All-trans Retinoic Acid-induced Abnormal Hippocampal Expression of Synaptic Genes SynDIG1 and DLG2 is Correlated with Anxiety or Depression-Like Behavior in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
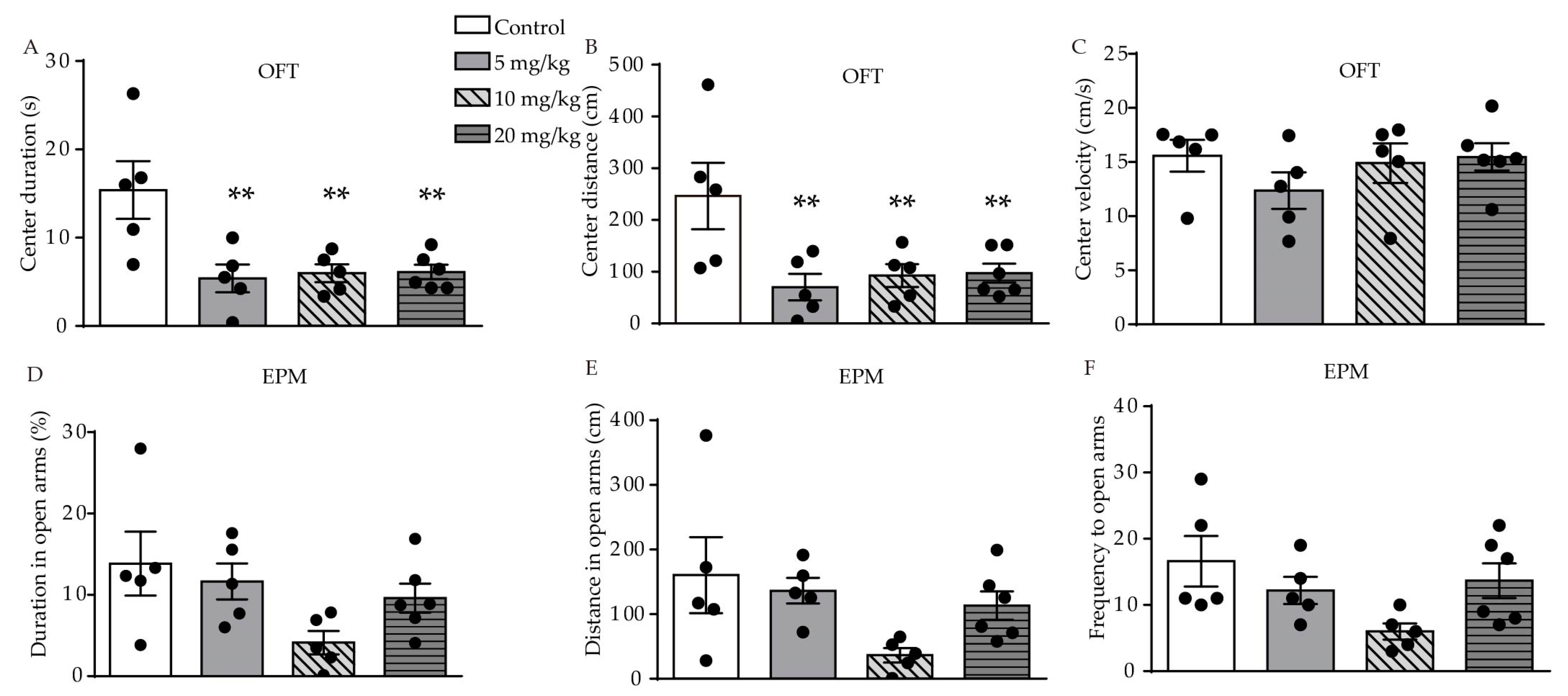
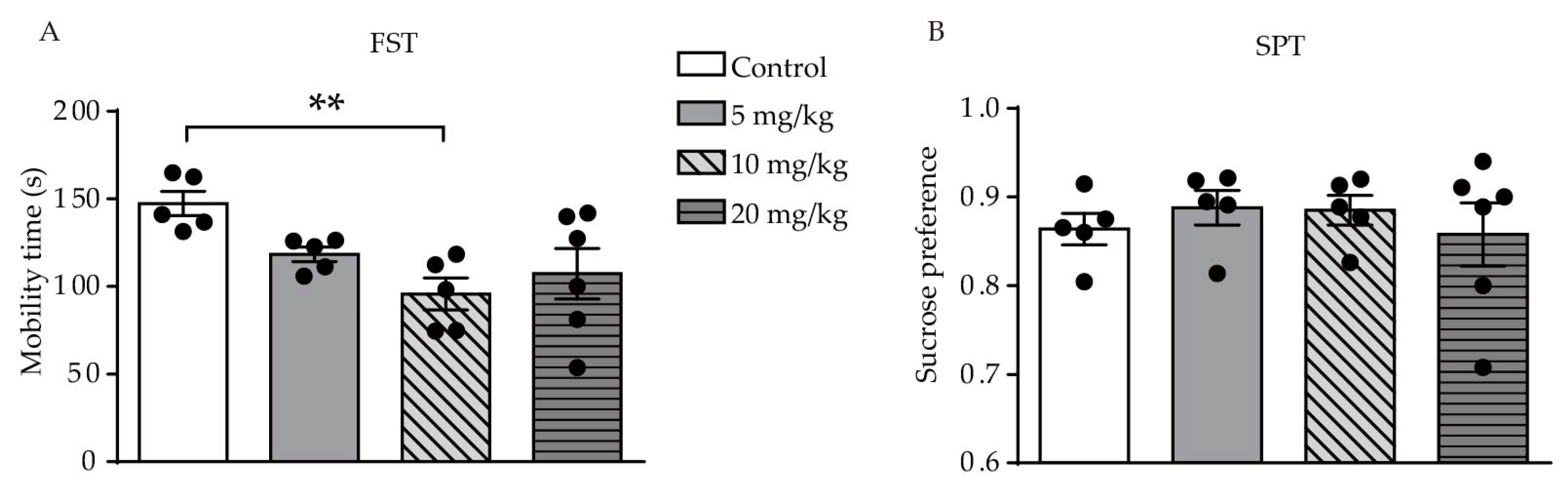
2.1. ATRA-Induced Anxiety- and Depression-Like Behavior in Young Mice
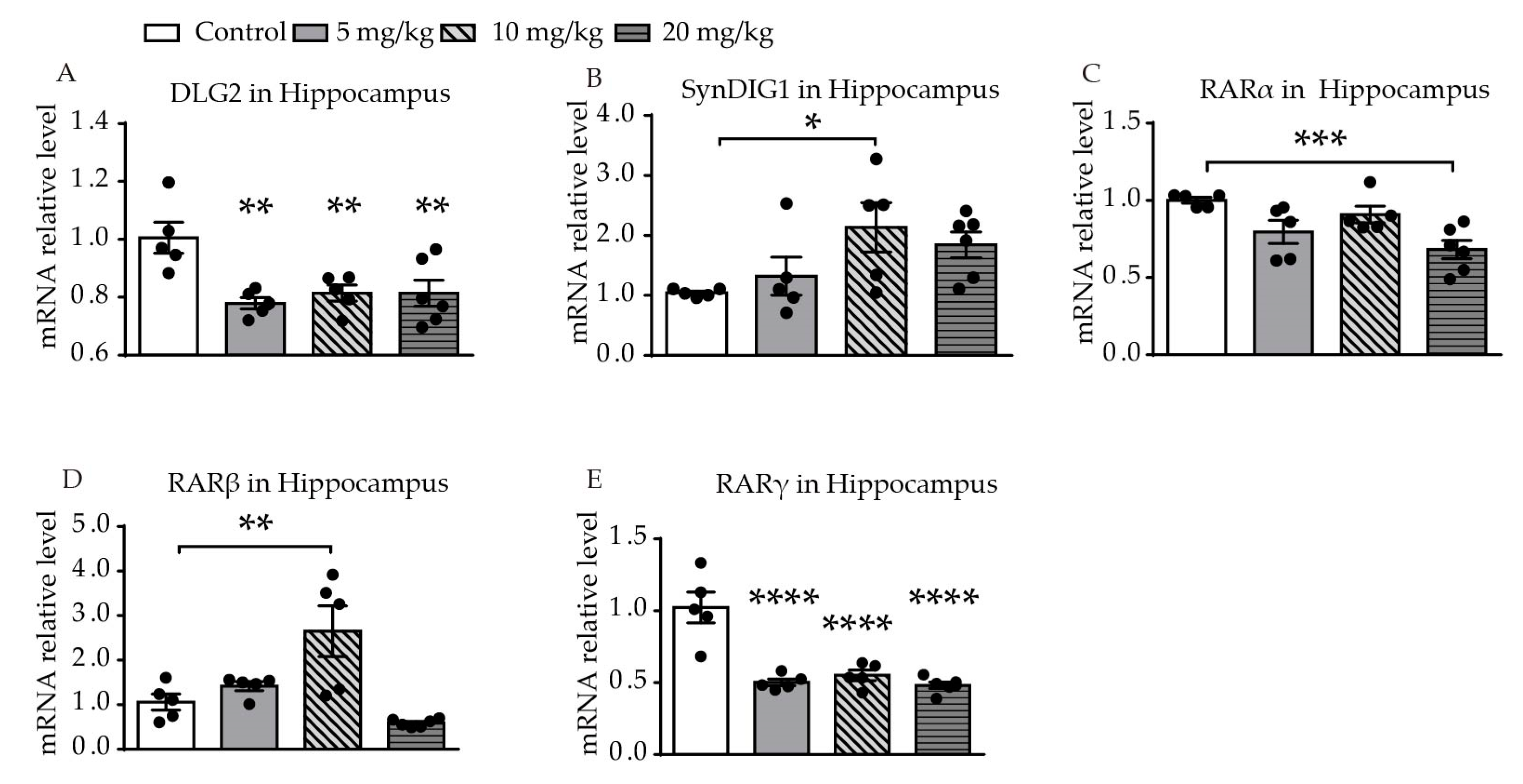
2.2. ATRA-Induced Changes in mRNA Expression of DLG2, SynDIG1, and Retinoic Acid Receptors in the Hippocampus
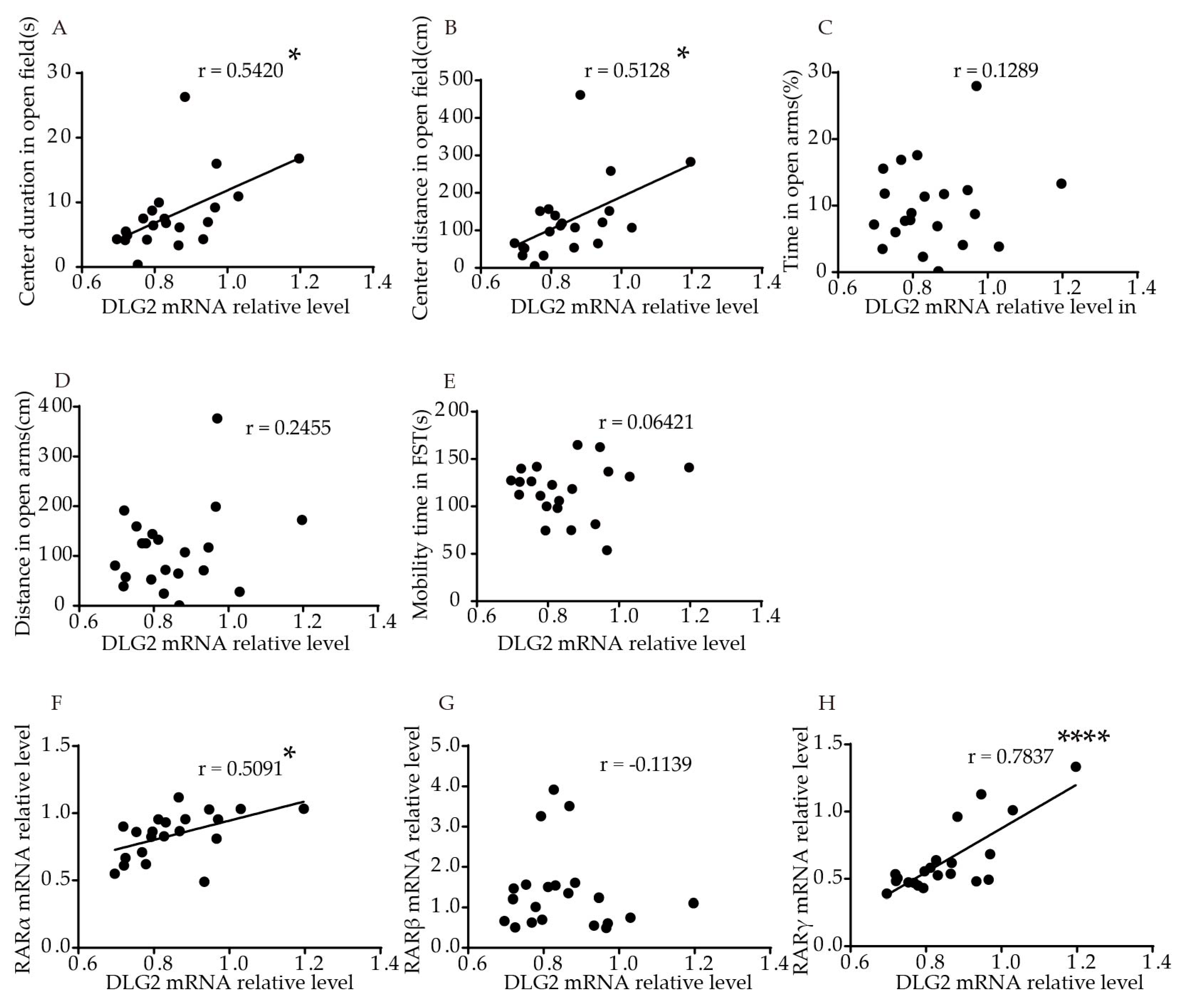
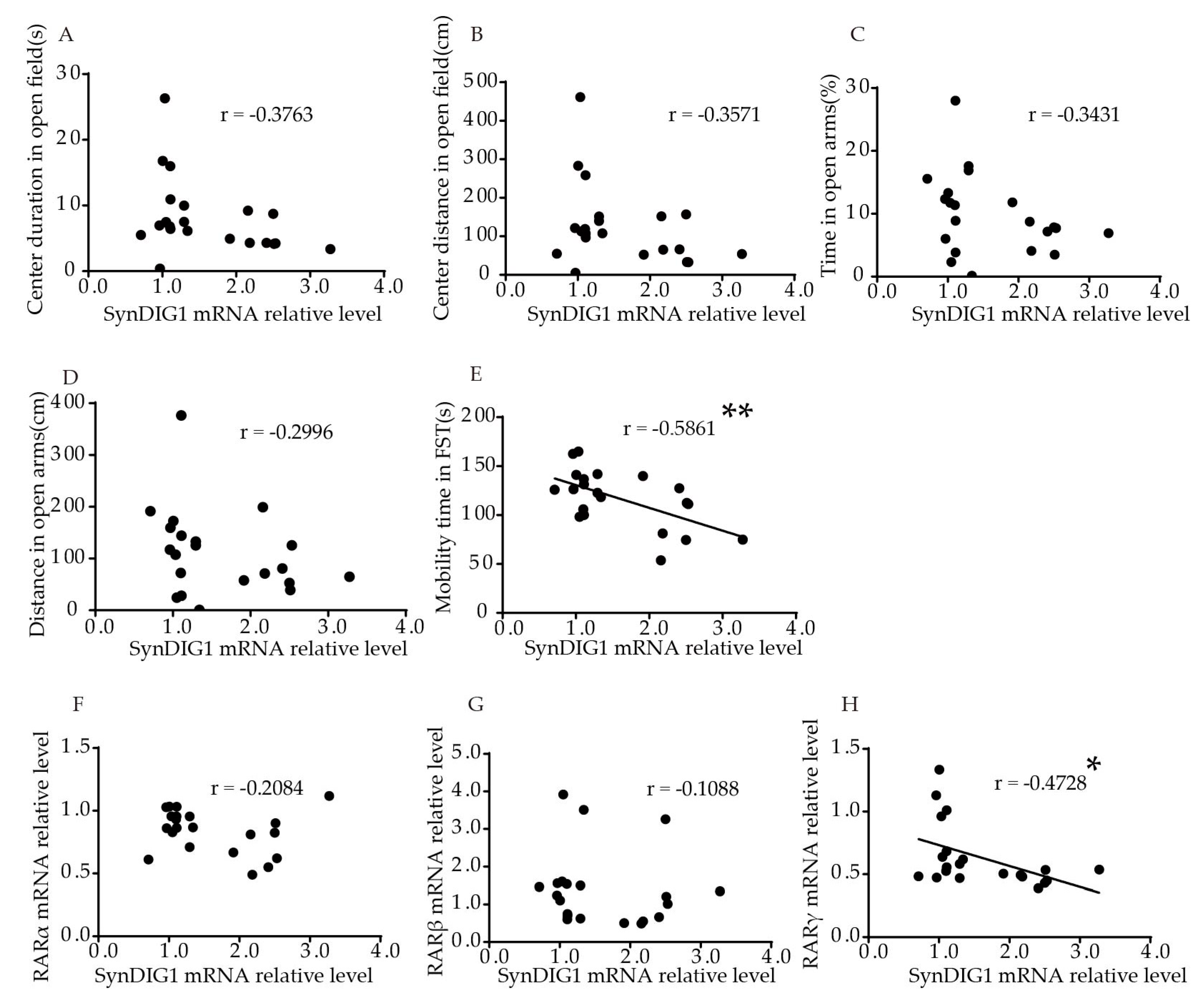
2.3. Association of DLG2 and SynDIG1 mRNA Levels with Anxiety- and Depression-Like Behavior and RARs in the Hippocampus
3. Discussion
4. Materials and Methods
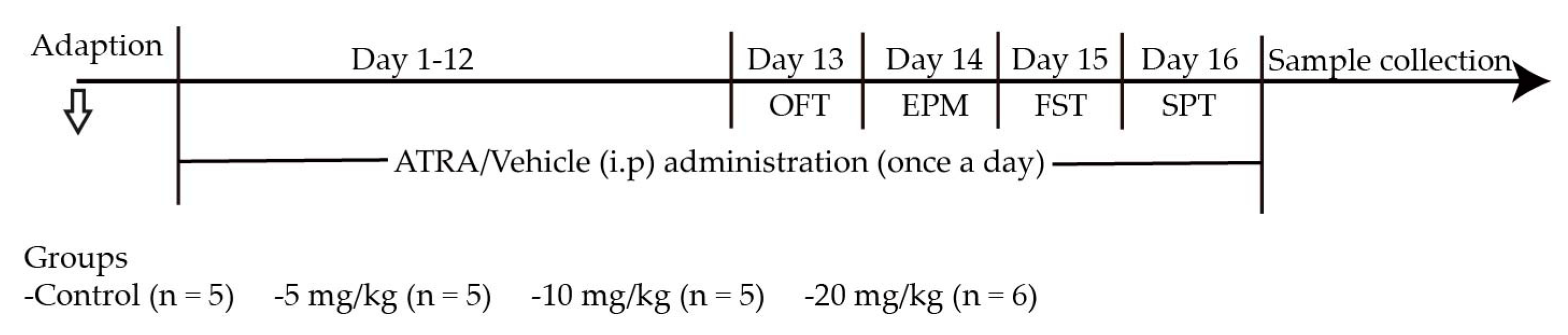
4.1. Animals and Drugs
4.2. Behavioral Tests
4.2.1. Open-Field Test (OFT)
4.2.2. Elevated-Plus Maze Test (EPM)
4.2.3. Forced Swimming Test (FST)
4.2.4. Sucrose Consumption Test (SPT)
4.3. Tissue Preparation
4.4. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPAR | α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid subtype glutamate receptors |
| ATRA | all-trans retinoic acid |
| CNS | central nervous system |
| DLG2 | discs large homolog 2 |
| EPM | elevated-plus maze |
| FST | forced swimming test |
| HPA | hypothalamic-pituitary-adrenal axis |
| LSD | least significant difference |
| NMDAR | N-methyl-D-aspartate receptor |
| OFT | open-field test |
| RARα | retinoic acid receptor α |
| RARβ | retinoic acid receptor β |
| RARγ | retinoic acid receptor γ |
| SPT | sucrose preference test |
| SynDIG1 | synapse differentiation-inducing gene protein 1 |
References
- Cocco, S.; Diaz, G.; Stancampiano, R.; Diana, A.; Carta, M.; Curreli, R.; Sarais, L.; Fadda, F. Vitamin A deficiency produces spatial learning and memory impairment in rats. Neuroscience 2002, 115, 475–482. [Google Scholar] [CrossRef]
- Bremner, J.D.; Shearer, K.D.; McCaffery, P.J. Retinoic acid and affective disorders: The evidence for an association. J. Clin. Psychiatry 2012, 73, 37–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremner, J.D.; McCaffery, P. The neurobiology of retinoic acid in affective disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 315–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafti, M.; Ghyselinck, N.B. Functional implication of the vitamin A signaling pathway in the brain. Arch. Neurol. 2007, 64, 1706–1711. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J. Polar hysteria: An expression of hypervitaminosis A. Am. J. Ther. 2004, 11, 507–516. [Google Scholar] [CrossRef]
- Hsu, P.; Litman, G.I.; Brodell, R.T. Overview of the treatment of acne vulgaris with topical retinoids. Postgrad. Med. 2011, 123, 153–161. [Google Scholar] [CrossRef]
- Jacobs, S.; Lie, D.C.; DeCicco, K.L.; Shi, Y.; DeLuca, L.M.; Gage, F.H.; Evans, R.M. Retinoic acid is required early during adult neurogenesis in the dentate gyrus. Proc. Natl. Acad. Sci. USA 2006, 103, 3902–3907. [Google Scholar] [CrossRef] [Green Version]
- Werner, E.A.; DeLuca, H.F. Retinoic acid is detected at relatively high levels in the CNS of adult rats. Am. J. Physiol.-Endoc. M 2002, 282, E672–E678. [Google Scholar] [CrossRef] [Green Version]
- Maden, M. Role and distribution of retinoic acid during CNS development. Int. Rev. Cytol. 2001, 209, 1–77. [Google Scholar]
- Cai, L.; Yan, X.B.; Chen, X.N.; Meng, Q.Y.; Zhou, J.N. Chronic all-trans retinoic acid administration induced hyperactivity of HPA axis and behavioral changes in young rats. Eur. Neuropsychopharmacol. 2010, 20, 839–847. [Google Scholar] [CrossRef]
- Hu, P.; Wang, Y.; Liu, J.; Meng, F.T.; Qi, X.R.; Chen, L.; van Dam, A.M.; Joels, M.; Lucassen, P.J.; Zhou, J.N. Chronic retinoic acid treatment suppresses adult hippocampal neurogenesis, in close correlation with depressive-like behavior. Hippocampus 2016, 26, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.N.; Meng, Q.Y.; Bao, A.M.; Swaab, D.F.; Wang, G.H.; Zhou, J.N. The involvement of retinoic acid receptor-alpha in corticotropin-releasing hormone gene expression and affective disorders. Biol. Psychiatry 2009, 66, 832–839. [Google Scholar] [CrossRef] [PubMed]
- MacQueen, G.M.; Campbell, S.; McEwen, B.S.; Macdonald, K.; Amano, S.; Joffe, R.T.; Nahmias, C.; Young, L.T. Course of illness, hippocampal function, and hippocampal volume in major depression. Proc. Natl. Acad. Sci. USA 2003, 100, 1387–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frodl, T.; Meisenzahl, E.M.; Zetzsche, T.; Born, C.; Groll, C.; Jager, M.; Leinsinger, G.; Bottlender, R.; Hahn, K.; Moller, H.J. Hippocampal changes in patients with a first episode of major depression. Am. J. Psychiatry 2002, 159, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Neumeister, A.; Wood, S.; Bonne, O.; Nugent, A.C.; Luckenbaugh, D.A.; Young, T.; Bain, E.E.; Charney, D.S.; Drevets, W.C. Reduced hippocampal volume in unmedicated, remitted patients with major depression versus control subjects. Biol. Psychiatry 2005, 57, 935–937. [Google Scholar] [CrossRef] [PubMed]
- Stockmeier, C.A.; Mahajan, G.J.; Konick, L.C.; Overholser, J.C.; Jurjus, G.J.; Meltzer, H.Y.; Uylings, H.B.M.; Friedman, L.; Rajkowska, G. Cellular changes in the postmortem hippocampus in major depression. Biol. Psychiatry 2004, 56, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Masi, G.; Brovedani, P. The hippocampus, neurotrophic factors and depression. CNS Drugs 2011, 25, 913–931. [Google Scholar] [CrossRef]
- Hajszan, T.; Dow, A.; Warner-Schmidt, J.L.; Szigeti-Buck, K.; Sallam, N.L.; Parducz, A.; Leranth, C.; Duman, R.S. Remodeling of hippocampal spine synapses in the rat learned helplessness model of depression. Biol. Psychiatry 2009, 65, 392–400. [Google Scholar] [CrossRef] [Green Version]
- Hajszan, T.; Szigeti-Buck, K.; Sallam, N.L.; Bober, J.; Parducz, A.; Maclusky, N.J.; Leranth, C.; Duman, R.S. Effects of estradiol on learned helplessness and associated remodeling of hippocampal spine synapses in female rats. Biol. Psychiatry 2010, 67, 168–174. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, M.R.; Silvestrin, R.B.; Mello, E.S.T.; Moreira, J.C. Oxidative stress in the hippocampus, anxiety-like behavior and decreased locomotory and exploratory activity of adult rats: Effects of sub acute vitamin A supplementation at therapeutic doses. Neurotoxicology 2007, 28, 1191–1199. [Google Scholar] [CrossRef]
- Aoto, J.; Nam, C.I.; Poon, M.M.; Ting, P.; Chen, L. Synaptic Signaling by All-Trans Retinoic Acid in Homeostatic Synaptic Plasticity. Neuron 2008, 60, 308–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarti, F.; Zhang, Z.; Schroeder, J.; Chen, L. Rapid suppression of inhibitory synaptic transmission by retinoic acid. J. Neurosci. 2013, 33, 11440–11450. [Google Scholar] [CrossRef] [PubMed]
- Maghsoodi, B.; Poon, M.M.; Nam, C.I.; Aoto, J.; Ting, P.; Chen, L. Retinoic acid regulates RAR alpha-mediated control of translation in dendritic RNA granules during homeostatic synaptic plasticity. Proc. Natl. Acad. Sci. USA 2008, 105, 16015–16020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlisle, H.J.; Fink, A.E.; Grant, S.G.; O’Dell, T.J. Opposing effects of PSD-93 and PSD-95 on long-term potentiation and spike timing-dependent plasticity. J. Physiol. 2008, 586, 5885–5900. [Google Scholar] [CrossRef]
- Duric, V.; Banasr, M.; Stockmeier, C.A.; Simen, A.A.; Newton, S.S.; Overholser, J.C.; Jurjus, G.J.; Dieter, L.; Duman, R.S. Altered expression of synapse and glutamate related genes in post-mortem hippocampus of depressed subjects. Int. J. Neuropsychopharmacol. 2013, 16, 69–82. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.J.; Liang, S.; Spadoni, A.D.; Kelsoe, J.R.; Simmons, A.N. Whole brain expression of bipolar disorder associated genes: structural and genetic analyses. PLoS ONE 2014, 9, e100204. [Google Scholar] [CrossRef] [Green Version]
- Kalashnikova, E.; Lorca, R.A.; Kaur, I.; Barisone, G.A.; Li, B.; Ishimaru, T.; Trimmer, J.S.; Mohapatra, D.P.; Diaz, E. SynDIG1: An activity-regulated, AMPA- receptor-interacting transmembrane prote in that regulates excitatory synapse development. Neuron 2010, 65, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Dunn, E.C.; Sofer, T.; Wang, M.J.; Soare, T.W.; Gallo, L.C.; Gogarten, S.M.; Kerr, K.F.; Chen, C.Y.; Stein, M.B.; Ursano, R.J.; et al. Major Depressive Disorder Working Group of the Psychiatric Genomics, C.; Wassertheil-Smoller, S.; Smoller, J.W., Genome-wide association study of depressive symptoms in the Hispanic Community Health Study/Study of Latinos. J. Psychiatr. Res. 2018, 99, 167–176. [Google Scholar] [CrossRef]
- Kuhn, H.G.; Dickinson-Anson, H.; Gage, F.H. Neurogenesis in the dentate gyrus of the adult rat: Age-related decrease of neuronal progenitor proliferation. J. Neurosci. 1996, 16, 2027–2033. [Google Scholar] [CrossRef] [Green Version]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef]
- O’Reilly, K.C.; Shumake, J.; Gonzalez-Lima, F.; Lane, M.A.; Bailey, S.J. Chronic Administration of 13-Cis-Retinoic Acid Increases Depression-Related Behavior in Mice. Neuropsychopharmacology 2006, 31, 1919–1927. [Google Scholar] [CrossRef] [Green Version]
- Pizzagalli, D.A. Depression, stress, and anhedonia: Toward a synthesis and integrated model. Annu. Rev. Clin. Psychol. 2014, 10, 393–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, S.A.; Butler, P.; Munafo, M.R.; Nutt, D.J.; Robinson, E.S. A translational rodent assay of affective biases in depression and antidepressant therapy. Neuropsychopharmacology 2013, 38, 1625–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, S.A.; Wood, C.M.; Robinson, E.S.J. Using the affective bias test to predict drug-induced negative affect: Implications for drug safety. Br. J. Pharmacol. 2017, 174, 3200–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, S.A.; Cisneros, F.J.; Gough, B.; Hanig, J.P.; Berry, K.J. Chronic oral treatment with 13-cis-retinoic acid (isotretinoin) or all-trans-retinoic acid does not alter depression-like behaviors in rats. Toxicol. Sci. 2005, 87, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Crandall, J.; Sakai, Y.; Zhang, J.; Koul, O.; Mineur, Y.; Crusio, W.E.; McCaffery, P. 13-cis-retinoic acid suppresses hippocampal cell division and hippocampal-dependent learning in mice. Proc. Natl. Acad. Sci. USA 2004, 101, 5111–5116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Li, R.; Zhou, J.N. Chronic all-trans retinoic acid administration induces CRF over-expression accompanied by AVP up-regulation and multiple CRF-controlling receptors disturbance in the hypothalamus of rats. Brain Res. 2015, 1601, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Liu, J.; Zhao, J.; Qi, X.R.; Qi, C.C.; Lucassen, P.J.; Zhou, J.N. All-trans retinoic acid-induced hypothalamus-pituitary-adrenal hyperactivity involves glucocorticoid receptor dysregulation. Transl. Psychiatry 2013, 3, e336. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.A.; Cisneros, F.J.; Hanig, J.P.; Berry, K.J. Oral treatment with ACCUTANE does not increase measures of anhedonia or depression in rats. Neurotoxicol. Teratol. 2007, 29, 642–651. [Google Scholar] [CrossRef]
- Trent, S.; Drew, C.J.; Mitchell, P.J.; Bailey, S.J. Chronic treatment with 13-cis-retinoic acid changes aggressive behaviours in the resident-intruder paradigm in rats. Eur. Neuropsychopharmacol. 2009, 19, 876–886. [Google Scholar] [CrossRef] [Green Version]
- Balmer, J.E.; Blomhoff, R. Gene expression regulation by retinoic acid. J. Lipid Res. 2002, 43, 1773–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, S.M.; Kallarackal, A.J.; Kvarta, M.D.; Van Dyke, A.M.; LeGates, T.A.; Cai, X. An excitatory synapse hypothesis of depression. Trends Neurosci. 2015, 38, 279–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovero, K.L.; Blankenship, S.M.; Shi, Y.; Nicoll, R.A. SynDIG1 promotes excitatory synaptogenesis independent of AMPA receptor trafficking and biophysical regulation. PLoS ONE 2013, 8, e66171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Galan, M.; De Bundel, D.; Van Eeckhaut, A.; Smolders, I.; Lindskog, M. Dysfunctional astrocytic regulation of glutamate transmission in a rat model of depression. Mol. Psychiatry 2013, 18, 582–594. [Google Scholar] [CrossRef]
- Soprano, D.R.; Qin, P.; Soprano, K.J. Retinoic acid receptors and cancers. Annu. Rev. Nutr. 2004, 24, 201–221. [Google Scholar] [CrossRef]
- Bonhomme, D.; Pallet, V.; Dominguez, G.; Servant, L.; Henkous, N.; Lafenetre, P.; Higueret, P.; Beracochea, D.; Touyarot, K. Retinoic acid modulates intrahippocampal levels of corticosterone in middle-aged mice: Consequences on hippocampal plasticity and contextual memory. Front. Aging Neurosci. 2014, 6, 6. [Google Scholar] [CrossRef]
- Dennert, G.; Lotan, R. Effects of retinoic acid on the immune system: Stimulation of T killer cell induction. Eur. J. Immunol. 1978, 8, 23–29. [Google Scholar] [CrossRef]
- Qi, C.C.; Zhang, Z.; Fang, H.; Liu, J.; Zhou, N.; Ge, J.F.; Chen, F.H.; Xiang, C.B.; Zhou, J.N. Antidepressant effects of abscisic acid mediated by the downregulation of corticotrophin-releasing hormone gene expression in rats. Int. J. Neuropsychopharmacol. 2014, 18. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.T.; Ni, R.J.; Zhang, Z.; Zhao, J.; Liu, Y.J.; Zhou, J.N. Inhibition of oestrogen biosynthesis induces mild anxiety in C57BL/6J ovariectomized female mice. Neurosci. Bull 2011, 27, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.H.; Lo, K.L.; Lee, Y.H.; Hung, T.H.; Lin, Y.C. Berberine produces antidepressant-like effects in the forced swim test and in the tail suspension test in mice. Life Sci. 2007, 81, 933–938. [Google Scholar] [CrossRef]
- Strekalova, T.; Spanagel, R.; Bartsch, D.; Henn, F.A.; Gass, P. Stress-induced anhedonia in mice is associated with deficits in forced swimming and exploration. Neuropsychopharmacology 2004, 29, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Kamphuis, W.; Huitinga, I.; Zhou, J.N.; Swaab, D.F. Gene expression analysis in the human hypothalamus in depression by laser microdissection and real-time PCR: The presence of multiple receptor imbalances. Mol. Psychiatry 2008, 13, 786–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, X.-Y.; Fang, H.; Shan, Q.-H.; Qi, C.-C.; Zhou, J.-N. All-trans Retinoic Acid-induced Abnormal Hippocampal Expression of Synaptic Genes SynDIG1 and DLG2 is Correlated with Anxiety or Depression-Like Behavior in Mice. Int. J. Mol. Sci. 2020, 21, 2677. https://doi.org/10.3390/ijms21082677
Qin X-Y, Fang H, Shan Q-H, Qi C-C, Zhou J-N. All-trans Retinoic Acid-induced Abnormal Hippocampal Expression of Synaptic Genes SynDIG1 and DLG2 is Correlated with Anxiety or Depression-Like Behavior in Mice. International Journal of Molecular Sciences. 2020; 21(8):2677. https://doi.org/10.3390/ijms21082677
Chicago/Turabian StyleQin, Xin-Ya, Hui Fang, Qing-Hong Shan, Cong-Cong Qi, and Jiang-Ning Zhou. 2020. "All-trans Retinoic Acid-induced Abnormal Hippocampal Expression of Synaptic Genes SynDIG1 and DLG2 is Correlated with Anxiety or Depression-Like Behavior in Mice" International Journal of Molecular Sciences 21, no. 8: 2677. https://doi.org/10.3390/ijms21082677
APA StyleQin, X. -Y., Fang, H., Shan, Q. -H., Qi, C. -C., & Zhou, J. -N. (2020). All-trans Retinoic Acid-induced Abnormal Hippocampal Expression of Synaptic Genes SynDIG1 and DLG2 is Correlated with Anxiety or Depression-Like Behavior in Mice. International Journal of Molecular Sciences, 21(8), 2677. https://doi.org/10.3390/ijms21082677