GABAergic Transmission in the Basolateral Amygdala Differentially Modulates Plasticity in the Dentate Gyrus and the CA1 Areas


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
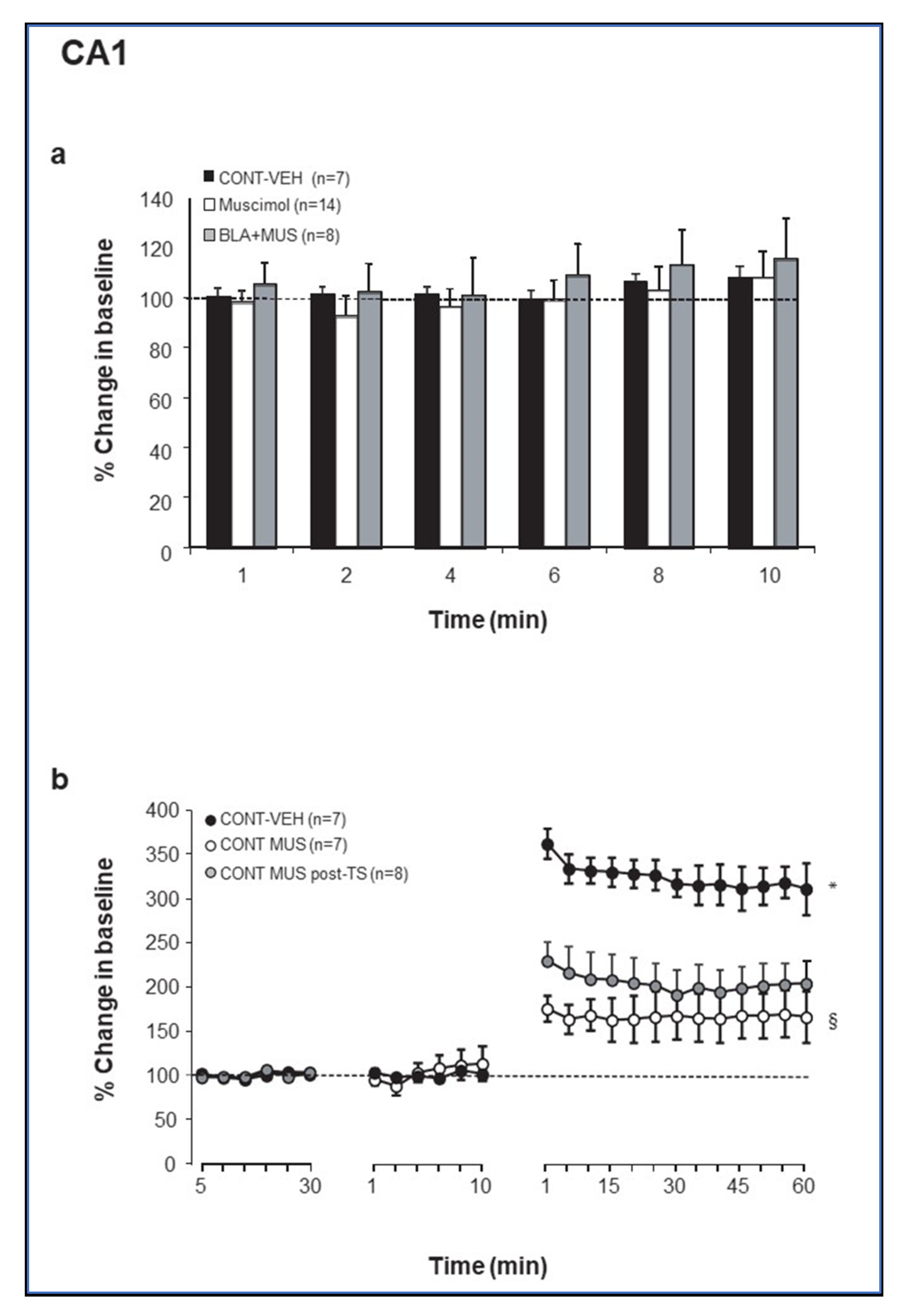
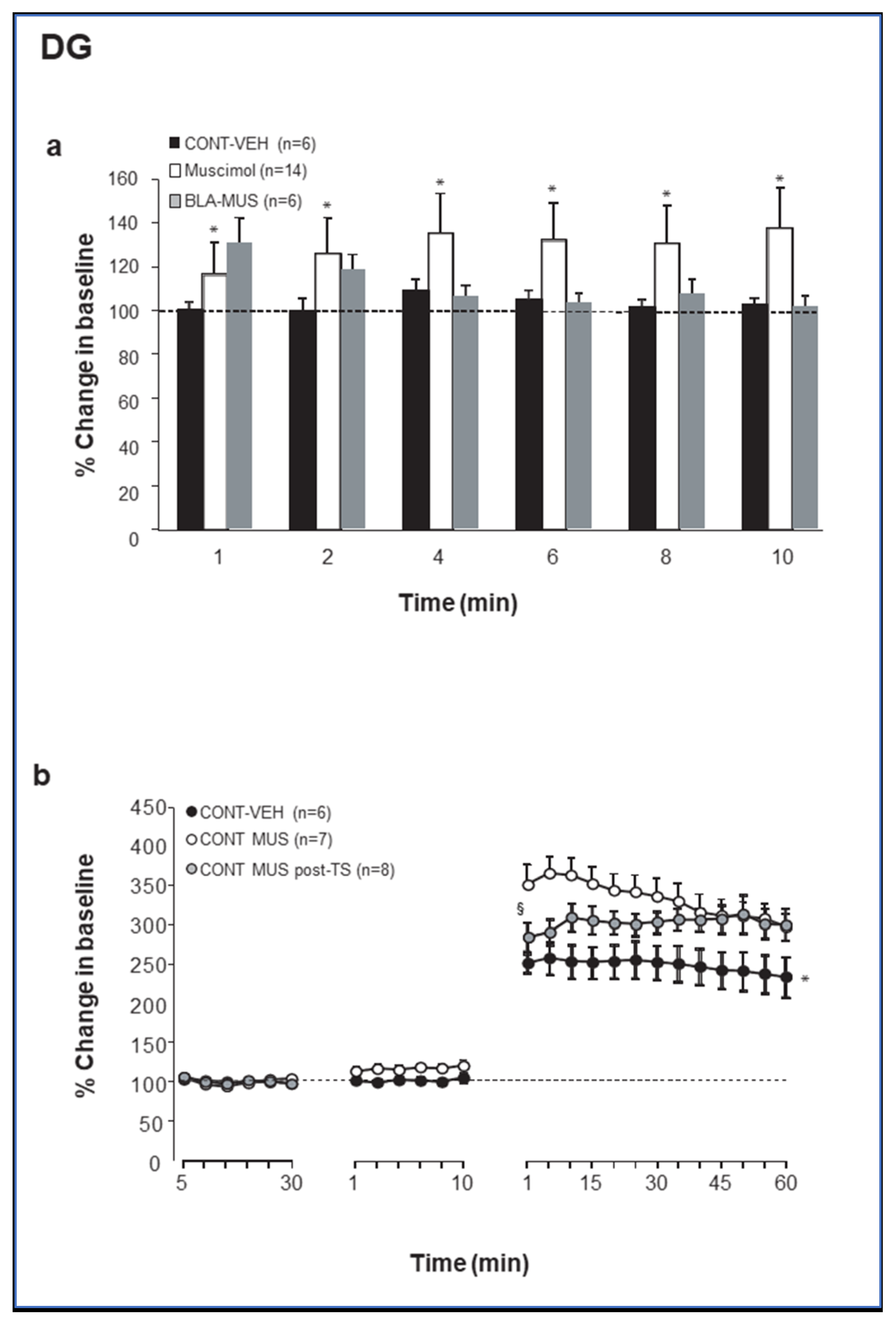
2.1. Muscimol Microinfusion into the BLA Increases Excitationin the PP-DG Pathway but Not in the CA1
2.2. Theta Stimulation Enhances Excitation in the DG but Not CA1 Following Increased GABAergic Transmission in the BLA
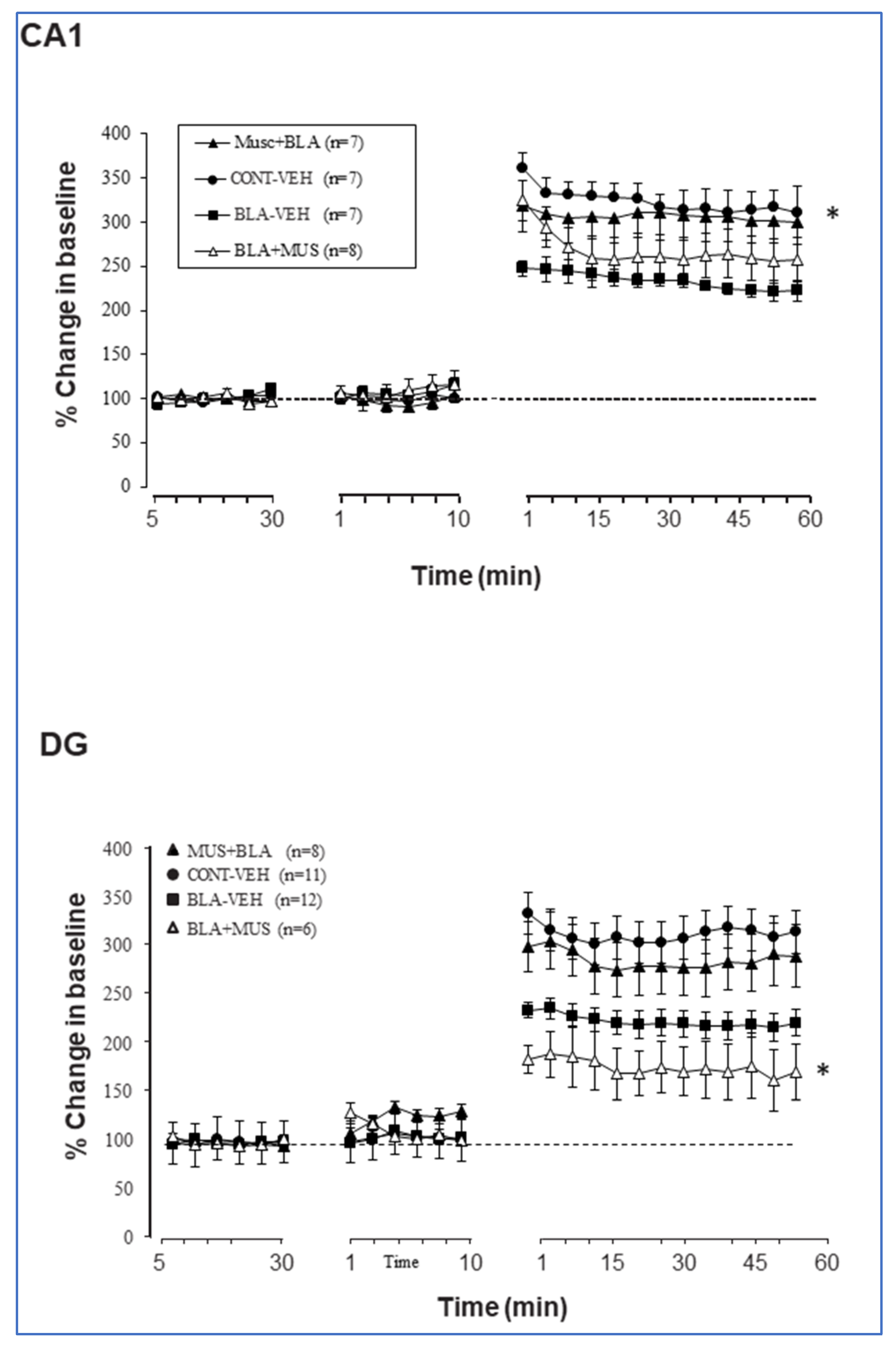
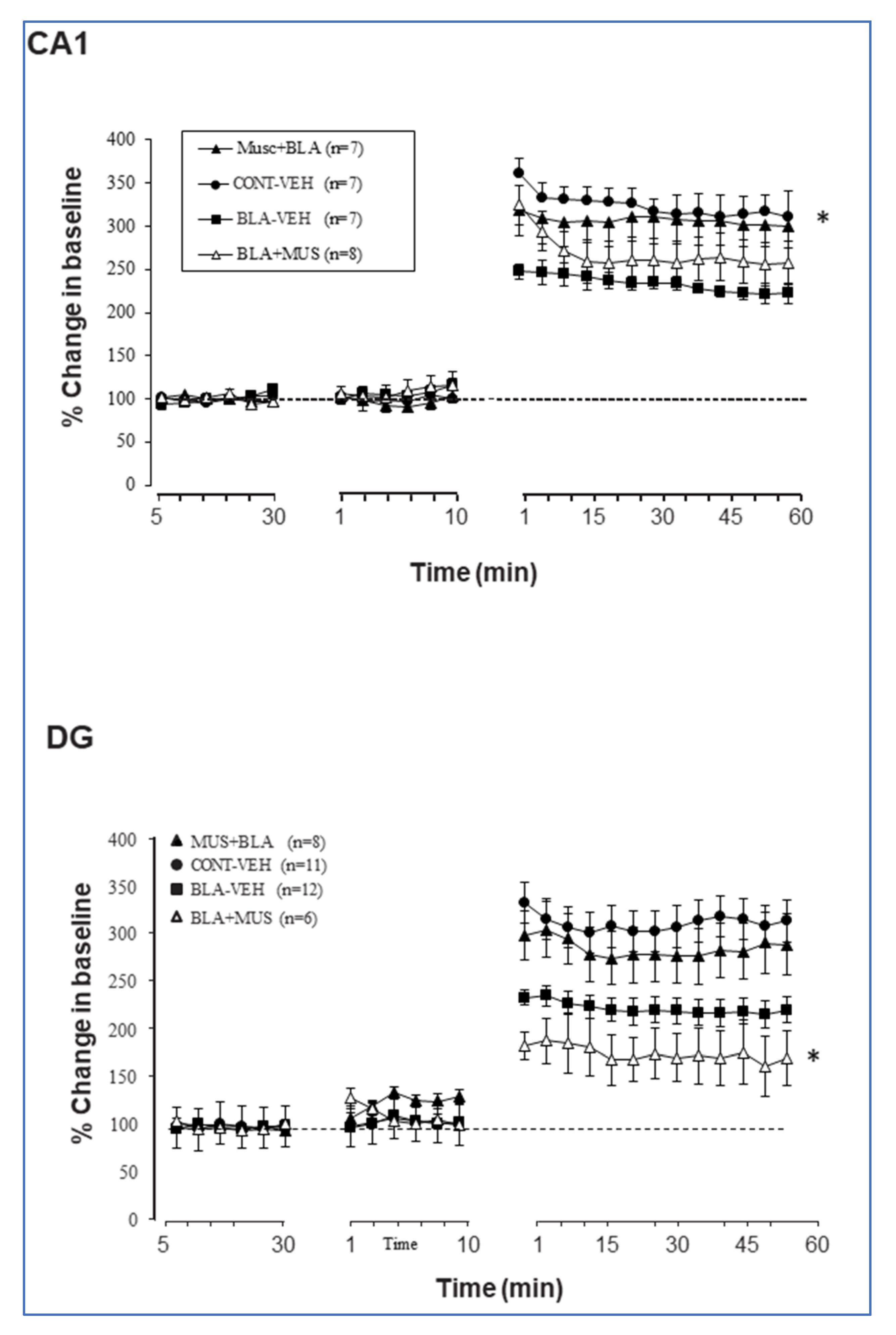
2.3. Muscimol Application to the Amygdala Increases DG Excitation, Which Is Attenuated if BLA Is Primed
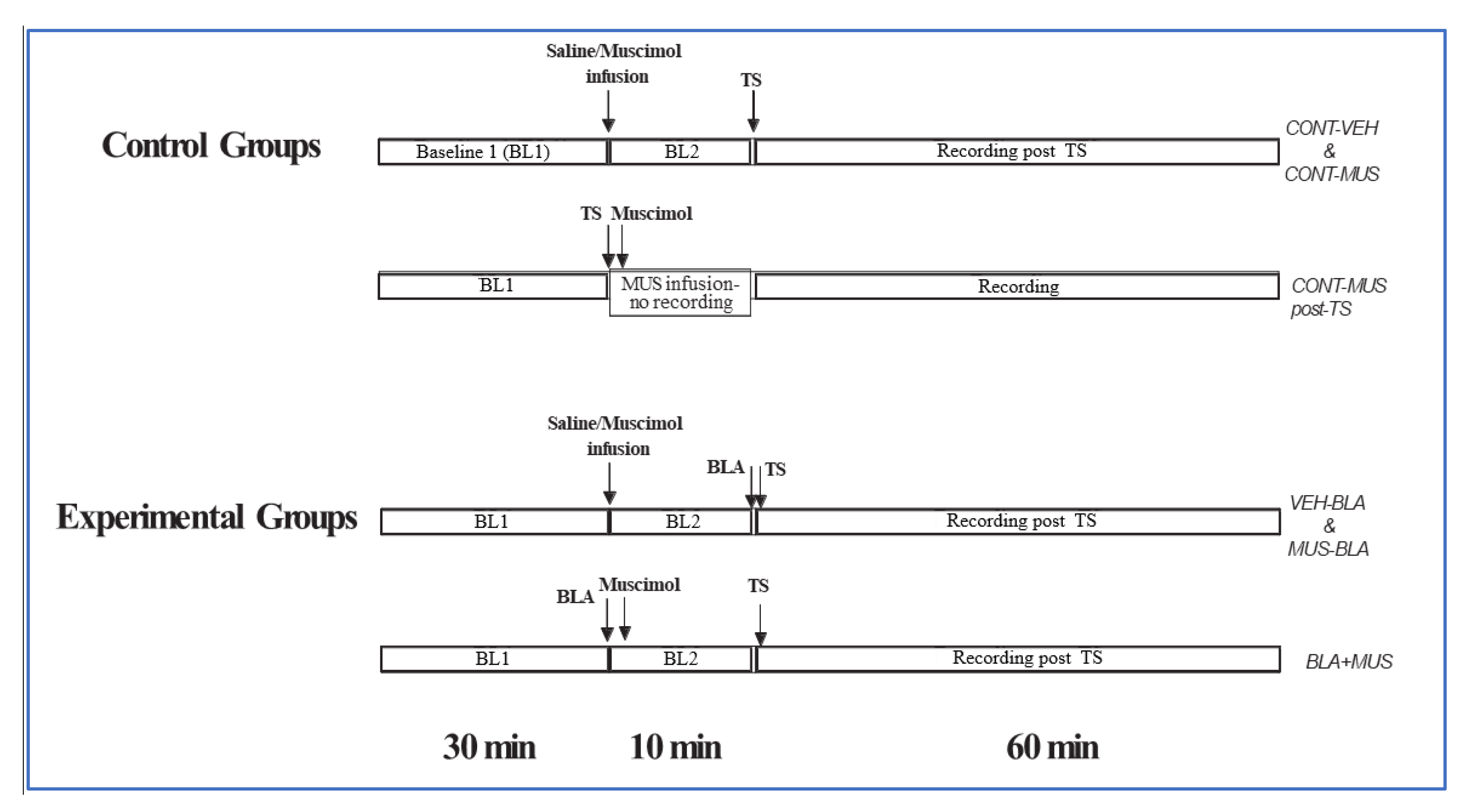
3. Materials and Methods
3.1. Subjects
3.2. Surgery
3.3. Evoked Field Potentials in the CA1 and DG
3.4. Amygdala Activation and Muscimol/Vehicle Injection
3.5. Stimulating and Recording Procedures
3.6. Drugs and Infusion
3.7. Protocols
3.7.1. Muscimol
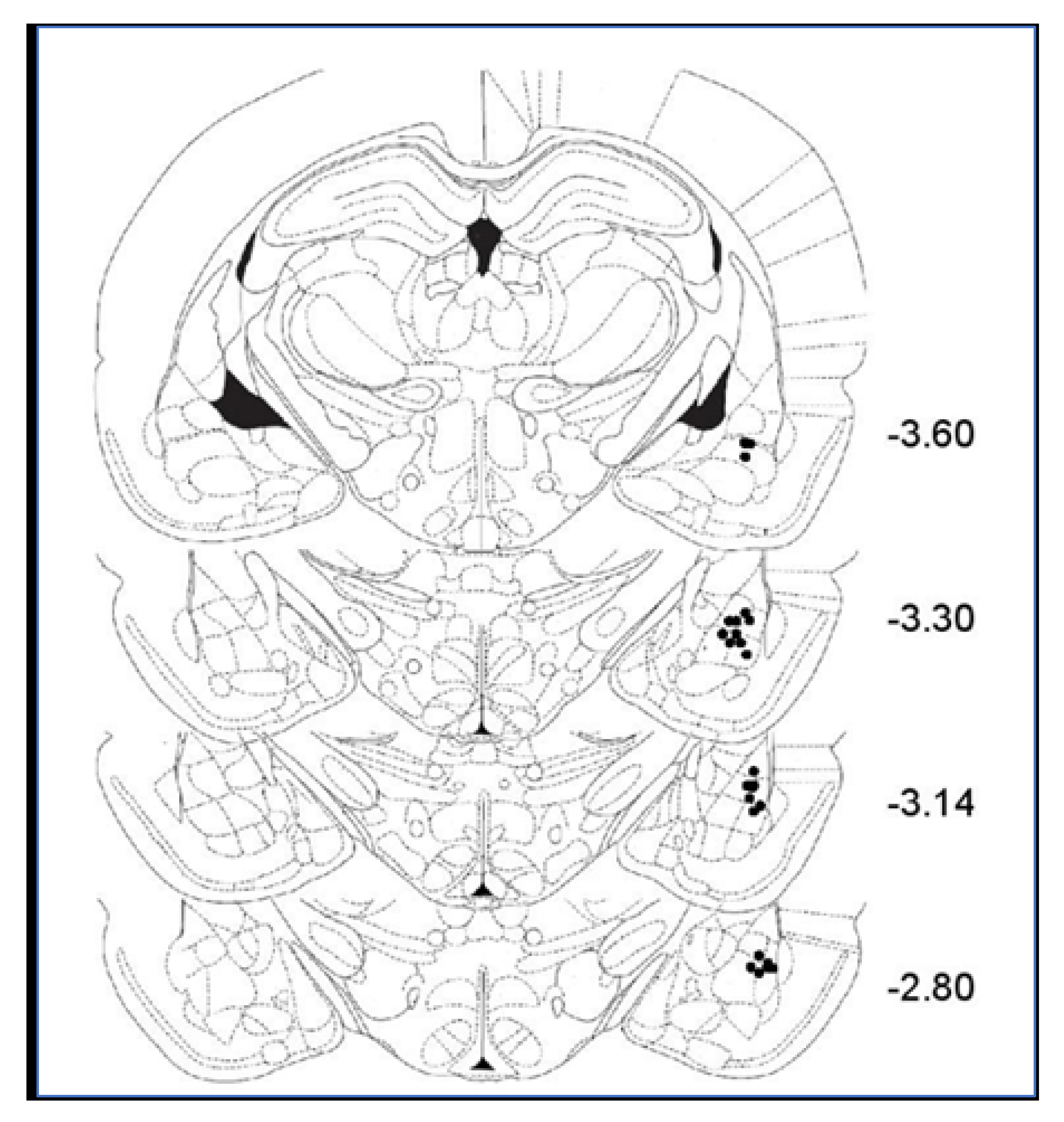
3.7.2. Histology
3.8. Data Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Abraham, W.C.; Bear, M.F. Metaplasticity: The plasticity of synaptic plasticity. Trends Neurosci. 1996, 19, 126–130. [Google Scholar] [CrossRef]
- Abraham, W.C.; Richter-Levin, G. From Synaptic Metaplasticity to Behavioral Metaplasticity. Neurobiol. Learn. Mem. 2018, 154, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.V.; Abraham, W.C.; Maroun, M.; Stork, O.; Richter-Levin, G. Stress-induced metaplasticity: From synapses to behavior. Neuroscience 2013, 250, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Richter-Levin, G.; Maroun, M. Stress and amygdala suppression of metaplasticity in the medial prefrontal cortex. Cereb. Cortex 2010, 20, 2433–2441. [Google Scholar] [CrossRef] [Green Version]
- Foy, M.R.; Stanton, M.E.; Levine, S.; Thompson, R.F. Behavioral stress impairs long-term potentiation in rodent hippocampus. Behav. Neural Biol. 1987, 48, 138–149. [Google Scholar] [CrossRef]
- Shors, T.J.; Seib, T.B.; Levine, S.; Thompson, R.F. Inescapable versus escapable shock modulates long-term potentiation in the rat hippocampus. Science 1989, 244, 224–226. [Google Scholar] [CrossRef]
- Diamond, D.M.; Bennett, M.C.; Fleshner, M.; Rose, G.M. Inverted-U relationship between the level of peripheral corticosterone and the magnitude of hippocampal primed burst potentiation. Hippocampus 1992, 2, 421–430. [Google Scholar] [CrossRef]
- Pavlides, C.; Ogawa, S.; Kimura, A.; McEwen, B.S. Role of adrenal steroid mineralocorticoid and glucocorticoid receptors in long-term potentiation in the CA1 field of hippocampal slices. Brain Res. 1996, 738, 229–235. [Google Scholar] [CrossRef]
- Pavlides, C.; Watanabe, Y.; McEwen, B.S. Effects of glucocorticoids on hippocampal long-term potentiation. Hippocampus 1993, 3, 183–192. [Google Scholar] [CrossRef]
- Shors, T.J.; Dryver, E. Effects of stress and long-term potentiation (LTP) on subsequent LTP and the theta burst response in the dentate gyrus. Brain Res. 1994, 666, 232–238. [Google Scholar] [CrossRef]
- Bramham, C.R.; Southard, T.; Ahlers, S.T.; Sarvey, J.M. Acute cold stress leading to elevated corticosterone neither enhances synaptic efficacy nor impairs LTP in the dentate gyrus of freely moving rats. Brain Res. 1998, 789, 245–255. [Google Scholar] [CrossRef]
- Kim, J.J.; Diamond, D.M. The stressed hippocampus, synaptic plasticity and lost memories. Nat. Rev. 2002, 3, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Kavushansky, A.; Richter-Levin, G. Effects of stress and corticosterone on activity and plasticity in the amygdala. J. Neurosci. Res. 2006, 84, 1580–1587. [Google Scholar] [CrossRef]
- Kavushansky, A.; Vouimba, R.-M.; Cohen, H.; Richter-Levin, G. Activity and plasticity in the CA1, the dentate gyrus, and the amygdala following controllable vs. uncontrollable water stress. Hippocampus 2006, 16, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Roozendaal, B.; McReynolds, J.R.; Van der Zee, E.A.; Lee, S.; McGaugh, J.L.; McIntyre, C.K. Glucocorticoid effects on memory consolidation depend on functional interactions between the medial prefrontal cortex and basolateral amygdala. J. Neurosci. 2009, 29, 14299–14308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez, C.M.; McGaugh, J.L.; Weinberger, N.M. Activation of the basolateral amygdala induces long-term enhancement of specific memory representations in the cerebral cortex. Neurobiol. Learn. Mem. 2013, 101, 8–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegaya, Y.; Saito, H.; Torii, K.; Nishiyama, N. Activin selectively abolishes hippocampal long-term potentiation induced by weak tetanic stimulation in vivo. Jpn. J. Pharmacol. 1997, 75, 87–89. [Google Scholar] [CrossRef] [Green Version]
- Akirav, I.; Richter-Levin, G. Biphasic modulation of hippocampal plasticity by behavioral stress and basolateral amygdala stimulation in the rat. J. Neurosci. 1999, 19, 10530–10535. [Google Scholar] [CrossRef] [Green Version]
- Nakao, K.; Matsuyama, K.; Matsuki, N.; Ikegaya, Y. Amygdala stimulation modulates hippocampal synaptic plasticity. Proc. Natl. Acad. Sci. USA 2004, 101, 14270–14275. [Google Scholar] [CrossRef] [Green Version]
- Bergado, J.A.; Scherf, T.; Almaguer-Melian, W.; Frey, S.; Lopez, J.; Frey, J.U. Stimulation of the nucleus raphe medialis modifies basal synaptic transmission at the dentate gyrus, but not long-term potentiation or its reinforcement by stimulation of the basolateral amygdala. Neurosci. Lett. 2009, 464, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Vouimba, R.M.; Richter-Levin, G. Different patterns of amygdala priming differentially affect dentate gyrus plasticity and corticosterone, but not CA1 plasticity. Front. Neural Circuits 2013, 7, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vouimba, R.M.; Richter-Levin, G. Physiological dissociation in hippocampal subregions in response to amygdala stimulation. Cereb. Cortex 2005, 15, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.; Rainnie, D.; Cassell, M. Neurotransmission in the rat amygdala related to fear and anxiety. Trends Neurosci. 1994, 17, 208–214. [Google Scholar] [CrossRef]
- Kilpatrick, L.; Cahill, L. Amygdala modulation of parahippocampal and frontal regions during emotionally influenced memory storage. Neuroimage 2003, 20, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Ferry, B.; Parrot, S.; Marien, M.; Lazarus, C.; Cassel, J.C.; McGaugh, J.L. Noradrenergic influences in the basolateral amygdala on inhibitory avoidance memory are mediated by an action on alpha2-adrenoceptors. Psychoneuroendocrinology 2015, 51, 68–79. [Google Scholar] [PubMed]
- Marek, R.; Jin, J.; Goode, T.D.; Giustino, T.F.; Wang, Q.; Acca, G.M.; Holehonnur, R.; Ploski, J.E.; Fitzgerald, P.J.; Lynagh, T.; et al. Hippocampus-driven feed-forward inhibition of the prefrontal cortex mediates relapse of extinguished fear. Nat. Neurosci. 2018, 21, 384–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vouimba, R.M.; Yaniv, D.; Richter-Levin, G. Glucocorticoid receptors and beta-adrenoceptors in basolateral amygdala modulate synaptic plasticity in hippocampal dentate gyrus, but not in area CA1. Neuropharmacology 2007, 52, 244–252. [Google Scholar] [CrossRef]
- Takagi, M.; Yamamoto, C. The long-lasting inhibition recorded in vitro from the lateral nucleus of the amygdala. Brain Res. 1981, 206, 474–478. [Google Scholar] [CrossRef]
- Washburn, M.S.; Moises, H.C. Inhibitory responses of rat basolateral amygdaloid neurons recorded in vitro. Neuroscience 1992, 50, 811–830. [Google Scholar] [CrossRef] [Green Version]
- Niehoff, D.L.; Kuhar, M.J. Benzodiazepine receptors: Localization in rat amygdala. J. Neurosci. 1983, 3, 2091–2097. [Google Scholar] [CrossRef] [Green Version]
- Lo, M.M.; Niehoff, D.L.; Kuhar, M.J.; Snyder, S.H. Differential localization of type I and type II benzodiazepine binding sites in substantia nigra. Nature 1983, 306, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Salinas, J.A.; McGaugh, J.L. Muscimol induces retrograde amnesia for changes in reward magnitude. Neurobiol. Learn. Mem. 1995, 63, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, S.K.; Shekhar, A. Regulation of anxiety by GABAA receptors in the rat amygdala. Pharmacol. Biochem. Behav. 1995, 52, 701–706. [Google Scholar] [CrossRef]
- McDonald, A.J.; Mascagni, F.; Muller, J.F. Immunocytochemical localization of GABABR1 receptor subunits in the basolateral amygdala. Brain Res. 2004, 1018, 147–158. [Google Scholar] [CrossRef]
- Dubrovina, N.I. GABA-Receptors in Modulation of Fear Memory Extinction. Zh. Vyssh. Nerv. Deiat. Im. I P Pavlova 2016, 66, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Squires, R.F.; Saederup, E. A review of evidence for GABergic predominance/glutamatergic deficit as a common etiological factor in both schizophrenia and affective psychoses: More support for a continuum hypothesis of “functional” psychosis. Neurochem. Res. 1991, 16, 1099–1111. [Google Scholar] [CrossRef]
- Pham, X.; Sun, C.; Chen, X.; van den Oord, E.J.; Neale, M.C.; Kendler, K.S.; Hettema, J.M. Association study between GABA receptor genes and anxiety spectrum disorders. Depress. Anxiety 2009, 26, 998–1003. [Google Scholar] [CrossRef] [Green Version]
- Akirav, I.; Raizel, H.; Maroun, M. Enhancement of conditioned fear extinction by infusion of the GABA(A) agonist muscimol into the rat prefrontal cortex and amygdala. Eur. J. Neurosci. 2006, 23, 758–764. [Google Scholar] [CrossRef]
- Akirav, I.; Richter-Levin, G. Factors that determine the non-linear amygdala influence on hippocampus-dependent memory. Dose. Response. 2006, 4, 22–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaniv, D.; Vouimba, R.M.; Diamond, D.M.; Richter-Levin, G. Simultaneous induction of long-term potentiation in the hippocampus and the amygdala by entorhinal cortex activation: Mechanistic and temporal profiles. Neuroscience 2003, 120, 1125–1135. [Google Scholar] [CrossRef]
- Martin, J.H. Autoradiographic estimation of the extent of reversible inactivation produced by microinjection of lidocaine and muscimol in the rat. Neurosci. Lett. 1991, 127, 160–164. [Google Scholar] [CrossRef]
- Krupa, D.J.; Weng, J.; Thompson, R.F. Inactivation of brainstem motor nuclei blocks expression but not acquisition of the rabbit’s classically conditioned eyeblink response. Behav. Neurosci. 1996, 110, 219–227. [Google Scholar] [CrossRef]
- Akirav, I.; Richter-Levin, G. Mechanisms of amygdala modulation of hippocampal plasticity. J. Neurosci. 2002, 22, 9912–9921. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Richter-Levin, G. Priming stimulation of basal but not lateral amygdala affects long-term potentiation in the rat dentate gyrus in vivo. Neuroscience 2013, 246, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Saha, R.; Shrivastava, K.; Jing, L.; Schayek, R.; Maroun, M.; Kriebel, M.; Volkmer, H.; Richter-Levin, G. Perturbation of GABAergic Synapses at the Axon Initial Segment of Basolateral Amygdala Induces Trans-regional Metaplasticity at the Medial Prefrontal Cortex. Cereb. Cortex 2018, 28, 395–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pape, H.C.; Pare, D. Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear. Physiol. Rev. 2010, 90, 419–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Amir, A.; Haufler, D.; Pare, D. Differential Recruitment of Competing Valence-Related Amygdala Networks during Anxiety. Neuron 2017, 96, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Sah, P. Fear, Anxiety, and the Amygdala. Neuron 2017, 96, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Pare, D.; Royer, S.; Smith, Y.; Lang, E.J. Contextual inhibitory gating of impulse traffic in the intra-amygdaloid network. Ann. N. Y. Acad. Sci. 2003, 985, 78–91. [Google Scholar] [CrossRef]
- Wanasuntronwong, A.; Wanakhachornkrai, O.; Phongphanphanee, P.; Isa, T.; Tantisira, B.; Tantisira, M.H. Modulation of Neuronal Activity on Intercalated Neurons of Amygdala Might Underlie Anxiolytic Activity of a Standardized Extract of Centella asiatica ECa233. Evid. Based. Complement. Altern. Med. 2018, 2018, 3853147. [Google Scholar] [CrossRef] [Green Version]
- Marowsky, A.; Vogt, K.E. Delta-subunit-containing GABAA-receptors mediate tonic inhibition in paracapsular cells of the mouse amygdala. Front. Neural Circuits 2014, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fino, E.; Vandecasteele, M.; Perez, S.; Saudou, F.; Venance, L. Region-specific and state-dependent action of striatal GABAergic interneurons. Nat. Commun. 2018, 9, 3335–3339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahanty, N.K.; Sah, P. Calcium-permeable AMPA receptors mediate long-term potentiation in interneurons in the amygdala. Nature 1998, 394, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wilson, W.A.; Moore, S.D. Role of NMDA, non-NMDA, and GABA receptors in signal propagation in the amygdala formation. J. Neurophysiol. 2001, 86, 1422–1429. [Google Scholar] [CrossRef]
- Berlau, D.J.; McGaugh, J.L. Enhancement of extinction memory consolidation: The role of the noradrenergic and GABAergic systems within the basolateral amygdala. Neurobiol. Learn. Mem. 2006, 86, 123–132. [Google Scholar] [CrossRef]
- Makkar, S.R.; Zhang, S.Q.; Cranney, J. Behavioral and neural analysis of GABA in the acquisition, consolidation, reconsolidation, and extinction of fear memory. Neuropsychopharmacology 2010, 35, 1625–1652. [Google Scholar] [CrossRef]
- Guo, Y.; Tang, X.; Zhang, J.; Jin, S.; Li, J.; Ding, L.; Zhang, K.; Yang, C.; Zhou, H.; He, X.; et al. Corticosterone Signaling and a Lateral Habenula-Ventral Tegmental Area Circuit Modulate Compulsive Self-Injurious Behavior in a Rat Model. J. Neurosci. 2018, 38, 5251–5266. [Google Scholar] [CrossRef]
- Inoue, R.; Abdou, K.; Hayashi-Tanaka, A.; Muramatsu, S.I.; Mino, K.; Inokuchi, K.; Mori, H. Glucocorticoid receptor-mediated amygdalar metaplasticity underlies adaptive modulation of fear memory by stress. eLife 2018, 7, e34135. [Google Scholar] [CrossRef]
- Majewska, M.D.; Bisserbe, J.C.; Eskay, R.L. Glucocorticoids are modulators of GABAA receptors in brain. Brain Res. 1985, 339, 178–182. [Google Scholar] [CrossRef]
- Karst, H.; Berger, S.; Erdmann, G.; Schütz, G.; Joëls, M. Metaplasticity of amygdalar responses to the stress hormone corticosterone. Proc. Natl. Acad. Sci. USA 2010, 107, 14449–14454. [Google Scholar] [CrossRef] [Green Version]
- Karst, H.; Joëls, M. Severe stress hormone conditions cause an extended window of excitability in the mouse basolateral amygdala. Neuropharmacology 2016, 110, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Desgranges, B.; Levy, F.; Ferreira, G. Anisomycin infusion in amygdala impairs consolidation of odor aversion memory. Brain Res. 2008, 1236, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Desmedt, A.; Marighetto, A.; Richter-Levin, G.; Calandreau, L. Adaptive emotional memory: The key hippocampal-amygdalar interaction. Stress 2015, 18, 297–308. [Google Scholar] [CrossRef]
- Hadad-Ophir, O.; Albrecht, A.; Stork, O.; Richter-Levin, G. Amygdala activation and GABAergic gene expression in hippocampal sub-regions at the interplay of stress and spatial learning. Front. Behav. Neurosci. 2014, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Bender, C.L.; Otamendi, A.; Calfa, G.D.; Molina, V.A. Prior stress promotes the generalization of contextual fear memories: Involvement of the gabaergic signaling within the basolateral amygdala complex. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 83, 18–26. [Google Scholar] [CrossRef]
- Cossart, R.; Bernard, C.; Ben-Ari, Y. Multiple facets of GABAergic neurons and synapses: Multiple fates of GABA signalling in epilepsies. Trends Neurosci. 2005, 28, 108–115. [Google Scholar] [CrossRef]
- Marty, A.; Llano, I. Excitatory effects of GABA in established brain networks. Trends Neurosci. 2005, 28, 284–289. [Google Scholar] [CrossRef]
- Quirk, G.J.; Gehlert, D.R. Inhibition of the amygdala: Key to pathological states? Ann. N. Y. Acad. Sci. 2003, 985, 263–272. [Google Scholar] [CrossRef]
- Johnson, S.A.; Turner, S.M.; Lubke, K.N.; Cooper, T.L.; Fertal, K.E.; Bizon, J.L.; Maurer, A.P.; Burke, S.N. Experience-Dependent Effects of Muscimol-Induced Hippocampal Excitation on Mnemonic Discrimination. Front. Syst. Neurosci. 2019, 12, 72. [Google Scholar] [CrossRef]
- Steele, P.M.; Mauk, M.D. Inhibitory control of LTP and LTD: Stability of synapse strength. J. Neurophysiol. 1999, 81, 1559–1566. [Google Scholar] [CrossRef]
- Edeline, J.M.; Hars, B.; Hennevin, E.; Cotillon, N. Muscimol diffusion after intracerebral microinjections: A reevaluation based on electrophysiological and autoradiographic quantifications. Neurobiol. Learn. Mem. 2002, 78, 100–124. [Google Scholar] [CrossRef] [PubMed]
- Holahan, M.R.; White, N.M. Intra-amygdala muscimol injections impair freezing and place avoidance in aversive contextual conditioning. Learn. Mem. 2004, 11, 436–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.H.; Schmidt, M.; Hall, W.C. Excitatory and inhibitory circuitry in the superficial gray layer of the superior colliculus. J. Neurosci. 2001, 21, 8145–8153. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Monfils, M.H.; Ledoux, J.E. GABA(C) Receptors in the Lateral Amygdala: A Possible Novel Target for the Treatment of Fear and Anxiety Disorders? Front. Behav. Neurosci. 2010, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Xiao, Y.; Zhang, X.; Shu, Y. Activation of axon initial segmental GABAA receptors inhibits action potential generation in neocortical GABAergic interneurons. Neuropharmacology 2018, 138, 97–105. [Google Scholar] [CrossRef]
- Hariri, A.R.; Holmes, A. Finding translation in stress research. Nat. Neurosci. 2015, 18, 1347–1352. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Akirav, I.; Richter-Levin, G. Short-term behavioral and electrophysiological consequences of underwater trauma. Physiol. Behav. 2000, 70, 327–332. [Google Scholar] [CrossRef]
- Akirav, I.; Sandi, C.; Richter-Levin, G. Differential activation of hippocampus and amygdala following spatial learning under stress. Eur. J. Neurosci. 2001, 14, 719–725. [Google Scholar] [CrossRef]
- Vouimba, R.M.; Yaniv, D.; Diamond, D.; Richter-Levin, G. Effects of inescapable stress on LTP in the amygdala versus the dentate gyrus of freely behaving rats. Eur. J. Neurosci. 2004, 19, 1887–1894. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vouimba, R.-M.; Anunu, R.; Richter-Levin, G. GABAergic Transmission in the Basolateral Amygdala Differentially Modulates Plasticity in the Dentate Gyrus and the CA1 Areas. Int. J. Mol. Sci. 2020, 21, 3786. https://doi.org/10.3390/ijms21113786
Vouimba R-M, Anunu R, Richter-Levin G. GABAergic Transmission in the Basolateral Amygdala Differentially Modulates Plasticity in the Dentate Gyrus and the CA1 Areas. International Journal of Molecular Sciences. 2020; 21(11):3786. https://doi.org/10.3390/ijms21113786
Chicago/Turabian StyleVouimba, Rose-Marie, Rachel Anunu, and Gal Richter-Levin. 2020. "GABAergic Transmission in the Basolateral Amygdala Differentially Modulates Plasticity in the Dentate Gyrus and the CA1 Areas" International Journal of Molecular Sciences 21, no. 11: 3786. https://doi.org/10.3390/ijms21113786
APA StyleVouimba, R. -M., Anunu, R., & Richter-Levin, G. (2020). GABAergic Transmission in the Basolateral Amygdala Differentially Modulates Plasticity in the Dentate Gyrus and the CA1 Areas. International Journal of Molecular Sciences, 21(11), 3786. https://doi.org/10.3390/ijms21113786