Arbuscular Mycorrhizal Symbiosis Mitigates Iron (Fe)-Deficiency Retardation in Alfalfa (Medicago sativa L.) Through the Enhancement of Fe Accumulation and Sulfur-Assisted Antioxidant Defense

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Impact of AMF on Alfalfa Morpho-Physiological Indices
2.2. Impact of AMF on the Mineral Content
2.3. Role of AMF Overcoming Oxidative Stress in Alfalfa
2.4. Effect of AMF on the Expression of Fe and S Transporter Genes
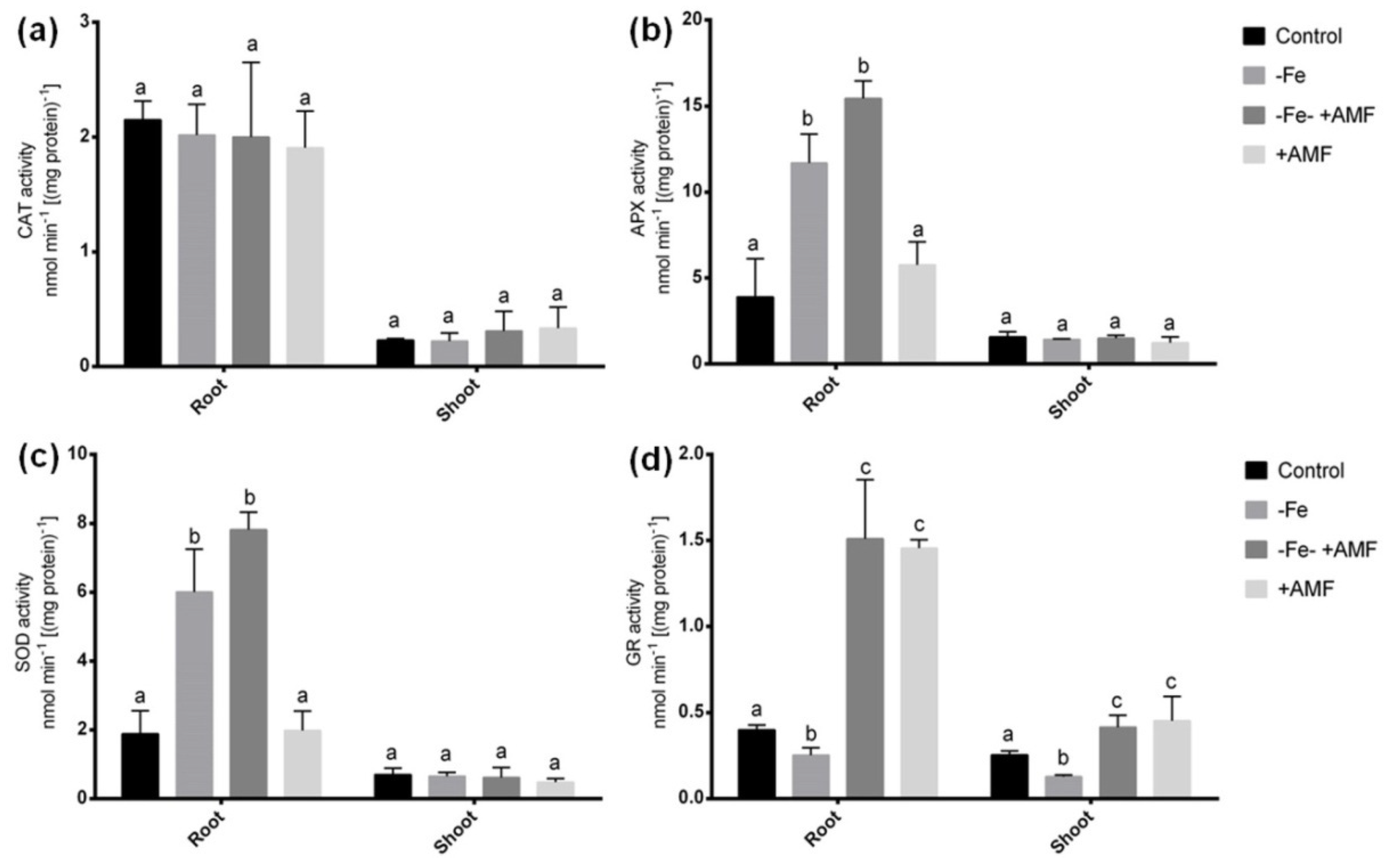
2.5. Role of AMF on Antioxidant Enzymes in Alfalfa
2.6. Effect of AMF on S-Metabolites
2.7. Sulfur Deprivation Effect Under Fe Deficiency
3. Discussion
3.1. Improvement of Alfalfa Plant Biomass and Physiological Parameters
3.2. Enhancement of Mineral Content in Alfalfa
3.3. Antioxidant Responses and Alleviation of Oxidative Stress Injury
4. Materials and Methods
4.1. Alfalfa Cultivation and AMF Supplementation
4.2. Determination of Mycorrhizal Colonization
4.3. Analysis of Morpho-Physiological Parameters
4.4. ICP-MS Analysis
4.5. Determination of Fe Chelate Reductase Activity
4.6. Estimation of Rhizosphere Acidification
4.7. Determinatin of Soluble Protein
4.8. Estimation of Electrolyte Leakage
4.9. Determination of Hydrogen Peroxide and Cell Death
4.10. RNA Extraction, cDNA Synthesis, and qRT-PCR Analysis
4.11. Analysis of S-Metabolites
4.12. Determination of Antioxidant Enzymes
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walker, E.L.; Connolly, E.L. Time to pump iron: iron-deficiency-signaling mechanisms of higher plants. Curr. Opin. Plant Biol. 2008, 11, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, E.; Romera, F.J.; Cañete, M.; de la Guardia, M.D. Effects of bicarbonate and iron supply on Fe(III) reducing capacity of roots and leaf chlorosis of the susceptible peach rootstock “Nemaguard”. J. Plant Nurt. 2000, 23, 1607–1617. [Google Scholar] [CrossRef]
- Kabir, A.H.; Paltridge, N.G.; Roessner, U.; Stangoulis, J.C.R. Mechanisms associated with Fe-deficiency tolerance and signaling in shoots of Pisum sativum. Physiol. Plant 2013, 147, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-Y.; Xia, R.-X.; Hu, L.-M.; Dong, T.; Wu, Q.-S. Arbuscular mycorrhizal fungi alleviate iron deficient chlorosis in Poncirus trifoliata L. Raf under calcium bicarbonate stress. J. Hortic. Sci. Biotech. 2015, 82, 776–780. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Sun, W.; Wang, T. The adaptive mechanism of plants to iron deficiency via iron uptake, transport, and homeostasis. Int. J. Mol. Sci. 2019, 20, 2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- M’sehli, W.; Jellali, N.; Dell’Orto, M.; Abdelly, C.; Zocchi, G.; Gharsalli, M. Responses of two lines of Medicago ciliaris to Fe deficiency under saline conditions. Plant Growth Regul. 2011, 64, 221–230. [Google Scholar] [CrossRef]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.-F.; Curie, C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [Green Version]
- Kabir, A.H.; Rahman, M.M.; Haider, S.A.; Paul, N.K. Mechanisms associated with differential tolerance to Fe deficiency in okra (Abelmoschus esculentus Moench). Environmen. Exp. Bot. 2015, 112, 16–26. [Google Scholar] [CrossRef]
- Młodzińska, E. Alteration of plasma membrane H+-ATPase in cucumber roots under different iron nutrition. Acta Physiol. Plant. 2012, 34, 2125–2133. [Google Scholar] [CrossRef] [Green Version]
- Kabir, A.H.; Paltridge, N.G.; Able, A.J.; Paull, J.G.; Stangoulis, J.C.R. Natural variation for Fe-efficiency is associated with upregulation of Strategy I mechanisms and enhanced citrate and ethylene synthesis in Pisum sativum L. Planta 2012, 235, 1409–1419. [Google Scholar] [CrossRef]
- Kabir, A.H.; Paltridge, N.G.; Stangoulis, J. Role of sulphur conferring differential tolerance to iron deficiency in Pisum sativum. Biologia 2015, 70, 922–928. [Google Scholar] [CrossRef]
- Sieh, D.; Watanabe, M.; Devers, E.A.; Brueckner, F.; Hoefgen, R.; Krajinski, F. The arbuscular mycorrhizal symbiosis influences sulfur starvation responses of Medicago truncatula. New Phytol. 2013, 197, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.; Hodge, A.; Fitter, A.H. Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material. New Phytol. 2009, 181, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.B.; Zeto, S.K. Mineral acquisition by arbuscular mycorrhizal plants. J. Plant Nurt. 2000, 23, 867–902. [Google Scholar] [CrossRef]
- Harrison, M.J.; Dewbre, G.R.; Liu, J. A phosphate transporter from Medicago truncatula involved in the acquisition of phosphate released by arbuscular mycorrhizal fungi. Plant cell 2002, 14, 2413–2429. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, U.; Schmelzer, E.; Bothe, H. Expression of nitrate transporter genes in tomato colonized by an arbuscular mycorrhizal fungus. Physiol. Plant 2002, 115, 125–136. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Javot, H.; Pumplin, N.; Harrison, M.J. Phosphate in the arbuscular mycorrhizal symbiosis: transport properties and regulatory roles. Plant Cell Environ. 2007, 30, 310–322. [Google Scholar] [CrossRef]
- Nagy, R.; Drissner, D.; Amrhein, N.; Jakobsen, I.; Bucher, M. Mycorrhizal phosphate uptake pathway in tomato is phosphorus-repressible and transcriptionally regulated. New Phytol. 2009, 181, 950–959. [Google Scholar] [CrossRef]
- Li, J.-F.; He, X.-H.; Li, H.; Zheng, W.-J.; Liu, J.-F.; Wang, M.-Y. Arbuscular mycorrhizal fungi increase growth and phenolics synthesis in Poncirus trifoliata under iron deficiency. Sci. Hortic. 2015, 183, 87–92. [Google Scholar] [CrossRef]
- Allen, J.W.; Shachar-Hill, Y. Sulfur transfer through an arbuscular mycorrhiza. Plant Physiol. 2009, 149, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Casieri, L.; Gallardo, K.; Wipf, D. Transcriptional response of Medicago truncatula sulphate transporters to arbuscular mycorrhizal symbiosis with and without sulphur stress. Planta 2012, 235, 1431–1447. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.T.; Fu, J.J.; Sun, Y.F.; Xu, Y.M.; Miao, Y.J.; Xu, Y.F.; Hu, T.M. Effect of arbuscular mycorrhizal fungi inoculation on cold stress-induced oxidative damage in leaves of Elymus nutans Griseb. S. Afr. J. Bot. 2016, 104, 21–29. [Google Scholar] [CrossRef]
- Yang, J.; Teng, Y.; Wang, J.; Li, J. Vanadium uptake by alfalfa grown in V–Cd-contaminated soil by pot experiment. Biol. Trace Elem. Res. 2011, 142, 787–795. [Google Scholar] [CrossRef]
- Carrasco-Gil, S.; Estebaranz-Yubero, M.; Medel-Cuesta, D.; Millán, R.; Hernández, L.E. Influence of nitrate fertilization on Hg uptake and oxidative stress parameters in alfalfa plants cultivated in a Hg-polluted soil. Environ. Exp. Bot. 2012, 75, 16–24. [Google Scholar] [CrossRef]
- Rodríguez-Celma, J.; Lattanzio, G.; Grusak, M.A.; Abadía, A.; Abadía, J.; López-Millán, A.-F. Root responses of Medicago truncatula plants grown in two different iron deficiency conditions: changes in root protein profile and riboflavin biosynthesis. J. Proteome Res. 2011, 10, 2590–2601. [Google Scholar] [CrossRef]
- Allen, A.E.; Laroche, J.; Maheswari, U.; Lommer, M.; Schauer, N.; Lopez, P.J.; Finazzi, G.; Fernie, A.R.; Bowler, C. Whole-cell response of the pennate diatom Phaeodactylum tricornutum to iron starvation. Proc. Natl. Acad. Sci. USA 2008, 105, 10438–10443. [Google Scholar] [CrossRef] [Green Version]
- Behrenfeld, M.J.; Milligan, A.J. Photophysiological expressions of iron stress in phytoplankton. Ann. Rev. Mar. Sci. 2013, 5, 217–246. [Google Scholar] [CrossRef]
- Petrou, K.; Trimborn, S.; Rost, B.; Ralph, P.J.; Hassler, C.S. The impact of iron limitation on the physiology of the antarctic diatom Chaetoceros simplex. Mar. Biol. 2014, 161, 925–937. [Google Scholar] [CrossRef] [Green Version]
- Roncel, M.; González-Rodríguez, A.A.; Naranjo, B.; Bernal-Bayard, P.; Lindahl, A.M.; Hervás, M.; Navarro, J.A.; Ortega, J.M. Iron deficiency induces a partial inhibition of the photosynthetic electron transport and a high sensitivity to light in the diatom Phaeodactylum tricornutum. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwe, M.A.; Stefels, J. Photosynthetic responses in Phaeocystis antarctica towards varying light and iron conditions. Biogeochemistry 2007, 83, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.M.; Guerinot, M.L. Facing the challenges of Cu, Fe and Zn homeostasis in plants. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caris, C.; Hördt, W.; Hawkins, H.-J.; Römheld, V.; George, E. Studies of iron transport by arbuscular mycorrhizal hyphae from soil to peanut and sorghum plants. Mycorrhiza 1998, 8, 35–39. [Google Scholar] [CrossRef]
- Huang, Y.-M.; Wu, Q.-S. Arbuscular mycorrhizal fungi and tolerance of Fe stress in plants. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Wu, Q.-S., Ed.; Springer: Singapore, 2017; pp. 131–145. [Google Scholar]
- Rillig, M.C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Can. J. Soil Sci. 2004, 84, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Curie, C.; Alonso, J.M.; Jean, M.L.; Ecker, J.R.; Briat, J.-F. Involvement of NRAMP1 from Arabidopsis thaliana in iron transport. Biochem. J. 2000, 347, 749–755. [Google Scholar] [CrossRef]
- Zuchi, S.; Cesco, S.; Varanini, Z.; Pinton, R.; Astolfi, S. Sulphur deprivation limits Fe-deficiency responses in tomato plants. Planta 2009, 230, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.; Yu, N.; Bano, S.A.; Liu, C.; Miller, A.J.; Cousins, D.; Zhang, X.; Ratet, P.; Tadege, M.; Mysore, K.S.; et al. A H+-ATPase that energizes nutrient uptake during mycorrhizal symbioses in rice and Medicago truncatula. Plant Cell 2014, 26, 1818–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts-Williams, S.J.; Cavagnaro, T.R. Arbuscular mycorrhizal fungi increase grain zinc concentration and modify the expression of root ZIP transporter genes in a modern barley (Hordeum vulgare) cultivar. Plant Sci. 2018, 274, 163–170. [Google Scholar] [CrossRef]
- Burleigh, S.H.; Kristensen, B.K.; Bechmann, I.E. A plasma membrane zinc transporter from Medicago truncatula is up-regulated in roots by Zn fertilization, yet down-regulated by arbuscular mycorrhizal colonization. Plant Mol. Biol. 2003, 52, 1077–1088. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Cavagnaro, T.R.; Watts-Williams, S.J. The effects of soil phosphorus and zinc availability on plant responses to mycorrhizal fungi: a physiological and molecular assessment. Sci. Rep. 2019, 9, 14880. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive species and antioxidants. Redox biology Is a fundamental theme of aerobic Life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noctor, G.; Foyer, C.H. Simultaneous measurement of foliar glutathione, γ-glutamylcysteine, and amino acids by high-performance liquid chromatography: comparison with two other assay methods for glutathione. Anal. Biochem. 1998, 264, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Kabir, A.H.; Begum, M.C.; Haque, A.; Amin, R.; Swaraz, A.M.; Haider, S.A.; Paul, N.K.; Hossain, M.M. Genetic variation in Fe toxicity tolerance is associated with the regulation of translocation and chelation of iron along with antioxidant defence in shoots of rice. Funct. Plant Biol. 2016, 43, 1070–1081. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Zhang, T.; Sun, Y.; Song, Y.; Tian, C.; Feng, G. On-site growth response of a desert ephemeral plant, Plantago minuta, to indigenous arbuscular mycorrhizal fungi in a central Asia desert. Symbiosis 2011, 55, 77–84. [Google Scholar] [CrossRef]
- Rahman, M.A.; Alam, I.; Kim, Y.-G.; Ahn, N.-Y.; Heo, S.-H.; Lee, D.-G.; Liu, G.; Lee, B.-H. Screening for salt-responsive proteins in two contrasting alfalfa cultivars using a comparative proteome approach. Plant Physiol. Biochem. 2015, 89, 112–122. [Google Scholar] [CrossRef]
- Albano, J.P.; Miller, W.B. Iron deficiency stress influences physiology of iron acquisition in marigold (Tagetes erecta L.). J. Am. Soc. Hortic. Sci. 1996, 121, 438. [Google Scholar] [CrossRef] [Green Version]
- Guy, C.; Haskell, D.; Neven, L.; Klein, P.; Smelser, C. Hydration-state-responsive proteins link cold and drought stress in spinach. Planta 1992, 188, 265–270. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Annal. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Rahman, M.A.; Yong-Goo, K.; Iftekhar, A.; Liu, G.-s.; Hyoshin, L.; Joo, L.J.; Byung-Hyun, L. Proteome analysis of alfalfa roots in response to water deficit stress. J. Integr. Agric. 2016, 15, 1275–1285. [Google Scholar] [CrossRef]
- Zhao, J.; Fujita, K.; Sakai, K. Oxidative stress in plant cell culture: a role in production of β-thujaplicin by Cupresssus lusitanica suspension culture. Biotechnol. Bioeng. 2005, 90, 621–631. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, S.; Landberg, T.; Greger, M. Cadmium uptake and interaction with phytochelatins in wheat protoplasts. Plant Physiol. Biochem. 2007, 45, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Goud, P.B.; Kachole, M.S. Antioxidant enzyme changes in neem, pigeonpea and mulberry leaves in two stages of maturity. Plant Signal Behav. 2012, 7, 1258–1262. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Zigman, S. An improved spectrophotometric assay for superoxide dismutase based on epinephrine autoxidation. Anal. Biochem. 1978, 90, 81–89. [Google Scholar] [CrossRef]
- Halliwell, B.; Foyer, C. Properties and physiological function of a glutathione reductase purified from spinach leaves by affinity chromatography. Planta 1978, 139, 9–17. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root | ||||
|---|---|---|---|---|
| Control | -Fe | -Fe +AMF | +AMF | |
| Fe | 149 ± 19.2 b | 88 ± 17.6 a | 131 ± 19.4 b | 177 ± 48.6 b |
| Zn | 147 ± 50.3 b | 52 ± 15.8 a | 123 ± 23.4 b | 112 ± 4.5 b |
| S | 14623 ± 323.6 b | 11107 ± 1662.5 a | 16177 ± 2571.4 b | 16856 ± 1637.3 b |
| Ca | 738 ± 220.9 b | 447 ± 88.8 a | 1282 ± 174.8 b | 1195 ± 60.0 b |
| P | 953 ± 145.1 c | 296 ± 55.1 a | 551 ± 77.6 b | 824 ± 125.5 c |
| K | 126 ± 64.0 a | 137 ± 24.9 a | 104 ± 21.6 a | 155 ± 55.8 a |
| Mg | 196 ± 21.2 a | 245 ± 56.6 a | 476 ± 69.2 b | 455 ± 11.2 b |
| Shoot | ||||
| Fe | 62 ± 10.6 b | 34 ± 6.5 a | 55 ± 13.2 b | 58 ± 7.7 b |
| Zn | 51 ± 7.6 b | 24 ± 1.4 a | 51 ± 9.6 b | 44 ± 10.8 b |
| S | 3538 ± 262.9 b | 2340 ± 66.4 a | 4479 ± 589.8 b | 4031 ± 1006.5 b |
| Ca | 460 ± 62.3 a | 428 ± 45.9 a | 751 ± 18.0 b | 658 ± 88.2 b |
| P | 260 ± 44.1c | 78 ± 7.0 a | 176 ± 42.5 b | 292 ± 60.2 c |
| K | 47 ± 10.8 a | 47 ± 20.1 a | 53 ± 13.4 a | 68 ± 49.8 a |
| Mg | 39 ± 11.8 a | 35 ± 4.7 a | 40 ± 2.6 a | 32 ± 3.6 a |
| S-metabolites | Control | −Fe | −Fe+AMF | +AMF |
|---|---|---|---|---|
| Glutathione | 5.5 ± 1.09 a | 6.8 ± 0.57 a | 15.0 ± 1.76 b | 6.2 ± 0.59 a |
| Cysteine | 2.4 ± 0.84 a | 2.0 ± 0.96 a | 6.5 ± 1.43 b | 2.7 ± 0.19 a |
| Methionine | 4.3 ± 0.81 a | 4.9 ± 1.03 a | 4.7 ± 0.85 a | 4.4 ± 0.75 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.A.; Parvin, M.; Das, U.; Ela, E.J.; Lee, S.-H.; Lee, K.-W.; Kabir, A.H. Arbuscular Mycorrhizal Symbiosis Mitigates Iron (Fe)-Deficiency Retardation in Alfalfa (Medicago sativa L.) Through the Enhancement of Fe Accumulation and Sulfur-Assisted Antioxidant Defense. Int. J. Mol. Sci. 2020, 21, 2219. https://doi.org/10.3390/ijms21062219
Rahman MA, Parvin M, Das U, Ela EJ, Lee S-H, Lee K-W, Kabir AH. Arbuscular Mycorrhizal Symbiosis Mitigates Iron (Fe)-Deficiency Retardation in Alfalfa (Medicago sativa L.) Through the Enhancement of Fe Accumulation and Sulfur-Assisted Antioxidant Defense. International Journal of Molecular Sciences. 2020; 21(6):2219. https://doi.org/10.3390/ijms21062219
Chicago/Turabian StyleRahman, Md. Atikur, Monika Parvin, Urmi Das, Esrat Jahan Ela, Sang-Hoon Lee, Ki-Won Lee, and Ahmad Humayan Kabir. 2020. "Arbuscular Mycorrhizal Symbiosis Mitigates Iron (Fe)-Deficiency Retardation in Alfalfa (Medicago sativa L.) Through the Enhancement of Fe Accumulation and Sulfur-Assisted Antioxidant Defense" International Journal of Molecular Sciences 21, no. 6: 2219. https://doi.org/10.3390/ijms21062219
APA StyleRahman, M. A., Parvin, M., Das, U., Ela, E. J., Lee, S.-H., Lee, K.-W., & Kabir, A. H. (2020). Arbuscular Mycorrhizal Symbiosis Mitigates Iron (Fe)-Deficiency Retardation in Alfalfa (Medicago sativa L.) Through the Enhancement of Fe Accumulation and Sulfur-Assisted Antioxidant Defense. International Journal of Molecular Sciences, 21(6), 2219. https://doi.org/10.3390/ijms21062219