Asynchronous Embryo Transfer Followed by Comparative Transcriptomic Analysis of Conceptus Membranes and Endometrium Identifies Processes Important to the Establishment of Equine Pregnancy


Abstract
:1. Introduction
2. Results
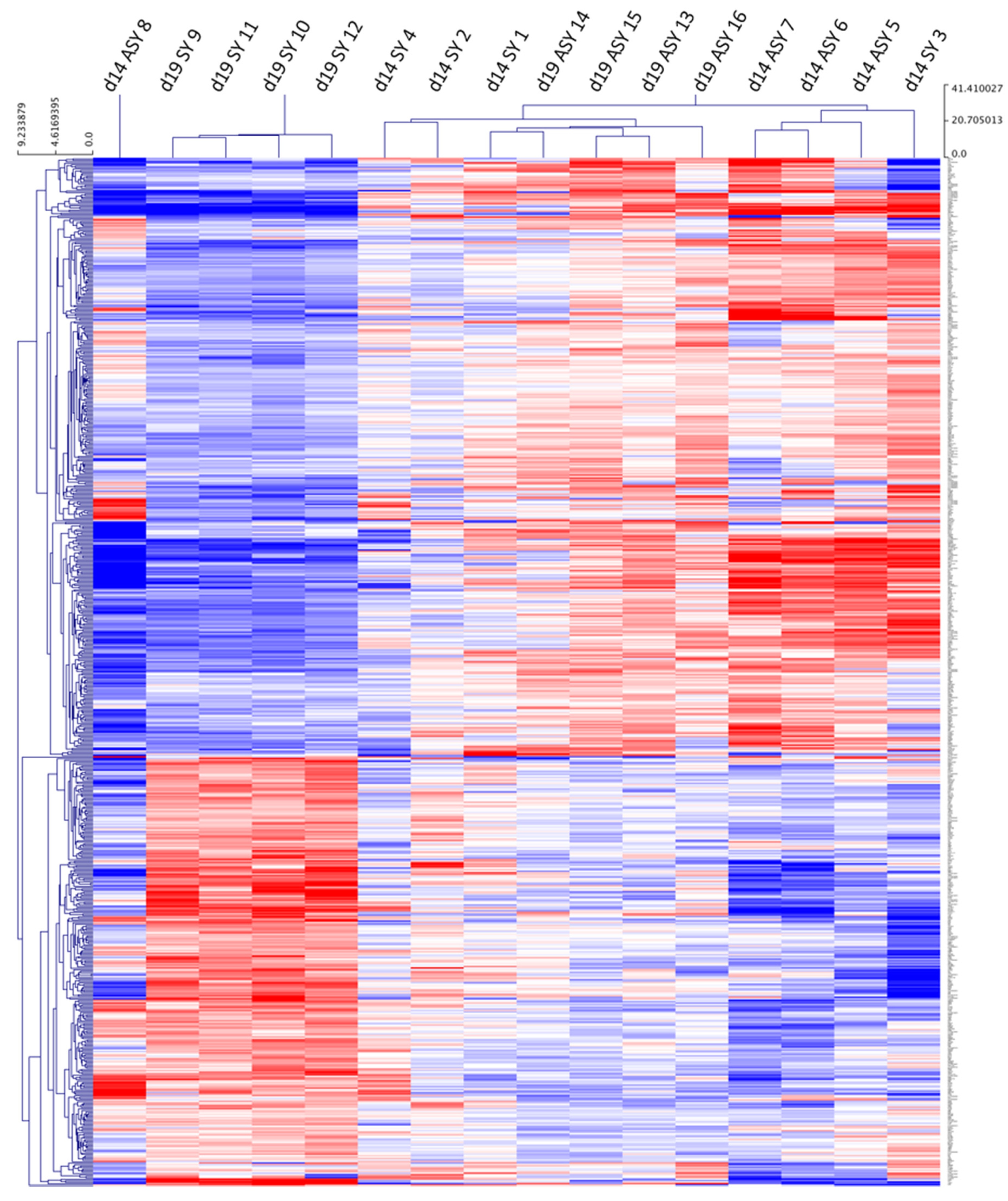
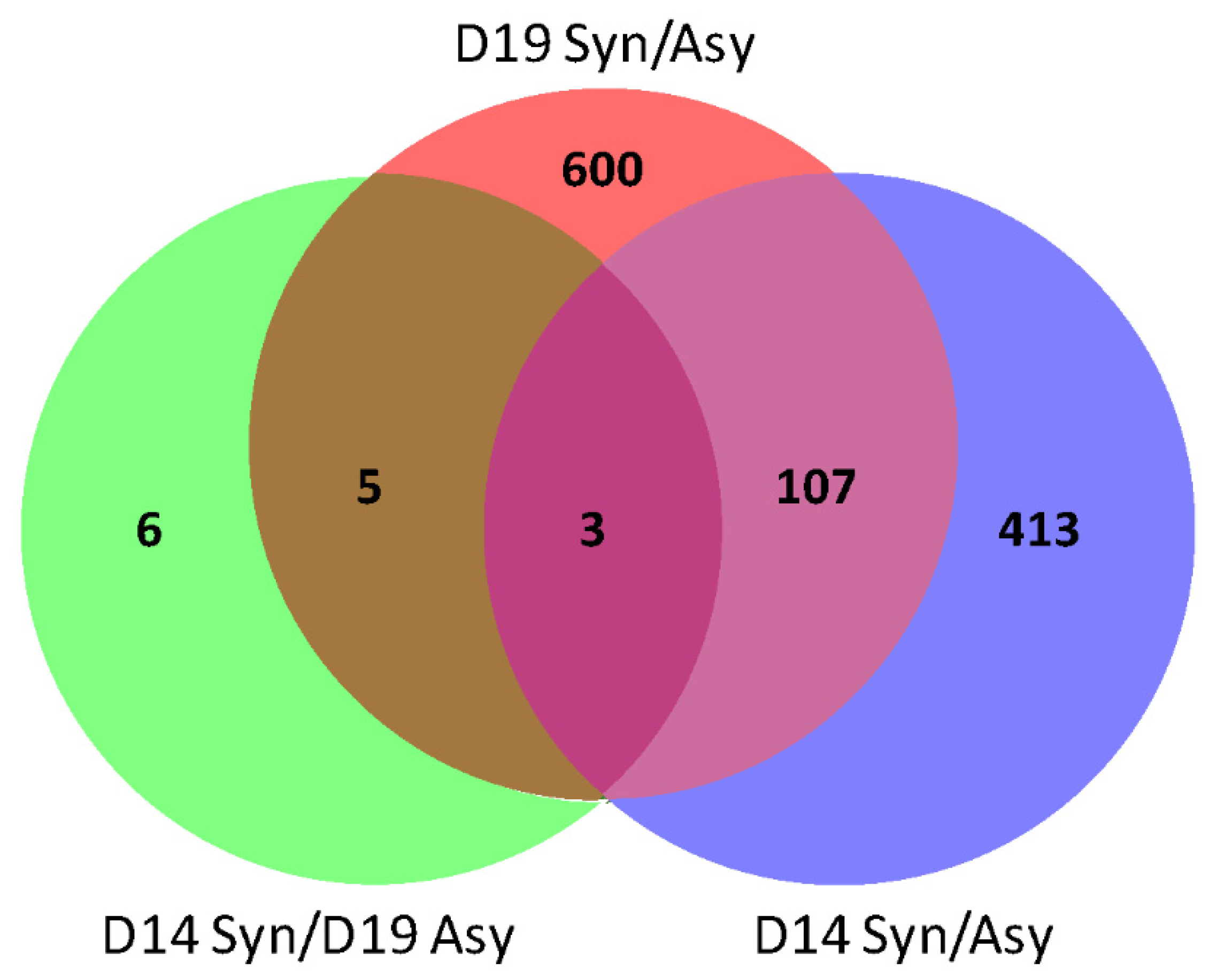
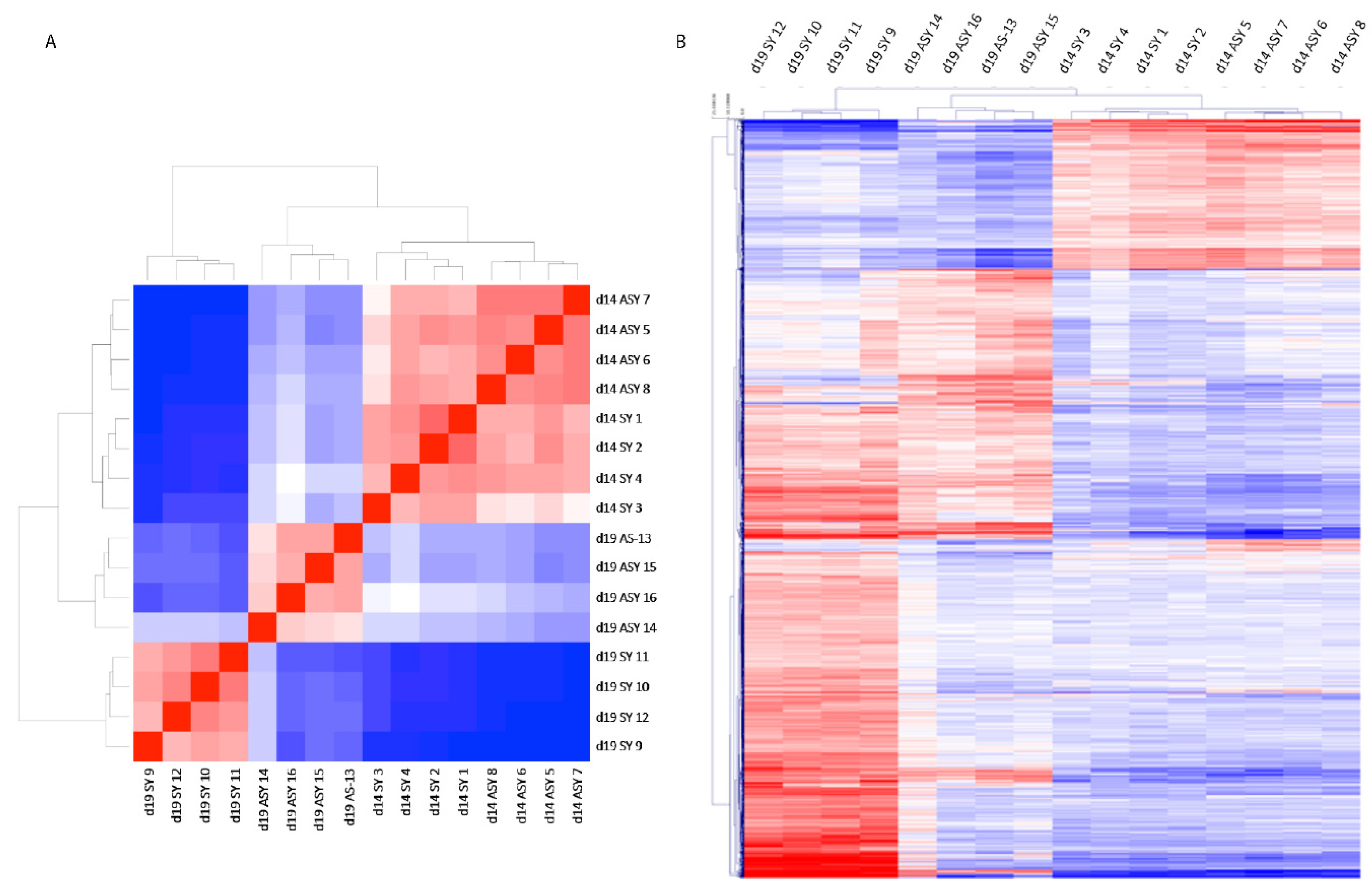
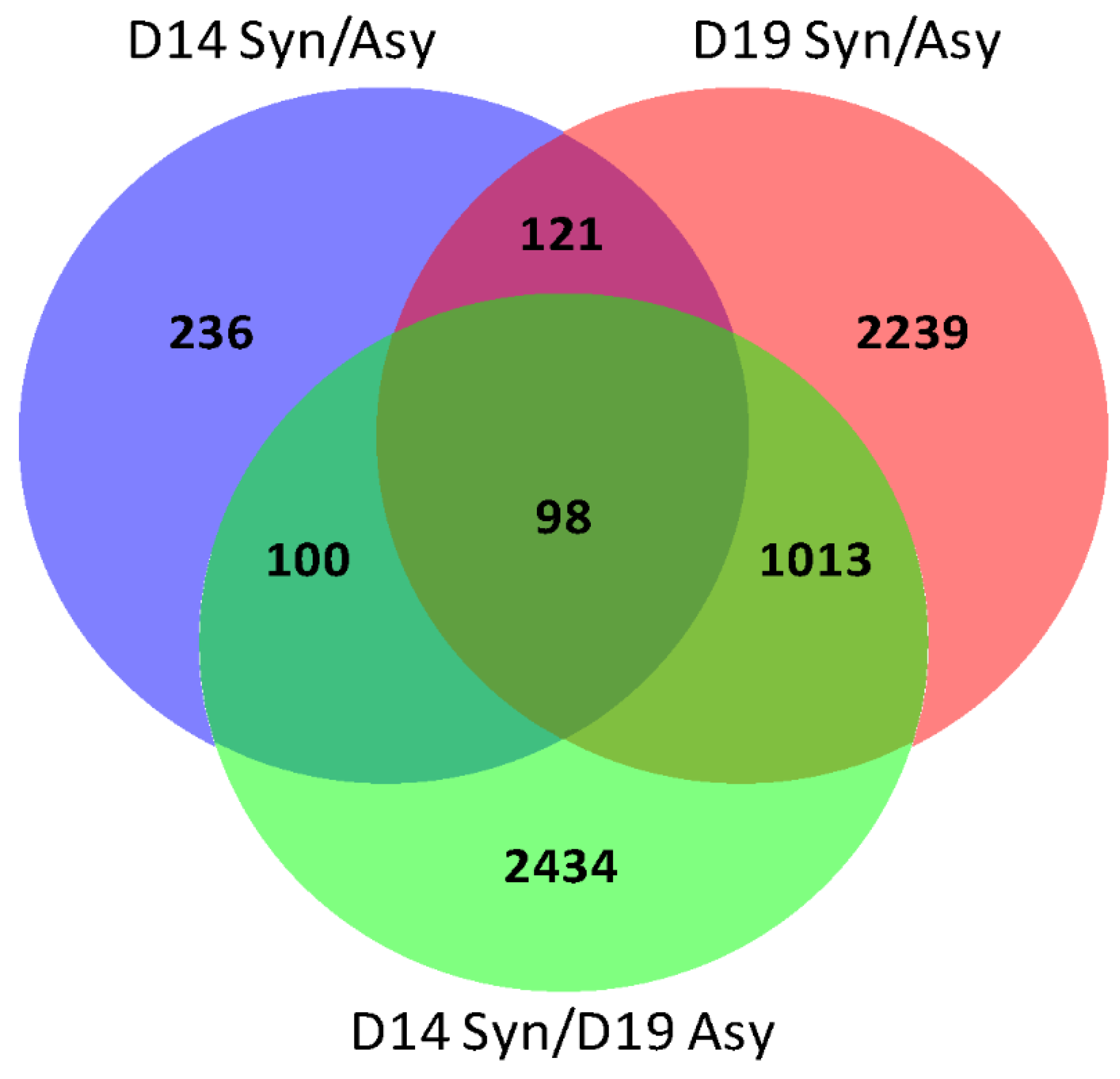
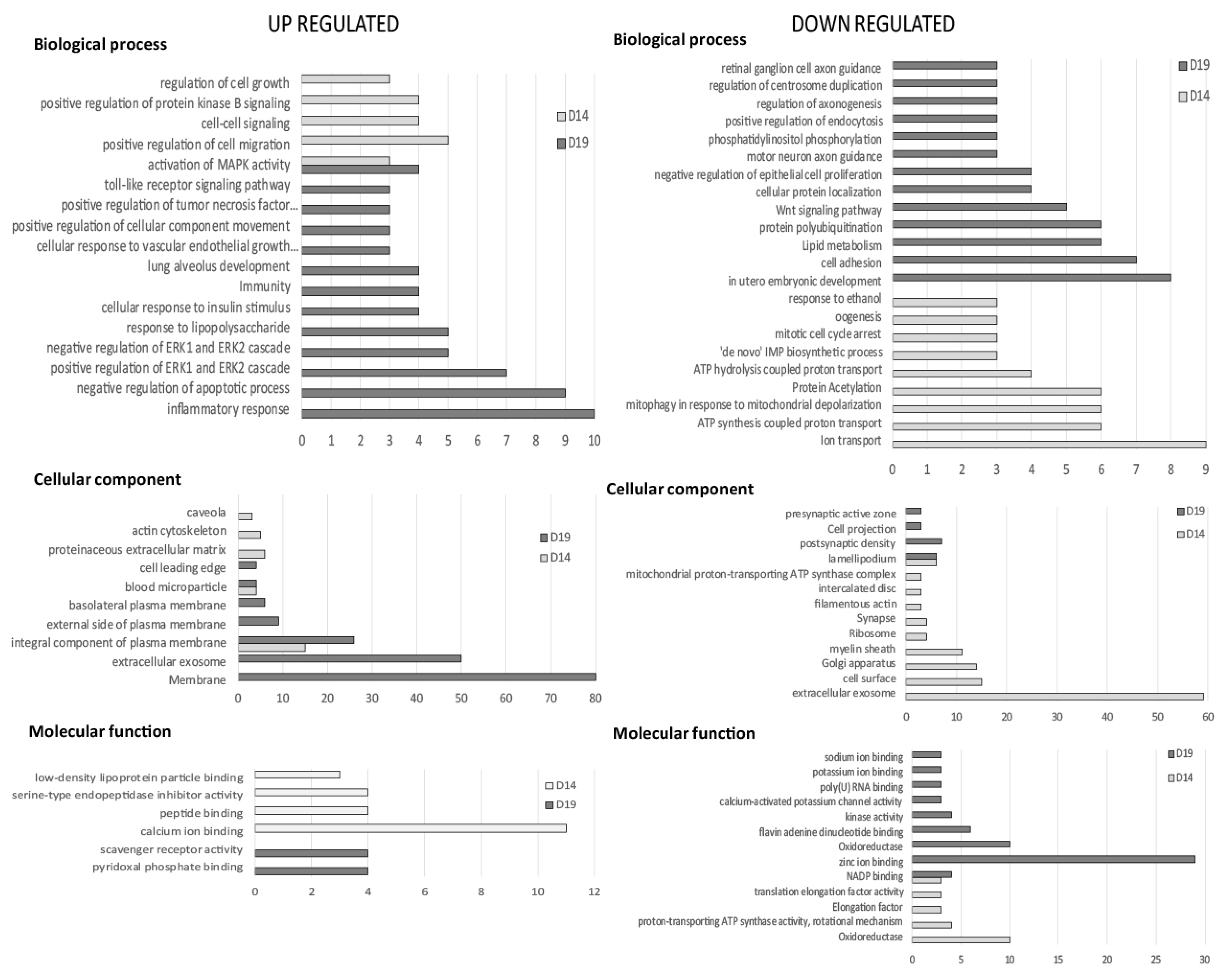
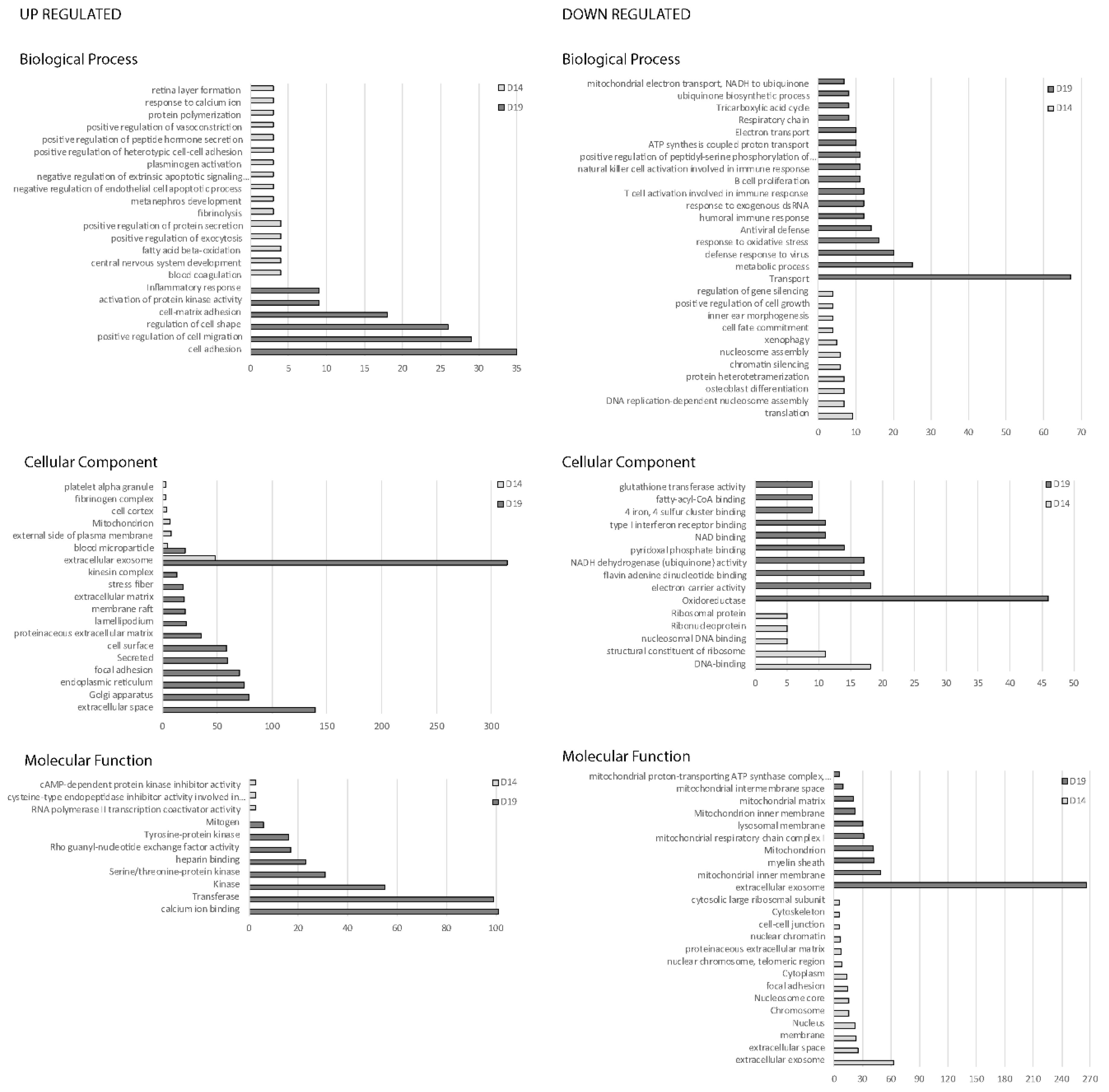
2.1. RNA-seq Analysis and Gene Ontology
2.2. Gene Ontology Analysis
2.3. RNA-seq Validation by qRT-PCR
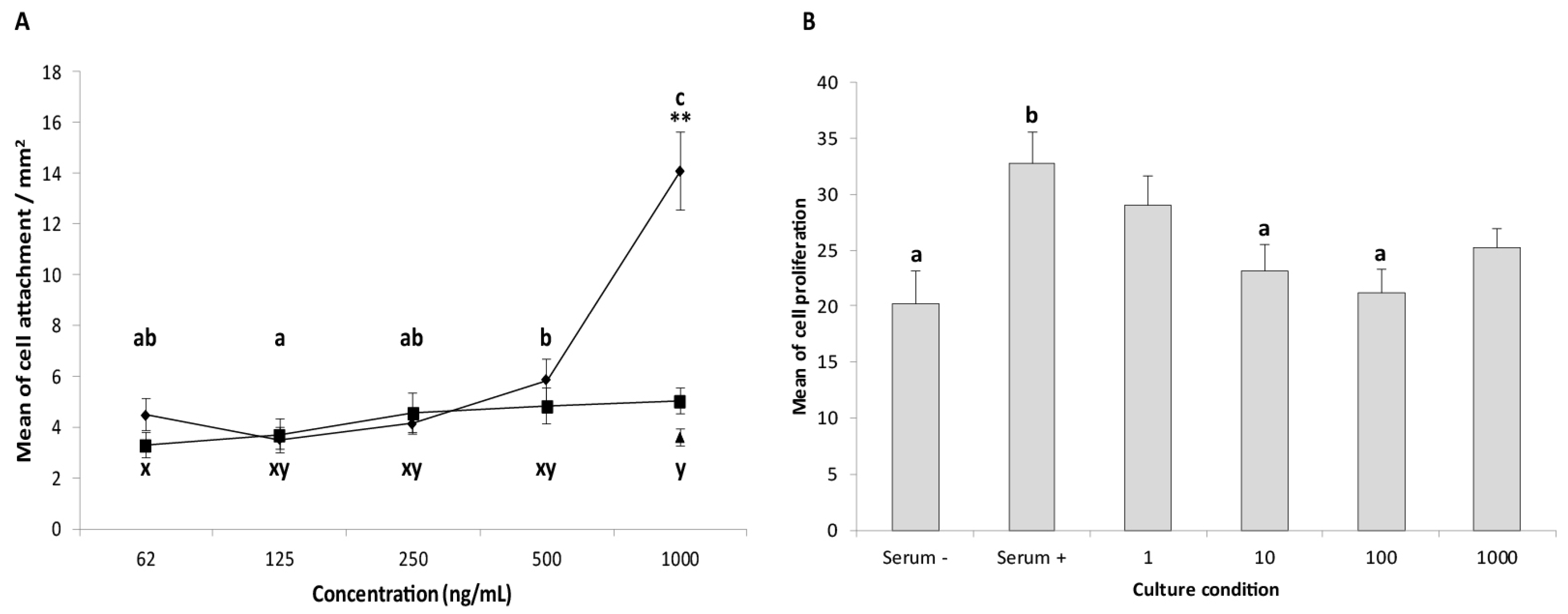
2.4. Cell Proliferation and Cell Attachment in Response to IGFBP3 Stimulation
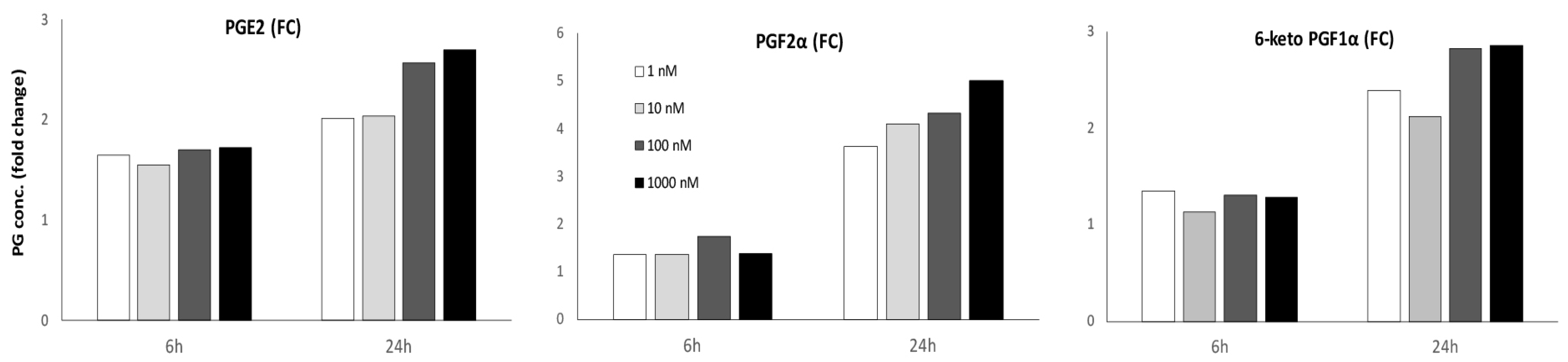
2.5. Prostaglandin Release after Bradykinin Stimulation
3. Discussion
4. Material and Methods
4.1. Animals and Tissue Collection
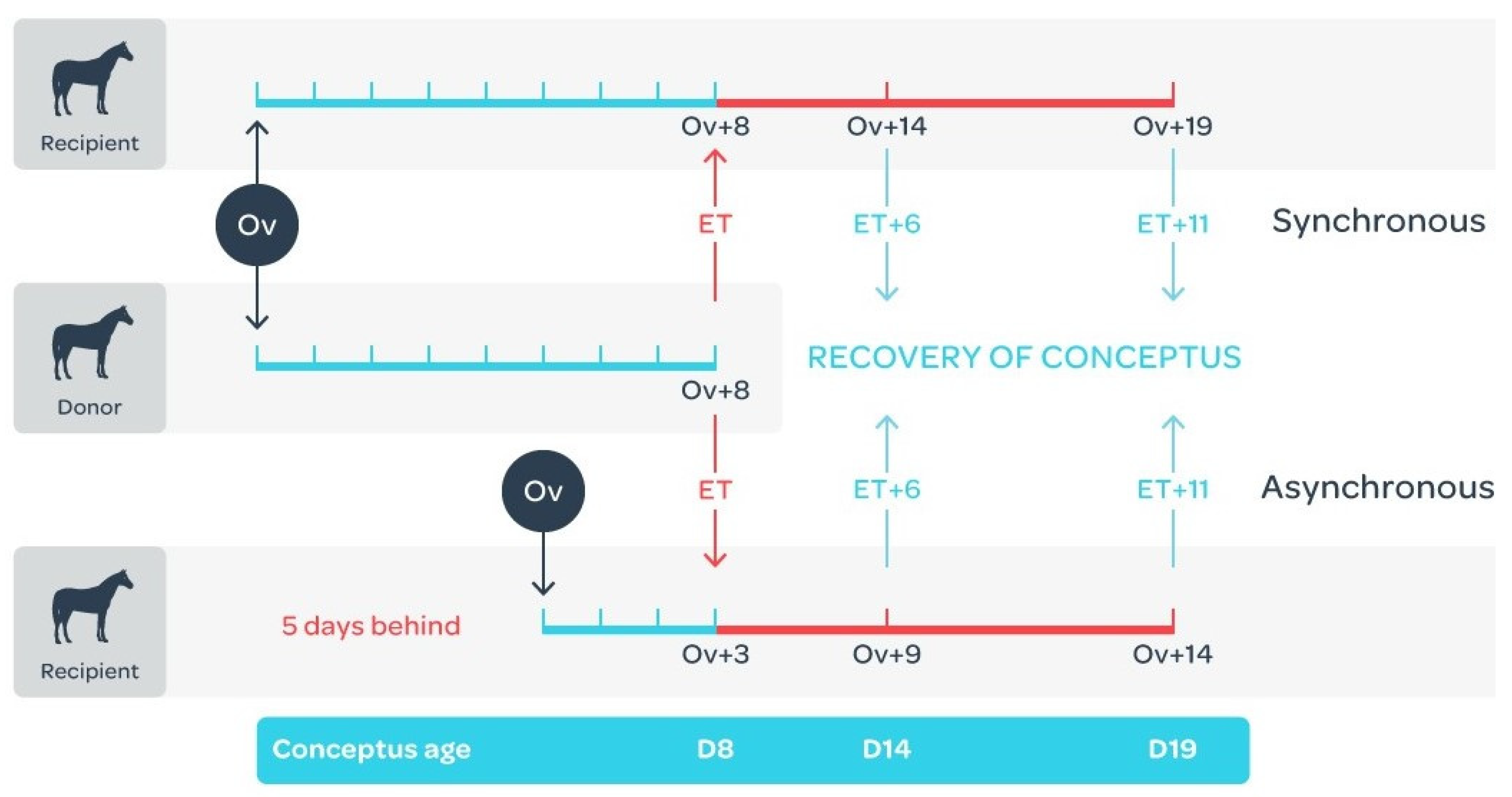
4.2. Embryo Collection and Transfer
4.3. RNA Extraction, DNA Library Preparation and Sequencing
4.4. RNA-seq Analysis and Bioinformatics
4.5. Quantitative RT-PCR
4.6. Culture of Conceptus Membrane Cells
4.7. Cell Attachment Assay
4.8. Cell Proliferation Assay
4.9. Bradykinin Stimulation
4.10. Prostaglandin Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Stout, T.A.E. Embryo-maternal communication during the first 4 weeks of equine pregnancy. Theriogenology 2016, 86, 349–354. [Google Scholar] [CrossRef]
- Freeman, D.A.; Weber, J.A.; Geary, R.T.; Woods, G.L. Time of embryo transport through the mare oviduct. Theriogenology 1991, 36, 823–830. [Google Scholar] [CrossRef]
- Allen, W.R.; Stewart, F. Equine placentation. Reprod. Fertil. Dev. 2001, 13, 623–634. [Google Scholar] [CrossRef]
- Oriol, J.G.; Sharom, F.J.; Betteridge, K.J. Developmentally regulated changes in the glycoproteins of the equine embryonic capsule. J. Reprod. Fertil. 1993, 99, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrler, A.; Stewart, F.; Crossett, B.; Pell, J.M.; Ellis, P.D.; Beier, H.M.; Allen, W.R. Identification of proteins in the equine embryonic capsule. J. Reprod. Fertil. 2000, Suppl 56, 601–606. [Google Scholar]
- Leith, G.S.; Ginther, O.J. Characterization of intrauterine mobility of the early equine conceptus. Theriogenology 1984, 21, 633–644. [Google Scholar] [CrossRef]
- McDowell, K.J.; Sharp, D.C.; Grubaugh, W.; Thatcher, W.W.; Wilcox, C.J. Restricted conceptus mobility results in failure of pregnancy maintenance in mares. Biol. Reprod. 1988, 39, 340–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starbuck, G.R.; Stout, T.A.E.; Lamming, G.E.; Allen, W.R.; Flint, A.P.F. Endometrial oxytocin receptor and uterine prostaglandin secretion in mares during the oestrous cycle and early pregnancy. Reproduction 1998, 113, 173–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stout, T.A.E.; Allen, W.R. Prostaglandin E(2) and F(2 alpha) production by equine conceptuses and concentrations in conceptus fluids and uterine flushings recovered from early pregnant and dioestrous mares. Reproduction 2002, 123, 261–268. [Google Scholar] [CrossRef]
- Goff, A.K.; Pontbriand, D.; Sirois, J. Oxytocin stimulation of plasma 15-keto-13,14-dihydro prostaglandin F-2 alpha during the oestrous cycle and early pregnancy in the mare. J. Reprod. Fertil. 1987, 35, 253–260. [Google Scholar]
- Stout, T.A.; Lamming, G.E.; Allen, W.R. Oxytocin administration prolongs luteal function in cyclic mares. J. Reprod. Fertil. 1999, 116, 281–287. [Google Scholar] [CrossRef]
- Klein, C.; Troedsson, M.H.T. Maternal recognition of pregnancy in the horse: A mystery still to be solved. Reprod. Fertil. Dev. 2011, 23, 952–963. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter-Villani, M.; Van Tol, H.T.A.; Stout, T.A.E. Effect of pregnancy on endometrial expression of luteolytic pathway components in the mare. Reprod. Fertil. Dev. 2015, 27, 834–845. [Google Scholar] [CrossRef]
- Merkl, M.; Ulbrich, S.E.; Otzdorff, C.; Herbach, N.; Wanke, R.; Wolf, E.; Handler, J.; Bauersachs, S. Microarray analysis of equine endometrium at days 8 and 12 of pregnancy. Biol. Reprod. 2010, 83, 874–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauersachs, S.; Wolf, E. Transcriptome analyses of bovine, porcine and equine endometrium during the pre-implantation phase. Anim. Reprod. Sci. 2012, 134, 84–94. [Google Scholar] [CrossRef]
- Klohonatz, K.M.; Cloeman, S.J.; Islas-Trejo, A.D.; Medrano, J.F.; Hess, A.M.; Kalbfleisch, T.; Thomas, M.G.; Bouma, G.J.; Breummer, J.E. Coding RNA sequencing of equine endometrium during maternal recognition of pregnancy. Genes 2019, 10, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klohonatz, K.M.; Hess, A.M.; Hansen, T.R.; Squires, E.L.; Bouma, G.J.; Bruemmer, J.E. Equine endometrial gene expression changes during and after maternal recognition of pregnancy. J. Anim. Sci. 2015, 93, 3364–3376. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Scoggin, K.E.; Ealy, A.D.; Troedsson, M.H.T.T. Transcriptional profiling of equine endometrium during the time of maternal recognition of pregnancy. Biol. Reprod. 2010, 83, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Klein, C.; Troedsson, M.H.T. Transcriptional profiling of equine conceptuses reveals new aspects of embryo-maternal communication in the horse. Biol. Reprod. 2011, 84, 872–885. [Google Scholar] [CrossRef] [Green Version]
- Klein, C. Novel equine conceptus-endometrial interactions on Day 16 of pregnancy based on RNA sequencing. Reprod. Fertil. Dev. 2015, 28, 1712–1720. [Google Scholar] [CrossRef]
- Swegen, A.; Grupen, C.G.; Gibb, Z.; Baker, M.A.; de Ruijter-Villani, M.; Smith, N.D.; Stout, T.A.E.; Aitken, R.J. From peptide masses to pregnancy maintenance: A comprehensive proteomic analysis of the early euine embryo secretome, blastocoel fluid, and capsule. Proteomics 2017, 17, 1600433. [Google Scholar] [CrossRef] [PubMed]
- Smits, K.; Willems, S.; Van Steendam, K.; Van De Velde, M.; De Lange, V.; Ververs, C.; Roels, K.; Govaere, J.; Van Nieuwerburgh, F.; Peelman, L.; et al. Proteins involved in embryo-maternal interaction around the signalling of maternal recognition of pregnancy in the horse. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Bazer, F.W.; Spencer, T.E.; Johnson, G.A.; Burghardt, R.C.; Wu, G. Comparative aspects of implantation. Reproduction 2009, 138, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Filant, J.; Spencer, T.E. Uterine glands: Biological roles in conceptus implantation, uterine receptivity and decidualization. Int. J. Dev. Biol. 2014, 58, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P.; Forde, N. The role of pogesterone in maternal recognition of pregnancy in domestic ruminants. In Regulation of Implantation and Establishment of Pregnancy in Mammals. Tribute to 45 Year Anniversary of Roger V. Short’s “Maternal Recognition of Pregnancy”; Geisert, D.R., Bazer, F.W., Eds.; Springer International Publishing: Basel, Switzerland, 2015; pp. 87–104. [Google Scholar]
- Satterfield, M.C.; Hayashi, K.; Song, G.; Black, S.G.; Bazer, F.W.; Spencer, T.E. Progesterone regulates FGF10, MET, IGFBP1, and IGFBP3 in the endometrium of the ovine uterus. Biol. Reprod. 2008, 79, 1226–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forde, N.; Carter, F.; Fair, T.; Crowe, M.A.; Evans, A.C.O.; Spencer, T.E.; Bazer, F.W.; McBride, R.; Boland, M.P.; O’Gaora, P.; et al. Progesterone-regulated changes in endometrial gene expression contribute to advanced conceptus development in cattle. Biol. Reprod. 2009, 81, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Forde, N.; Lonergan, P. The role of progesterone and conceptus-derived factors in uterine biology during early pregnancy in ruminants. J. Dairy Sci. 2016, 99, 5941–5950. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.R.; Mathias, S.; Lennard, S.N.; Greenwood, R.E. Serial measurement of peripheral oestrogen and progesterone concentrations in oestrous mares to determine optimum mating time and diagnose ovulation. Equine Vet. J. 1995, 27, 460–464. [Google Scholar] [CrossRef]
- Mann, G.E.; Lamming, G.E. Relationship between maternal endocrine environment, early embryo development and inhibition of the luteolytic mechanism in cows. Reproduction 2001, 121, 175–180. [Google Scholar] [CrossRef]
- Willman, C.; Budik, S.; Walter, I.; Aurich, C. Influences of treatment of early pregnant mares with the progestin altrenogest on embryonic development and gene expression in the endometrium and conceptus. Theriogenology 2011, 76, 61–73. [Google Scholar] [CrossRef]
- Beyer, T.; Rink, B.E.; Scarlet, D.; Walter, I.; Kunert, S.; Aurich, C. Early luteal phase progestin concentration influences endometrial function in pregnant mares. Theriogenology 2019, 125, 236–241. [Google Scholar] [CrossRef]
- Randi, F.; Fernandez-Fuertes, B.; McDonald, M.; Forde, N.; Kelly, A.K.; Bastos Amorin, H.; Muniz de Lima, E.; Morotti, F.; Marcondes Seneda, M.; Lonergan, P. Asynchronous embryo transfer as a tool to understand embryo–uterine interaction in cattle: Is a large conceptus a good thing? Reprod. Fertil. Dev. 2016, 28, 1999–2006. [Google Scholar] [CrossRef]
- Geisert, R.D.; Fox, T.C.; Morgan, G.L.; Wells, M.E.; Wettemann, R.P.; Zavy, M.T. Survival of bovine embryos transferred to progesterone-treated asynchronous recipients. Reproduction 1991, 92, 475–482. [Google Scholar] [CrossRef]
- Ledgard, A.M.; Berg, M.C.; McMillan, W.H.; Smolenski, G.; Peterson, A.J. Effect of asynchronous transfer on bovine embryonic development and relationship with early cycle uterine proteome profiles. Reprod. Fertil. Dev. 2012, 24, 962–972. [Google Scholar] [CrossRef]
- Wilsher, S.; Clutton-Brock, A.; Allen, W.R. Successful transfer of day 10 horse embryos: Influence of donor-recipient asynchrony on embryo development. Reproduction 2010, 139, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.C.F.; Haag, K.T.; Santos, G.O.; Oliveira, J.P.; Gastal, M.O.; Gastal, E.L. Effect of embryo age and recipient asynchrony on pregnancy rates in a commercial equine embryo transfer program. Theriogenology 2012, 77, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Cuervo-Arango, J.; Claes, A.N.; Stout, T.A.E. In vitro produced horse embryos exhibit a very narrow window of acceptable recipient mare uterine synchrony compared with in vivo derived embryos. Reprod. Fertil. Dev. 2019, 31, 1904–1911. [Google Scholar] [CrossRef] [PubMed]
- Wilsher, S.; Allen, W.R. Uterine influences on embryogenesis and early placentation in the horse revealed by transfer of day 10 embryos to day 3 recipient mares. Reproduction 2009, 137, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Gibson, C.; de Ruijter-Villani, M.; . Stout, T.A.E. Negative uterine asynchrony retards early equine conceptus development and upregulation of placental imprinted genes. Placenta 2017, 57, 175–182. [Google Scholar] [CrossRef]
- Gibson, C.; de Ruijter-Villani, M.; Rietveld, J.; Stout, T.A.E. Amino acid transporter expression in the endometrium and conceptus membranes during early equine pregnancy. Reprod. Fertil. Dev. 2018, 30, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Gibson, C.; de Ruijter-Villani, M.; Rietveld, J.; Stout, T.A.E. Expression of glucose transporters in the endometrium and early conceptus membranes of the horse. Placenta 2018, 68, 22–32. [Google Scholar] [CrossRef]
- Burns, G.W.; Brooks, K.E.; Spencer, T.E. Extracellular vesicles originate from the conceptus and uterus during early pregnancy in the sheep. Biol. Reprod. 2016, 94, 56. [Google Scholar] [CrossRef] [Green Version]
- Burns, G.W.; Brooks, K.E.; O’Neil, E.V.; Hagen, D.E.; Behura, S.K.; Spencer, T.E. Progesterone effects on extracellular vesicles in th esheep uterus. Biol. Reprod. 2018, 98, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Quinn, B.A.; Hayes, M.A.; Waelchi, R.O.; Kennedy, M.W.; Betteridge, K.J. Changes in major proteins in the embryonic capsule during immobilization (fixtaion) of the conceptus in the third week of pregnancy in the mare. Reproduction 2007, 134, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Crossett, B.; Allen, W.R.; Stewart, F. A 19 kDa protein secreted by the endometrium of the mare is a novel member of the lipocalin family. Biochem. J. 1996, 320, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, W.R.; Gower, S.; Wilsher, S. Immunohistochemical localization of vascular endothelial growth factor (VEGF) and its two receptors (Flt-I and KDR) in the endometrium and placenta of the mare during the oestrous cycle and pregnancy. Reprod. Domest. Anim. 2007, 42, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E. Lipid metabolism in pregnancy and its consequences in the fetus and newborn. Endocrine 2002, 19, 43–55. [Google Scholar] [CrossRef]
- Suire, S.; Stewart, F.; Beauchamp, J.; Kennedy, M.W. Uterocalin, a lipocalin provisioning the preattachment equine conceptus: Fatty acid and retinol binding properties, and structural characterization. Biochem. J. 2001, 356, 369–376. [Google Scholar] [CrossRef]
- Bourdiec, A.; Ahmad, S.F.; Lachhab, A.; Akoum, A. Regulation of inflammatory and angiogenesis mediators in a functional model of decidualized endometrial stromal cells. Reprod. Biomed. Online 2016, 32, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Vonnahme, K.A.; Fernando, S.C.; Ross, J.W.; Ashworth, M.D.; DeSilva, U.; Malayer, J.R.; Geisert, R.D. Porcine endometrial expression of kininogen, factor XII, and plasma kallikrein in cyclic and pregnant gilts. Biol. Reprod. 2004, 70, 132–138. [Google Scholar] [CrossRef]
- Golias, C.; Charalabopoulos, A.; Stagikas, D.; Charalabopoulos, K.; Batistatou, A. The kinin system–bradykinin: Biological effects and clinical implications. Multiple role of the kinin system–bradykinin., Hippokratia 2007, 11, 124–128. [Google Scholar]
- Hillmeister, P.; Persson, P.B. The Kallikrein-Kinin system. Acta Physiol. 2012, 206, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.R.; Zhang, B.R.; Hettinger, A.M.; Goad, D.W.; Malayer, J.R.; Geisert, R.D. Detection of bradykinin and bradykinin-beta(2) receptors in the porcine endometrium during the estrous cycle and early pregnancy. Biol. Reprod. 2002, 574–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawato, H.; Tabata, T.; Minoura, H.; Murabayashi, N.; Ma, N.; Fang Wang, D.; Sagawa, N. Factor XII gene expression in endometrial stromal cells during decidualisation. Reprod. Fertil. Dev. 2009, 21, 840–847. [Google Scholar] [CrossRef]
- Ullah, K.; Rahman, T.U.; Pan, H.T.; Guo, M.X.; Dong, X.Y.; Liu, J.; Jin, L.Y.; Cheng, Y.; Ke, Z.H.; Ren, J.; et al. Serum estradiol levels in controlled ovarian stimulation directly affect the endometrium. J. Mol. Endocrinol. 2017, 59, 105–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erices, R.; Corthorn, J.; Lisboa, F.; Valdés, G. Bradykinin promotes migration and invasion of human immortalized trophoblasts. Reprod. Biol. Endocrinol. 2011, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- Stout, T.A.; Allen, W.R. Role of prostaglandins in intrauterine migration of the equine conceptus. Reproduction 2001, 121, 771–775. [Google Scholar] [CrossRef]
- Morrissey, N.K.; Bellenger, C.R.; Baird, A.W. Bradykinin stimulates prostaglandin E2 production and cyclooxygenase activity in equine nonglandular and glandular gastric mucosa in vitro. Equine Vet. J. 2008, 40, 332–336. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Espey, L.; Hosoi, Y.; Adachi, T.S.; Atlas, J.; Ghodgaonkar, R.B.; Dubin, N.H.; Wallach, E.E. The Effects of Bradykinin on Ovulation and Prostaglandin Production by the Perfused Rabbit Ovary. Endocrinology 1988, 122, 2540–2546. [Google Scholar] [CrossRef]
- Hellberg, P.; Larson, L.; Olofsson, J.; Hedin, L.; Brannstrom, M. Stimulatory effects of bradykinin on the ovulatory process in the in vitro-perfused rat ovary. Biol Reprod. 1991, 44, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Nie, M.; Pang, L.; Inoue, H.; Knox, A.J. Transcriptional regulation of cyclooxygenase 2 by bradykinin and interleukin-1 beta in human airway smooth muscle cells: Involvement of different promoter elements, transcription factors, and histone h4 acetylation. Mol. Cell. Biol. 2003, 23, 9233–9244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, D.A.; Newton, R.; Zhu, Y.M.; El-Haroun, H.; Corbett, L.; Knox, A.J. Cyclooxygenase-2 induction by bradykinin in human pulmonary artery smooth muscle cells is mediated by the cyclic AMP response element through a novel autocrine loop involving endogenous prostaglandin E2, E-prostanoid 2 (EP2), and EP4 Receptors. J. Biol. Chem. 2003, 278, 49954–49964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorniak, P.; Bazer, F.W.; Wu, G.; Spencer, T.E. Conceptus-derived prostaglandins regulate endometrial function in sheep. Biol. Reprod. 2012, 87, 1–7. [Google Scholar]
- Simmons, R.M.; Erikson, D.W.; Kim, J.; Burghardt, R.C.; Bazer, F.W.; Johnson, G.A.; Spencer, T.E. Insulin-like growth factor binding protein-1 in the ruminant uterus: Potential endometrial marker and regulator of conceptus elongation. Endocrinology 2009, 150, 4295–4305. [Google Scholar] [CrossRef]
- Robinson, R. The expression of the IGF system in the bovine uterus throughout the oestrous cycle and early pregnancy. J. Endocrinol. 2000, 165, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Herrler, A.; Pell, J.M.; Allen, W.R.; Beier, H.M.; Stewart, F. Horse conceptuses secrete insulin-like growth factor-binding protein 3. Biol. Reprod. 2000, 62, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.W.; Roser, J.F.; Anderson, G.B. Maternal-conceptus signaling during early pregnancy in mares: Oestrogen and insulin-like growth factor 1. Reproduction 2001, 121, 331–338. [Google Scholar] [CrossRef]
- Forbes, K.; Souquet, B.; Garside, R.; Aplin, J.D.; Westwood, M. Transforming growth factor-β (TGFβ) receptors I/II differentially regulate TGFβ1 and IGF-binding protein-3 mitogenic effects in the human placenta. Endocrinology 2010, 151, 1723–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, R.C. Insulin-like growth factor binding protein-3 (IGFBP-3): Novel ligands mediate unexpected functions. J. Cell Commun. Signal. 2013, 7, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fong, K.W.; Tang, M.; Han, X.; Gong, Z.; Ma, W.; Hebert, M.; Songyang, Z.; Chen, J. Fam118B, a newly identified component of Cajal bodies, is required for Cajal body formation, snRNP biogenesis and cell viability. J. Cell Sci. 2014, 127, 2029–2039. [Google Scholar] [CrossRef] [Green Version]
- de Ruijter-Villani, M.; van Boxtel, P.R.M.; Stout, T.A.E. Fibroblast growth factor-2 expression in the preimplantation equine conceptus and endometrium of pregnant and cyclic mares. Theriogenology 2013, 80, 979–989. [Google Scholar] [CrossRef]
- Stout, T.A.E. Equine embryo transfer: Review of developing potential. Equine Vet. J. 2006, 38, 467–478. [Google Scholar] [CrossRef]
- Blankenberg, D.; Von Kuster, G.; Coraor, N.; Ananda, G.; Mangan, M.; Nekrutenko, A.; Taylor, J. Galaxy: A web-based genome analysis tool for experimentalists. Cuur. Protoc. Mol. Biol. 2010, 19, 10. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaidatzis, D.; Lerch, A.; Hahne, F.; Stadler, M.B. QuasR: Quantification and annotation of short reads in R. Bioinformatics 2015, 31, 1130–1132. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- De Grauw, J.C.; Van De Lest, C.H.A.; van Weeren, P.R. A targeted lipidomics approach to the study of eicosanoid release in synovial joints. Arthritis Res. Ther. 2011, 13, R123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cokelaere, S.M.; Plomp, S.G.M.; de Boef, E.; de Leeuw, M.; Bool, S.; van de Lest, C.H.A.; van Weeren, P.R.; Korthagen, N.M. Sustained intra-articular release of celecoxib in an equine repeated LPS synovitis model. Eur. J. Pharm. Biopharm. 2018, 128, 327–336. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endometrium | DEGs | Upregulated | Downregulated |
|---|---|---|---|
| D14–Syn/Asyn | 523 | 174 | 349 |
| D19–Syn/Asyn | 715 | 283 | 432 |
| D14–Syn/D19–Asyn | 14 | 7 | 7 |
| Conceptus | |||
| D14–Syn/Asyn | 556 | 285 | 271 |
| D19–Syn/Asyn | 3473 | 1870 | 1603 |
| D14–Syn/D19–Asyn | 3646 | 1961 | 1685 |
| Gene Name | Gene Symbol | D14 RNA-seq | D14 PCR | D19 RNA-seq | D19 PCR | ||||
|---|---|---|---|---|---|---|---|---|---|
| Log2 FC | P-Adjusted | Log2 FC | p-Value | Log2 FC | P-Adjusted | Log2 FC | p-Value | ||
| Interleukin 6 signal transducer | IL6ST | 0.33 | 0.572 | 0.77 | 0.249 | 0.65 | <0.001 | 0.27 | 0.283 |
| Insulin receptor | INSR | 0.96 | 0.009 | 0.58 | 0.291 | 0.99 | <0.001 | 0.69 | 0.065 |
| Leukemia inhibitory factor (LIF) | LIF | 1.35 | 0.022 | 1.37 | 0.082 | 2.57 | <0.001 | 3.07 | 0.001 |
| LIF receptor | LIFR | 0.27 | 0.419 | 1.22 | 0.148 | −0.20 | 0.639 | −0.44 | 0.879 |
| Oxytocin receptor | OXTR | 1.51 | 0.007 | 0.74 | 0.226 | 0.63 | 0.071 | 0.04 | 0.711 |
| Prostaglandin F receptor | PTGFR | −1.47 | <0.001 | −1.83 | 0.016 | −0.47 | 0.161 | −1.46 | 0.153 |
| SLC family 1, member 1 | SLC1A1 | 0.08 | 0.962 | −0.02 | 0.919 | −1.49 | <0.001 | −1.75 | 0.002 |
| SLC family 2, member 1 | SLC2A1 | 1.89 | <0.001 | 3.16 | 0.001 | 0.11 | 0.889 | −1.32 | 0.379 |
| SLC family 38, member 2 | SLC38A2 | 0.36 | 0.283 | 0.09 | 0.636 | 1.10 | <0.001 | 0.75 | 0.061 |
| SLC family 43, member 2 | SLC43A2 | −0.47 | 0.425 | −1.22 | 0.050 | −0.33 | 0.450 | −0.98 | 0.173 |
| SLC family 5, member 1 | SLC5A1 | −0.28 | 0.752 | −0.36 | 0.492 | −0.08 | 0.935 | −0.90 | 0.369 |
| Gene Name | Gene Symbol | D14 RNA-seq | D14 PCR | D19 RNA-seq | D19 PCR | ||||
|---|---|---|---|---|---|---|---|---|---|
| Log2 FC | P-Adjusted | Log2 FC | p-Value | Log2 FC | P-Adjusted | Log2 FC | p-Value | ||
| DNA-methyltransferase 3B | DNMT3B | −0.71 | <0.001 | −0.64 | 0.048 | 0.12 | 0.613 | 0.66 | 0.018 |
| H19, Imprinted maternally expressed transcript | H19 | 0.03 | 0.968 | 0.11 | 0.572 | 0.40 | 0.083 | 0.62 | 0.058 |
| Insulin-like growth factor 1 | IGF1 | 0.43 | 0.302 | 2.20 | 0.007 | 1.48 | 0.027 | 1.67 | 0.082 |
| Necdin | NDN | −0.17 | 0.729 | −0.53 | 0.405 | 1.42 | 0.001 | 1.57 | 0.019 |
| Paternally expressed gene 10 | PEG10 | 0.32 | 0.441 | 0.45 | 0.522 | 2.70 | <0.001 | 2.89 | 0.004 |
| SLC family 1, member 4 | SLC1A4 | 0.79 | 0.008 | 1.10 | 0.002 | 0.72 | 0.003 | 0.77 | 0.062 |
| SLC family 1, member 5 | SLC1A5 | −1.65 | <0.001 | −2.52 | 0.004 | 2.15 | <0.001 | 3.07 | 0.029 |
| SLC family 2, member 3 | SLC2A3 | 0.22 | 0.305 | 0.54 | 0.167 | −0.16 | 0.200 | −0.02 | 0.680 |
| SLC family 2, member 5 | SLC2A5 | 0.49 | 0.012 | 1.55 | 0.002 | −1.20 | <0.001 | −1.42 | 0.001 |
| SLC family 7, member 5 | SLC7A5 | −0.04 | 0.941 | 0.69 | 0.464 | 1.64 | <0.001 | 2.52 | 0.007 |
| Small nuclear ribonucleoprotein-associated protein | SNRPN | −0.04 | 0.914 | −0.03 | 0.772 | −0.05 | 0.866 | −0.40 | 0.542 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibson, C.; de Ruijter-Villani, M.; Bauersachs, S.; Stout, T.A.E. Asynchronous Embryo Transfer Followed by Comparative Transcriptomic Analysis of Conceptus Membranes and Endometrium Identifies Processes Important to the Establishment of Equine Pregnancy. Int. J. Mol. Sci. 2020, 21, 2562. https://doi.org/10.3390/ijms21072562
Gibson C, de Ruijter-Villani M, Bauersachs S, Stout TAE. Asynchronous Embryo Transfer Followed by Comparative Transcriptomic Analysis of Conceptus Membranes and Endometrium Identifies Processes Important to the Establishment of Equine Pregnancy. International Journal of Molecular Sciences. 2020; 21(7):2562. https://doi.org/10.3390/ijms21072562
Chicago/Turabian StyleGibson, Charlotte, Marta de Ruijter-Villani, Stefan Bauersachs, and Tom A.E. Stout. 2020. "Asynchronous Embryo Transfer Followed by Comparative Transcriptomic Analysis of Conceptus Membranes and Endometrium Identifies Processes Important to the Establishment of Equine Pregnancy" International Journal of Molecular Sciences 21, no. 7: 2562. https://doi.org/10.3390/ijms21072562
APA StyleGibson, C., de Ruijter-Villani, M., Bauersachs, S., & Stout, T. A. E. (2020). Asynchronous Embryo Transfer Followed by Comparative Transcriptomic Analysis of Conceptus Membranes and Endometrium Identifies Processes Important to the Establishment of Equine Pregnancy. International Journal of Molecular Sciences, 21(7), 2562. https://doi.org/10.3390/ijms21072562