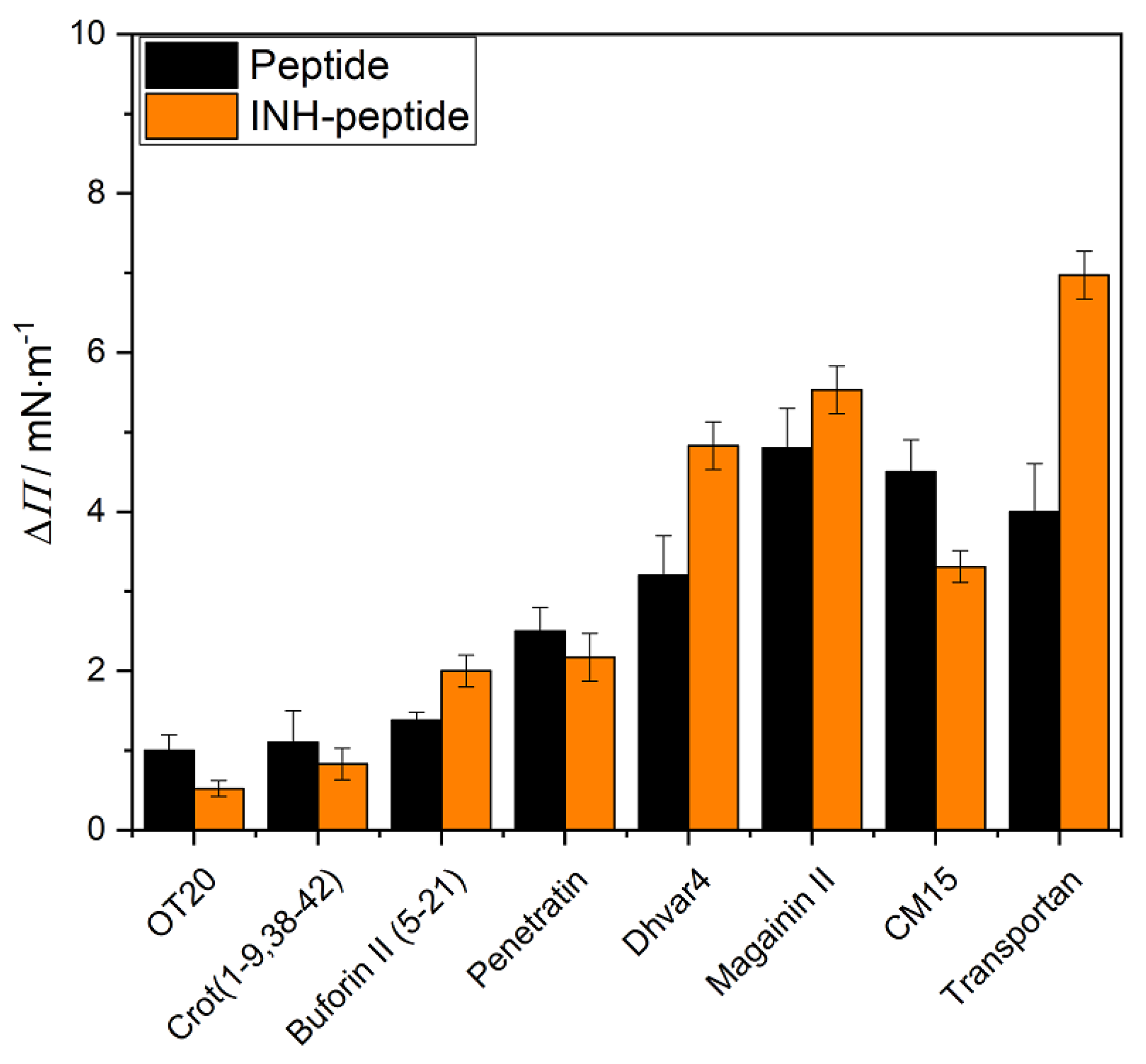
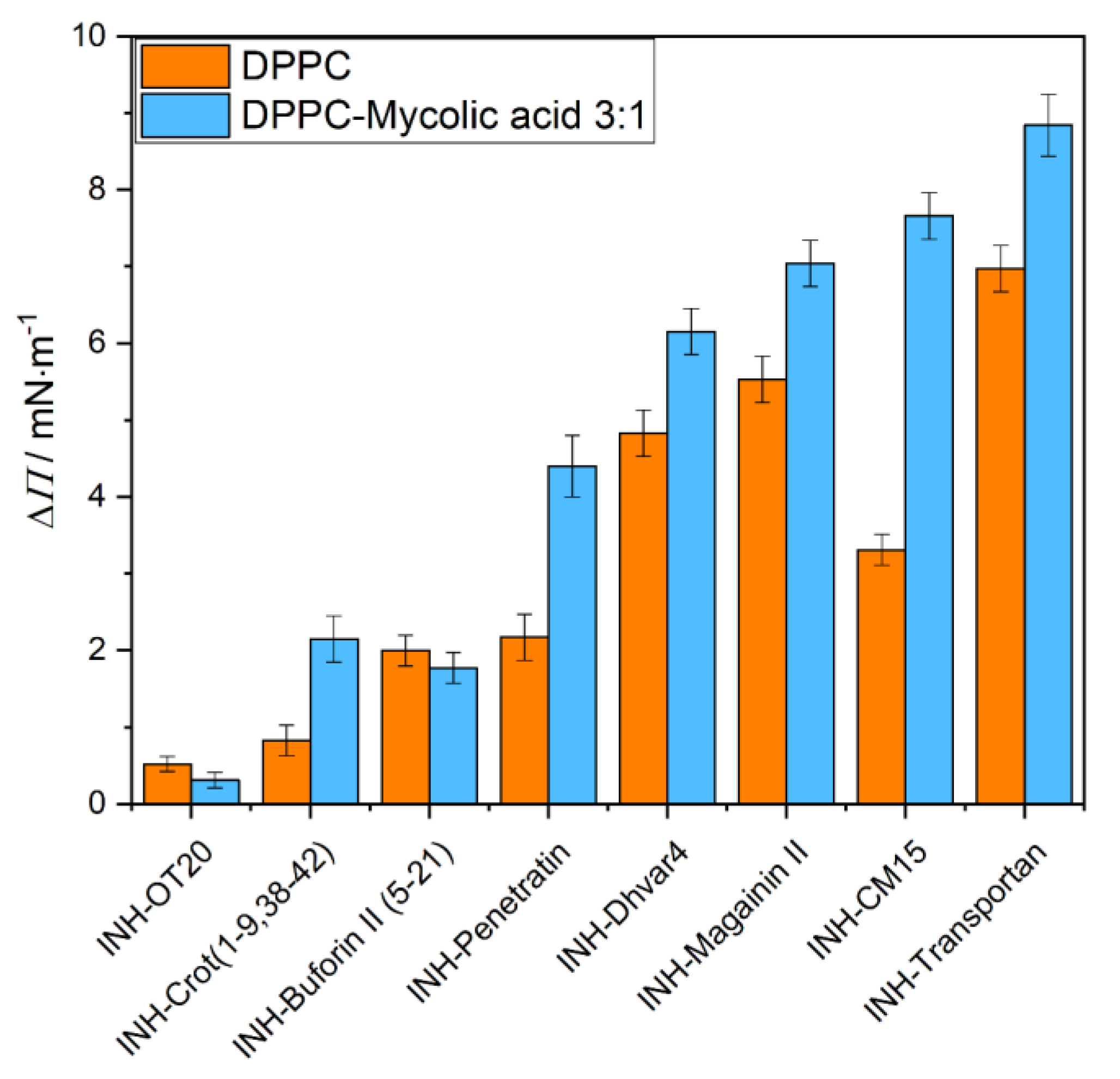
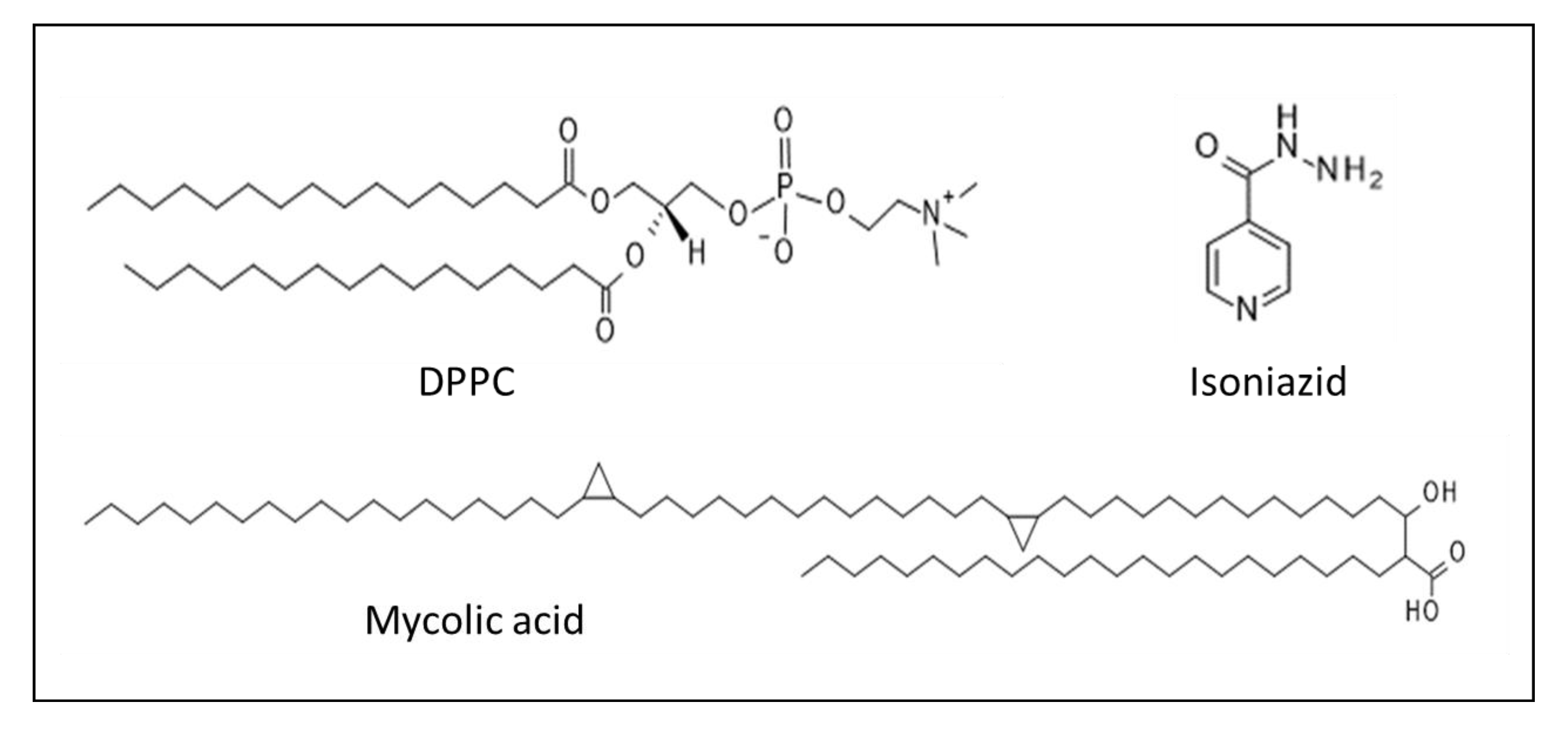
2.2. Interaction of Peptides and INH–Peptide Conjugates with Lipid Monolayers
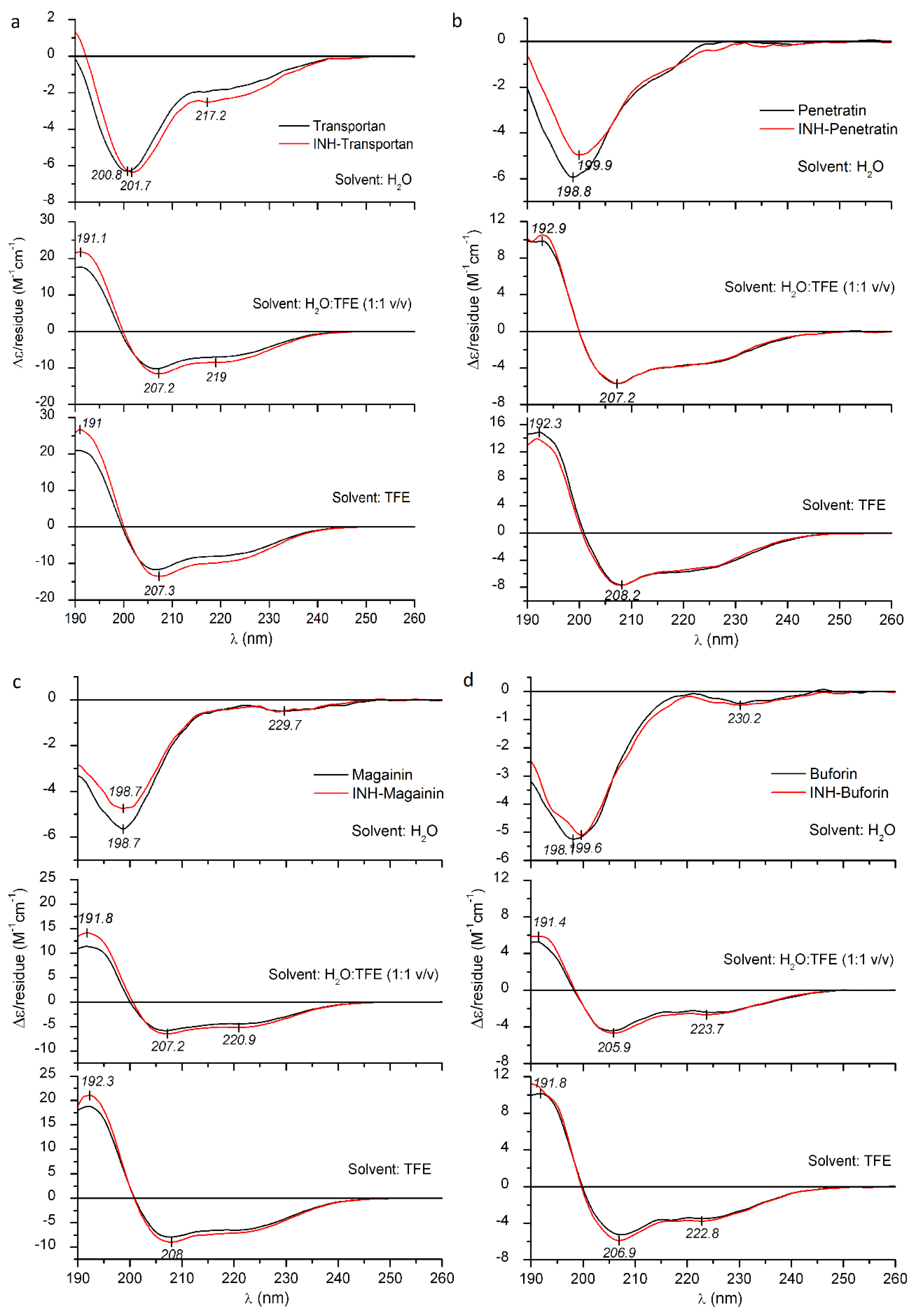
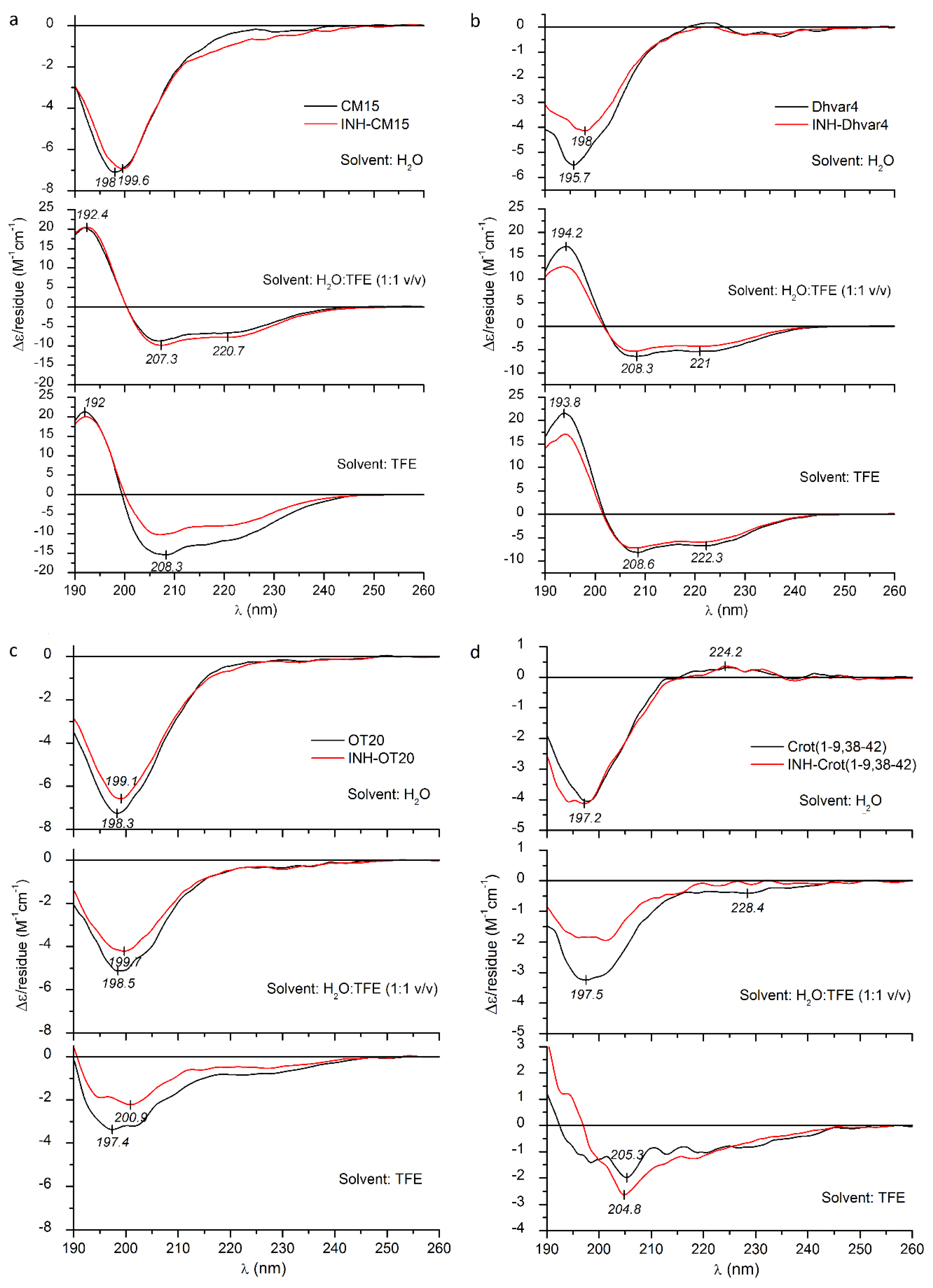
2.3. Circular Dichroism Spectroscopic Evaluation of the Secondary Structure of CPPs/AMPs and Their INH Conjugates
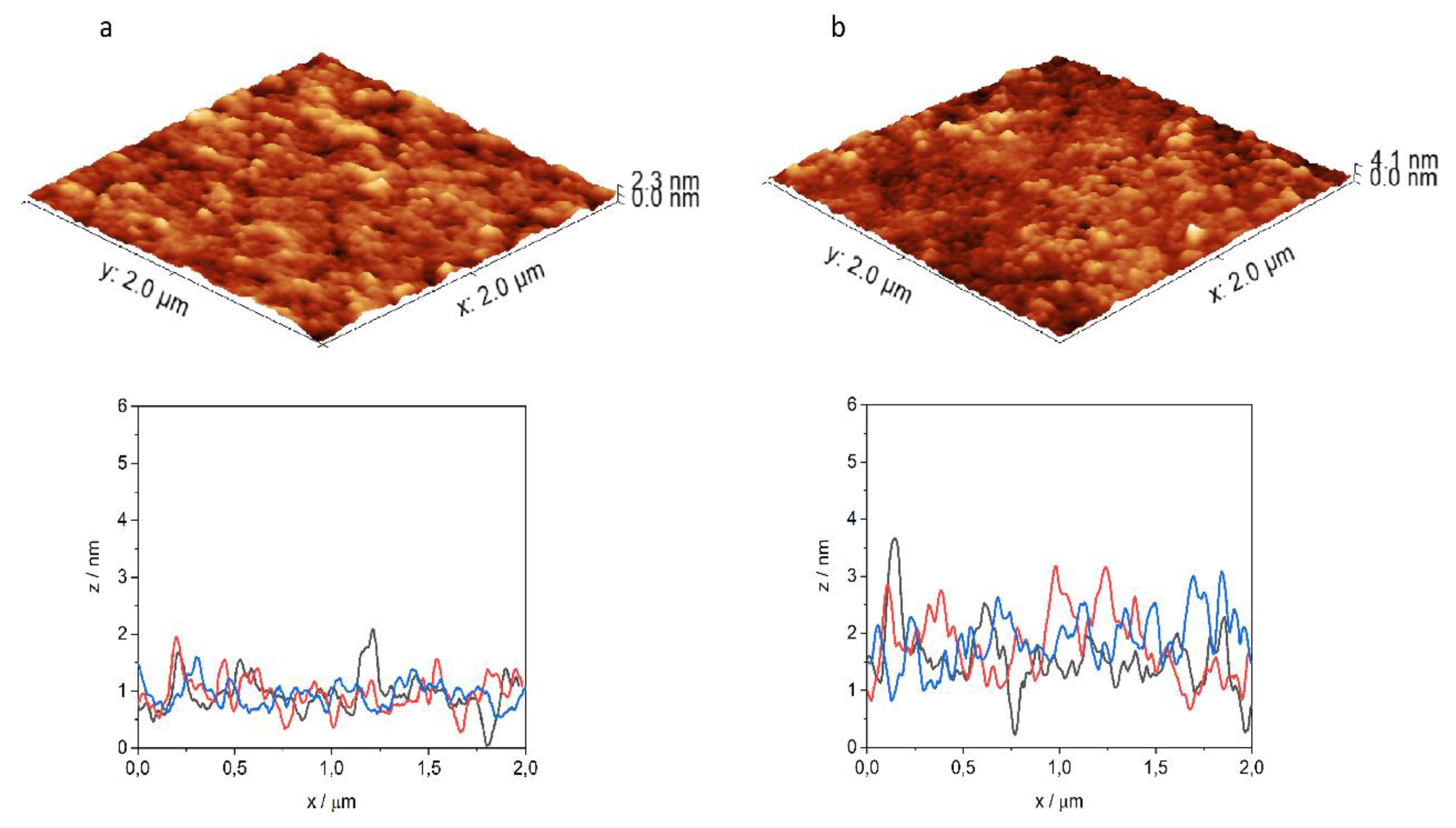
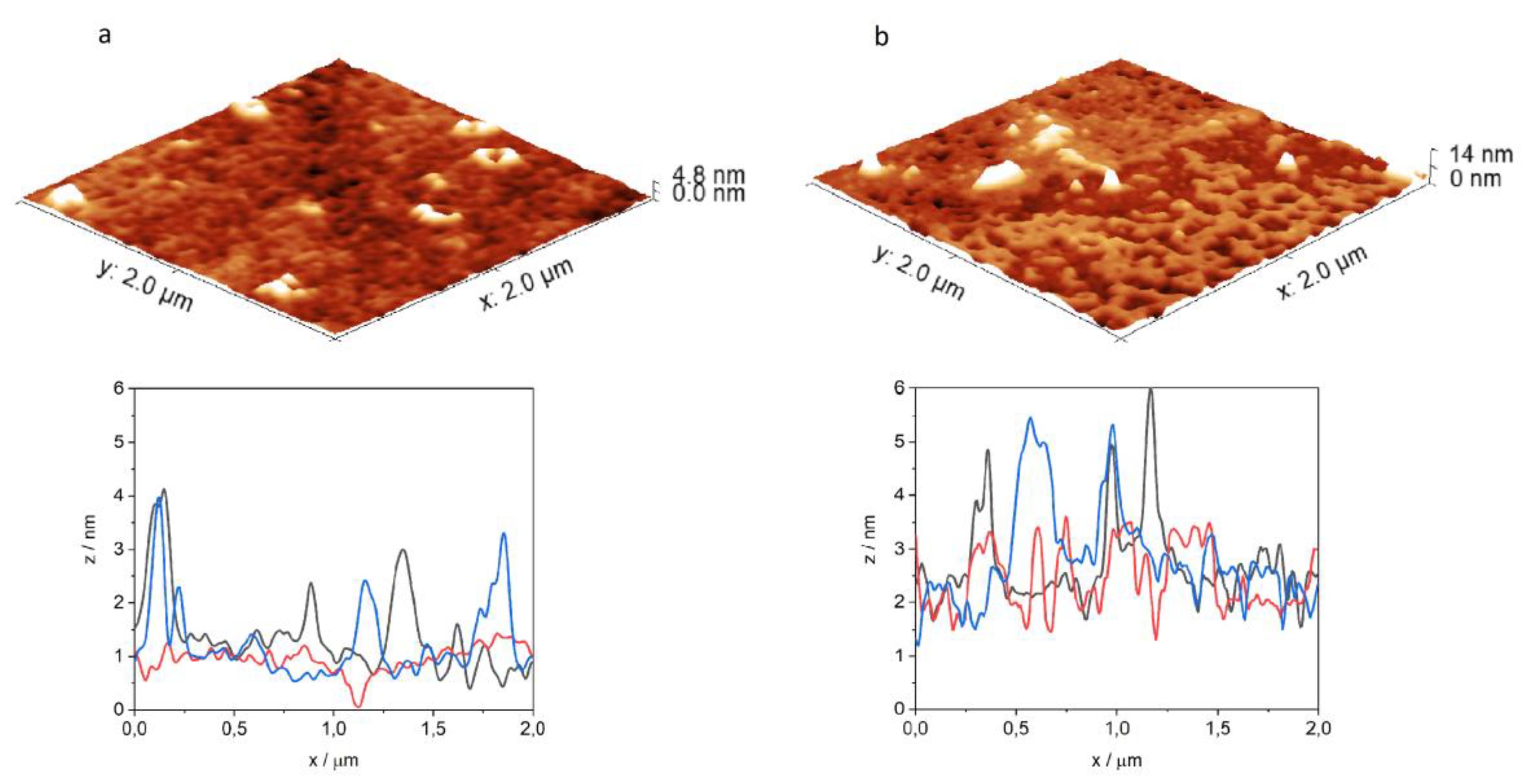
2.4. Atomic Force Microscopy (AFM)
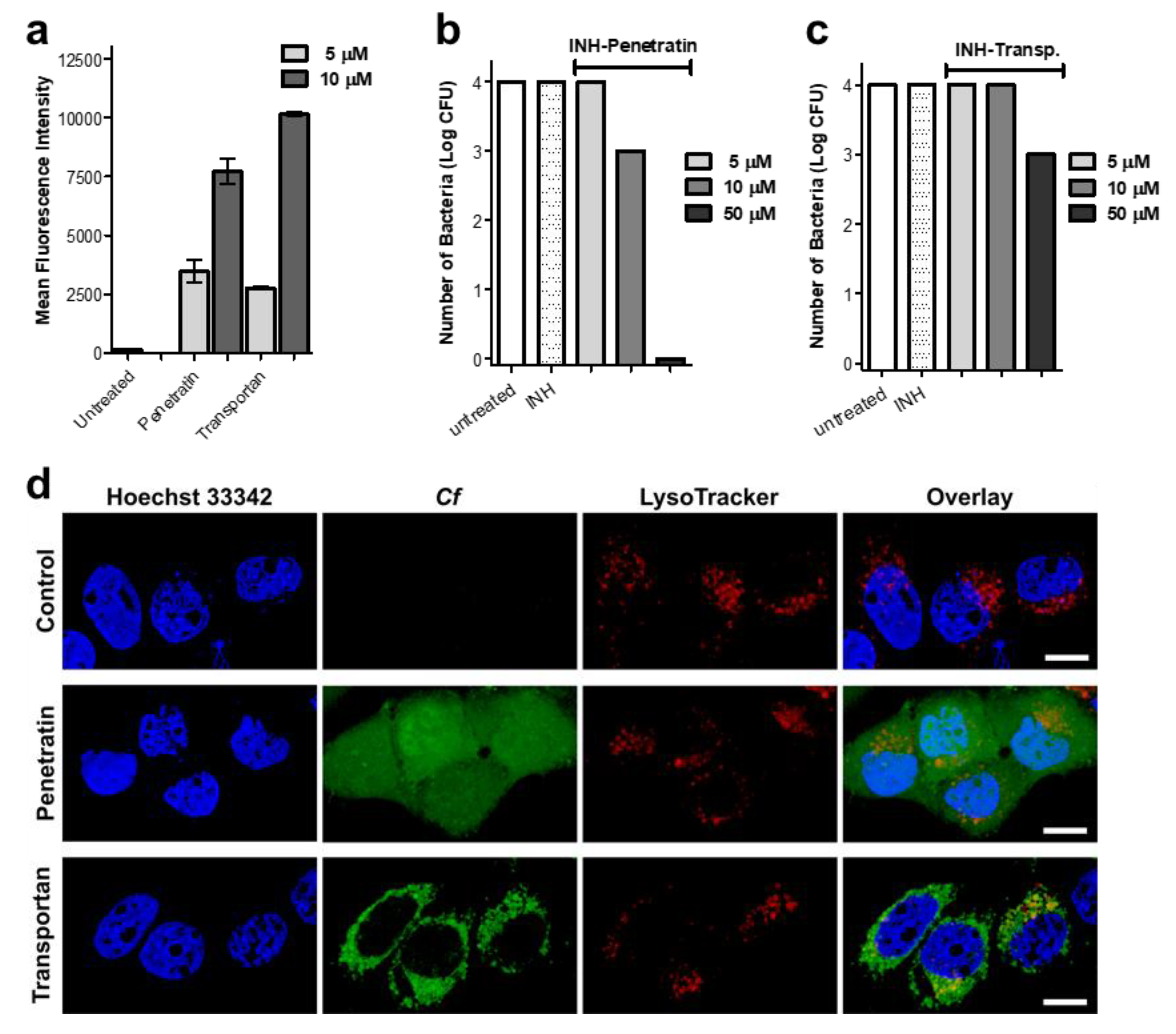
2.5. Cellular Uptake, Localization and Efficacy against Intracellular Mycobacterium Tuberculosis
3. Materials and Methods
3.1. Peptide Synthesis and Characterization
Synthetic Procedures
3.2. Membrane Affinity Measurements
3.3. Circular Dichroism (CD) Spectroscopic Measurements
3.4. Atomic Force Microscopy Measurements
3.5. In Vitro Assays
3.5.1. Cellular Uptake and Localization of the Peptides
3.5.2. Determination of Antibacterial Efficacy against Intracellular Mycobacterium tuberculosis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Han, X.Y.; Seo, Y.H.; Sizer, K.C.; Schoberle, T.; May, G.S.; Spencer, J.S.; Li, W.; Nair, R.G. A new Mycobacterium species causing diffuse lepromatous leprosy. Am. J. Clin. Pathol. 2008, 13, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Global Tuberculosis Report 2018; Report No.; WHO: Geneva, Switzerland, 2018; ISBN 978-92-4-156564-6. [Google Scholar]
- Chiaradia, L.; Lefebvre, C.; Parra, J.; Marcoux, J.; Burlet-Schiltz, O.; Etienne, G.; Tropis, M.; Daffé, M. Dissecting the mycobacterial cell envelope and defining the composition of the native mycomembrane. Nat. Sci. Rep. 2017, 12807, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuber, B.; Chami, M.; Houssin, C.; Dubochet, J.; Griffiths, G.; Daffé, M. Direct visualization of the outer membrane of mycobacteria and corynebacteria in their native state. J. Bacteriol. 2008, 190, 5672–5680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Chevalier, F.; Cascioferro, A.; Majlessi, L.; Herrmann, J.L.; Brosch, R. Mycobacterium tuberculosis evolutionary pathogenesis and its putative impact on drug development. Future Microbiol. 2014, 9, 969–985. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, T.; Amino, S.; Kitamura, S.; Matsumoto, L.; Katada, S.; Nishijo, J. Study of the Molecular Conformation of r- and Keto-Mycolic Acid Monolayers by the Langmuir-Blodgett Technique and Fourier Transform Infrared Reflection-Absorption Spectroscopy. Langmuir 2003, 19, 105–109. [Google Scholar] [CrossRef]
- Chimote, G.; Banerjee, R. Lung surfactant dysfunction in tuberculosis: Effect of mycobacterial tubercular lipids on dipalmitoylphosphatidylcholine surface activity. Colloid. Surface B. 2005, 45, 215–223. [Google Scholar] [CrossRef]
- Groenewald, W.; Baird, M.S.; Verschoor, J.A.; Minnikin, D.E.; Croft, A.K. Differential spontaneous folding of mycolic acids from Mycobacterium tuberculosis. Chem. Phys. Lipids 2014, 180, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, M.; Giner-Casares, J.J.; Lúcio, M.; Caio, J.M.; Moiteiro, C.; Lima, J.L.; Reis, S.; Camacho, L. Interplay of mycolic acids, antimycobacterial compounds and pulmonary surfactant membrane: A biophysical approach to disease. Biochim. Biophys. Acta 2013, 1828, 896–905. [Google Scholar] [CrossRef] [Green Version]
- McNeil, M.; Brennan, P. Structure, function and biogenesis of the cell envelope of mycobacteria in relation to bacterial physiology, pathogenesis and drug resistance; some thoughts and possibilities arising from recent structural information. Res. Microbiol. 1991, 142, 451–463. [Google Scholar] [CrossRef]
- Phan, M.D.; Shin, K.A. Langmuir Monolayer: Ideal Model Membrane to Study Cell. J. Chem. Biol. Interfaces 2014, 2, 1–5. [Google Scholar] [CrossRef]
- Gyulai, G.; Pénzes, C.B.; Mohai, M.; Csempesz, F.; Kiss, É. Influence of surface properties of polymeric drug delivery nanoparticles on their membrane affinity. Eur. Polym. J. 2013, 49, 2495–2503. [Google Scholar] [CrossRef]
- Ábrahám, Á.; Katona, M.; Kasza, G.; Kiss, É. Amphiphilic polymer layer–cell membrane interaction studied by QCM and AFM in model systems. Eur. Polym. J. 2017, 93, 212–221. [Google Scholar] [CrossRef]
- Hasegawa, T.; Nishijo, J.; Watanabe, M. Conformational characterization of r-mycolic acid in a monolayer film by the Langmuir-Blodgett technique and atomic force microscopy. Langmuir 2000, 16, 7325–7330. [Google Scholar] [CrossRef]
- Knobloch, J.; Suhendro, D.K.; Zieleniecki, J.L.; Shapter, J.G.; Köper, I. Membrane–drug interactions studied using model membrane systems. Saudi. J. Biol. Sci. 2015, 22, 714–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimote, G.; Banerjee, R. Effect of antitubercular drugs on dipalmitoylphosphatidylcholine monolayers: Implications for drug loaded surfactants. Respir. Physiol. Neurobiol. 2005, 145, 65–77. [Google Scholar] [CrossRef]
- Pénzes, C.B.; Schnöller, D.; Horváti, K.; Bősze, S.; Mező, G.; Kiss, É. Membrane affinity of antituberculotic drug conjugate using lipid monolayer containing mycolic acid. Colloid. Surface. A. 2012, 413, 142–148. [Google Scholar] [CrossRef]
- Hill, K.; Pénzes, C.B.; Schnöller, D.; Horváti, K.; Bősze, S.; Hudecz, F.; Keszthelyi, T.; Kiss, É. Characterization of the membrane affinity of an isoniazide peptide-conjugate by tensiometry, atomic force microscopy and sum-frequency vibrational spectroscopy, using a phospholipid Langmuir monolayer model. Phys. Chem. Chem. Phys. 2010, 12, 11498–11506. [Google Scholar] [CrossRef] [Green Version]
- Ábrahám, Á.; Baranyai, Z.; Gyulai, G.; Pári, E.; Horváti, K.; Bősze, S.; Kiss, É. Comparative analysis of new antitubercular drug peptide conjugates—Model membrane and in vitro studies. Colloid. Surface B. 2016, 147, 106–115. [Google Scholar] [CrossRef]
- Schnöller, D.; Pénzes, C.B.; Horváti, K.; Bősze, S.; Hudecz, F.; Kiss, É. Membrane affinity of new antitubercular drug candidates using a phospholipid Langmuir monolayer model and LB technique. Prog. Colloid Polym. Sci. 2011, 138, 131–138. [Google Scholar]
- Kiss, É.; Heine, E.T.; Hill, K.; He, Y.-C.; Keusgen, N.; Pénzes, C.B.; Schnöller, D.; Gyulai, G.; Mendrek, A.; Keul, H.; et al. Membrane affinity and antimicrobal properties of polyelectrolytes with different hydrophobicity. Macromol. Biosci. 2012, 12, 1181–1189. [Google Scholar] [CrossRef]
- Nyström, L.; Malmsten, M. Membrane interactions and cell selectivity of amphiphilic anticancer peptides. Curr. Opin. Colloid Interface Sci. 2018, 38, 1–17. [Google Scholar] [CrossRef]
- Derossi, D.; Joliot, A.H.; Chassaing, G.; Prochiantz, A. The third helix of the Antennapedia homeodomain translocates through biological membranes. J. Biol. Chem. 1994, 269, 10444–10450. [Google Scholar]
- Langel, U.; Pooga, M.; Kairane, C.; Zilmer, M.; Bartfai, T. A galanin-mastoparan chimeric peptide activates the Na+,K(+)-ATPase and reverses its inhibition by ouabain. Regul. Pept. 1996, 62, 47–52. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, M.S.; Kim, S.C. A novel antimicrobial peptide from Bufo bufo gargarizans. Biochem. Biophys. Res. Commun. 1996, 218, 408–413. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef] [Green Version]
- Helmerhorst, E.J.; Van’t Hof, W.; Veerman, E.C.; Simoons-Smit, I.; Amerongen, A.V.N. Synthetic histatin analogues with broad-spectrum antimicrobial activity. Biochem. J. 1997, 326, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Andreu, D.; Ubach, J.; Boman, A.; Wahlin, B.; Wade, D.; Merrifield, R.B.; Boman, H.G. Shortened cecropin A-melittin hybrids. Significant size reduction retains potent antibiotic activity. FEBS Lett. 1992, 296, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Radis-Baptista, G.; de la Torre, B.G.; Andreu, D. A novel cell-penetrating peptide sequence derived by structural minimization of a snake toxin exhibits preferential nucleolar localization. J. Med. Chem. 2008, 51, 7041–7044. [Google Scholar] [CrossRef]
- Mezo, G.; Kalászi, A.; Reményi, J.; Majer, Z.; Hilbert, A.; Láng, O.; Köhidai, L.; Barna, K.; Gaál, D.; Hudecz, F. Synthesis, conformation, and immunoreactivity of new carrier molecules based on repeated tuftsin-like sequence. Biopolymers 2004, 73, 645–656. [Google Scholar] [CrossRef]
- Kalafatovic, D.; Giralt, E. Cell-penetrating peptides: Design strategies beyond primary structure and amphipathicity. Molecules 2017, 22, 1929. [Google Scholar] [CrossRef] [Green Version]
- Di Pisa, M.; Chassaing, G.; Swiecicki, J.M. Translocation mechanism(s) of cell-penetrating peptides: Biophysical studies using artificial membrane bilayers. Biochemistry 2015, 54, 194–207. [Google Scholar] [CrossRef]
- Lee, E.Y.; Wong, G.C.L.; Ferguson, A.L. Machine learning-enabled discovery and design of membrane active peptides. Bioorg. Med. Chem. 2018, 26, 2708–2718. [Google Scholar] [CrossRef]
- Horváti, K.; Bacsa, B.; Mlinkó, T.; Szabó, N.; Hudecz, F.; Zsila, F.; Bősze, S. Comparative analysis of internalisation, haemolytic, cytotoxic and antibacterial effect of membrane-active cationic peptides: Aspects of experimental setup. Amino Acids 2017, 49, 1053–1067. [Google Scholar] [CrossRef] [Green Version]
- Horváti, K.; Mező, G.; Szabó, N.; Hudecz, F.; Bősze, S. Peptide conjugates of therapeutically used antitubercular isoniazid-design, synthesis and antimycobacterial effect. J. Pept. Sci. 2009, 15, 385–391. [Google Scholar] [CrossRef]
- Hopp, T.P.; Woods, K.R. Prediction of protein antigenic determinants from amino acid sequences. Proc. Natl. Acad. Sci. USA 1981, 78, 3824–3828. [Google Scholar] [CrossRef] [Green Version]
- Kiss, É.; Gyulai, G.; Pári, E.; Horváti, K.; Bősze, S. Membrane affinity and fluorescent labelling: Comparative study of monolayer interaction, cellular uptake and cytotoxicity profile of carboxyfluorescein-conjugated cationic peptides. Amino Acids 2018, 50, 1557–1571. [Google Scholar] [CrossRef] [Green Version]
- Via, M.A.; Del Pópolo, M.G.; Wilke, N. Negative dipole potentials and carboxylic polar head-groups foster the insertion of cell-penetrating-peptides into lipid monolayers. Langmuir 2018, 34, 3102–3111. [Google Scholar] [CrossRef]
- Toniolo, C.; Formaggio, F.; Woody, R.W. Electronic Circular Dichroism of Peptides. In Comprehensive Chiroptical Spectroscopy: Applications in Stereochemical Analysis of Synthetic Compounds, Natural Products, and Biomolecules; Berova, N., Polavarapu, P.L., Nakanishi, K., Woody, R.W., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 499–544. [Google Scholar]
- Gopal, R.; Park, J.S.; Seo, C.H.; Park, Y. Applications of circular dichroism for structural analysis of gelatin and antimicrobial peptides. Int. J. Mol. Sci. 2012, 13, 3229–3244. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of α-helical antimicrobial peptides. Antimicrob. Agents. Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Ikonomova, S.P.; Karlsson, A.J. Secondary structure of cell-penetrating peptides during interaction with fungal cells. Protein. Sci. 2018, 27, 702–713. [Google Scholar] [CrossRef] [Green Version]
- Dathe, M.; Wieprecht, T. Structural features of helical antimicrobial peptides: Their potential to modulate activity on model membranes and biological cells. Biochim. Biophys. Acta 1999, 1462, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Arouri, A.; Dathe, M.; Blume, A. The helical propensity of KLA amphipathic peptides enhances their binding to gel-state lipid membranes. Biophys. Chem. 2013, 180, 10–21. [Google Scholar] [CrossRef]
- Cherry, M.A.; Higgins, S.K.; Melroy, H.; Lee, H.S.; Pokorny, A. Peptides with the same composition, hydrophobicity, and hydrophobic moment bind to phospholipid bilayers with different affinities. J. Phys. Chem. B. 2014, 118, 12462–12470. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Liang, X.; Liu, C.; Cheng, Y.; Zhou, L.; Wang, K.; Zhao, L. Influence of proline substitution on the bioactivity of mammalian-derived antimicrobial peptide NK-2. Probiotics Antimicrob. Proteins 2018, 10, 118–127. [Google Scholar] [CrossRef]
- Imai, K.; Mitaku, S. Mechanisms of secondary structure breakers in soluble proteins. Biophysics 2005, 1, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Zsila, F.; Bősze, S.; Horváti, K.; Szigyárto, I.C.; Beke-Somfai, T. Drug and dye binding induced folding of the intrinsically disordered antimicrobial peptide CM15. RSC Adv. 2017, 7, 41091–41097. [Google Scholar] [CrossRef] [Green Version]
- Zsila, F.; Juhász, T.; Bősze, S.; Horváti, K.; Beke-Somfai, T. Hemin and bile pigments are the secondary structure regulators of intrinsically disordered antimicrobial peptides. Chirality 2018, 30, 195–205. [Google Scholar] [CrossRef]
- Zsila, F.; Kohut, G.; Beke-Somfai, T. Disorder-to-helix conformational conversion of the human immunomodulatory peptide LL-37 induced by antiinflammatory drugs, food dyes and some metabolites. Int. J. Biol. Macromol. 2019, 129, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Garcin, D.; Marq, J.B.; Iseni, F.; Martin, S.; Kolakofsky, D. A short peptide at the amino terminus of the Sendai virus C protein acts as an independent element that induces STAT1 instability. J. Virol. 2004, 78, 8799–8811. [Google Scholar] [CrossRef] [Green Version]
- Fealey, M.E.; Binder, B.P.; Uversky, V.N.; Hinderliter, A.; Thomas, D.D. Structural impact of phosphorylation and dielectric constant variation on synaptotagmin’s IDR. Biophys. J. 2018, 114, 550–561. [Google Scholar] [CrossRef] [Green Version]
- Anderson, V.L.; Ramlall, T.F.; Rospigliosi, C.C.; Webb, W.W.; Eliezer, D. Identification of a helical intermediate in trifluoroethanol-induced alpha-synuclein aggregation. Proc. Natl. Acad. Sci. USA 2010, 107, 18850–18855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalbandian, A.; Yan, B.S.; Pichugin, A.; Bronson, R.T.; Kramnik, I. Lung carcinogenesis induced by chronic tuberculosis infection: The experimental model and genetic control. Oncogene 2009, 28, 1928–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cukic, V. The Association Between Lung Carcinoma and Tuberculosis. Med. Arch. 2017, 71, 212–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derossi, D.; Chassaing, G.; Prochiantz, A. Trojan peptides: The penetratin system for intracellular delivery. Trends. Cell. Biol. 1998, 8, 84–87. [Google Scholar] [CrossRef]
- LeCher, J.C.; Nowak, S.J.; McMurry, J.L. Breaking in and busting out: Cell-penetrating peptides and the endosomal escape problem. Biomol. Concepts 2017, 8, 131–141. [Google Scholar] [CrossRef]
- Hiraki, S.; Miyai, M.; Seto, T.; Tamura, T.; Watanabe, Y.; Ozawa, S.; Ikeda, H.; Nakata, Y.; Ohnoshi, T.; Kimura, I. Establishment of human continuous cell lines from squamous cell, adeno- and small cell carcinoma of the lung and the results of hetero transplantation. Lung Cancer 1982, 22, 53–58. [Google Scholar]
- Imanishi, K.; Yamaguchi, K.; Suzuki, M.; Honda, S.; Yanaihara, N.; Abe, K. Production of transforming growth factor-alpha in human tumour cell lines. Br. J. Cancer 1989, 59, 761–765. [Google Scholar] [CrossRef] [Green Version]
- Horváti, K.; Bacsa, B.; Szabó, N.; Dávid, S.; Mező, G.; Grolmusz, V.; Vértessy, B.; Hudecz, F.; Bősze, S. Enhanced cellular uptake of a new, in silico identified antitubercular candidate by peptide conjugation. Bioconjug. Chem. 2012, 23, 900–907. [Google Scholar]
- Abes, R.; Arzumanov, A.A.; Moulton, H.M.; Abes, S.; Ivanova, G.D.; Iversen, P.L.; Gait, M.J.; Lebleu, B. Cell-penetrating-peptide-based delivery of oligonucleotides: An overview. Biochem. Soc. Trans. 2007, 35, 775–779. [Google Scholar] [CrossRef] [Green Version]
- Mäe, M.; Langel, U. Cell-penetrating peptides as vectors for peptide, protein and oligonucleotide delivery. Curr. Opin. Pharmacol. 2006, 6, 509–514. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound a | Sequence | Mb | Rtc | Hd | Ze |
|---|---|---|---|---|---|
| OT20 | TKPKGTKPKGTKPKGTKPKG | 2062.3 | 6.7 | 1.1 | 9+ |
| Crot(1–9,38–42) | YKQCHKKGGKKGSG | 1503.8 | 6.8 | 0.8 | 6+ |
| Buforin II (5–21) | RAGLQFPVGRVHRLLRK | 2001.2 | 10.9 | 0.2 | 6+ |
| Penetratin | RQIKIWFQNRRMKWKK | 2244.3 | 10.9 | 0.5 | 8+ |
| Dhvar4 | KRLFKKLLFSLRKY | 1838.2 | 11.8 | 0.3 | 7+ |
| Magainin II | GIGKFLHSAKKFGKAFVGEIMNS | 2464.3 | 12.8 | −0.1 | 4+ |
| CM15 | KWKLFKKIGAVLKVL | 1769.2 | 13.7 | −0.1 | 6+ |
| Transportan | AGYLLGKINLKALAALAKKIL | 2180.4 | 16.8 | −0.3 | 5+ |
| INH–OT20 | INH-TKPKGTKPKGTKPKGTKPKG | 2239.3 | 7.0 | - | 8+ |
| INH–Crot(1–9,38–42) | INH-YKQCHKKGGKKGSG | 1680.9 | 7.3 | - | 5+ |
| INH–Buforin II (5–21) | INH-RAGLQFPVGRVHRLLRK | 2178.3 | 11.0 | - | 5+ |
| INH–Penetratin | INH-RQIKIWFQNRRMKWKK | 2421.4 | 11.5 | - | 7+ |
| INH–Dhvar4 | INH-KRLFKKLLFSLRKY | 2015.2 | 12.8 | - | 6+ |
| INH–Magainin II | INH-GIGKFLHSAKKFGKAFVGEIMNS | 2641.4 | 13.3 | - | 3+ |
| INH–CM15 | INH-KWKLFKKIGAVLKVL | 1946.2 | 14.3 | - | 5+ |
| INH–Transportan | INH-AGYLLGKINLKALAALAKKIL | 2357.5 | 16.9 | - | 4+ |
| Peptide | Lipid System | Ra/nm | Rq/nm | Rz/nm |
|---|---|---|---|---|
| DPPC | 0.10 | 0.13 | 0.36 | |
| DPPC+mycolic acid | 0.25 | 0.33 | 1.08 | |
| INH–Penetratin | DPPC | 0.21 | 0.27 | 0.92 |
| DPPC+mycolic acid | 0.35 | 0.45 | 1.33 | |
| INH–Transportan | DPPC | 0.23 | 0.37 | 0.90 |
| DPPC+mycolic acid | 0.59 | 0.94 | 2.40 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pári, E.; Horváti, K.; Bősze, S.; Biri-Kovács, B.; Szeder, B.; Zsila, F.; Kiss, É. Drug Conjugation Induced Modulation of Structural and Membrane Interaction Features of Cationic Cell-Permeable Peptides. Int. J. Mol. Sci. 2020, 21, 2197. https://doi.org/10.3390/ijms21062197
Pári E, Horváti K, Bősze S, Biri-Kovács B, Szeder B, Zsila F, Kiss É. Drug Conjugation Induced Modulation of Structural and Membrane Interaction Features of Cationic Cell-Permeable Peptides. International Journal of Molecular Sciences. 2020; 21(6):2197. https://doi.org/10.3390/ijms21062197
Chicago/Turabian StylePári, Edit, Kata Horváti, Szilvia Bősze, Beáta Biri-Kovács, Bálint Szeder, Ferenc Zsila, and Éva Kiss. 2020. "Drug Conjugation Induced Modulation of Structural and Membrane Interaction Features of Cationic Cell-Permeable Peptides" International Journal of Molecular Sciences 21, no. 6: 2197. https://doi.org/10.3390/ijms21062197
APA StylePári, E., Horváti, K., Bősze, S., Biri-Kovács, B., Szeder, B., Zsila, F., & Kiss, É. (2020). Drug Conjugation Induced Modulation of Structural and Membrane Interaction Features of Cationic Cell-Permeable Peptides. International Journal of Molecular Sciences, 21(6), 2197. https://doi.org/10.3390/ijms21062197