Inhibition of Heparanase Expression Results in Suppression of Invasion, Migration and Adhesion Abilities of Bladder Cancer Cells


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
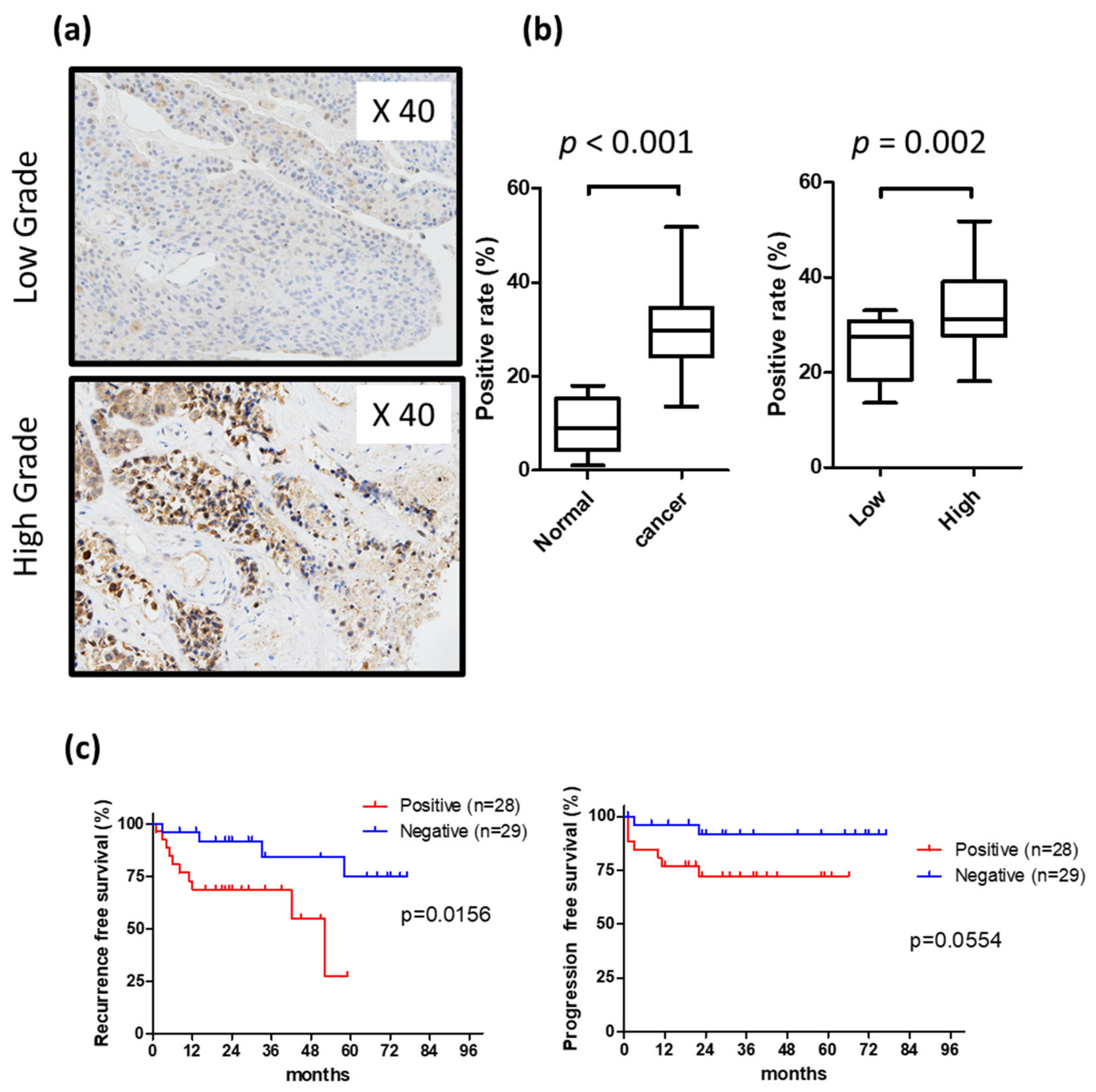
2.1. Heparanase Is Overexpressed in Human Urothelial Carcinoma of the Urinary Bladder, and Heparanase Expression Levels Are Associated with Intravesical Recurrence
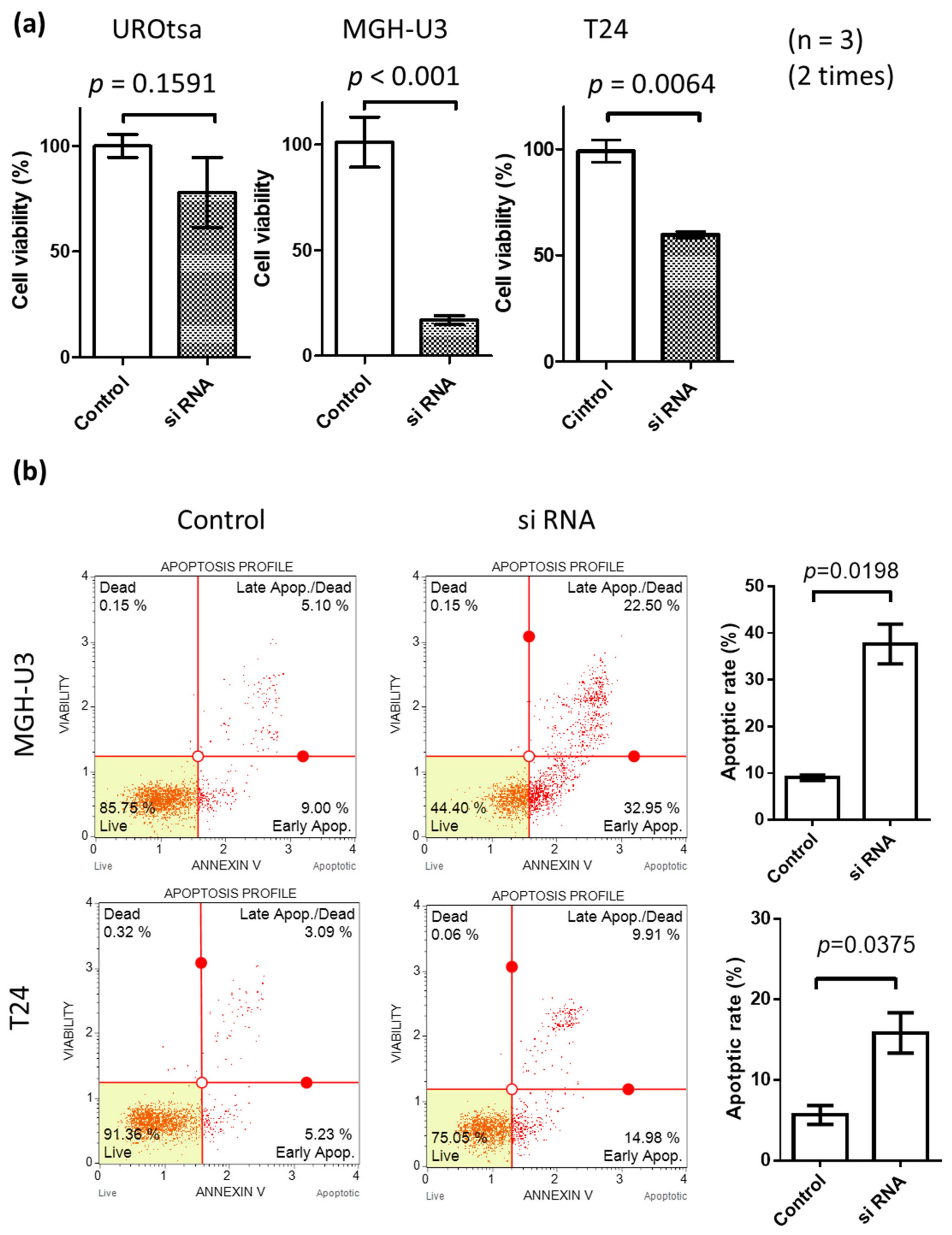
2.2. Knockdown of Heparanase-Induced Apoptosis in Urothelial Carcinoma Cells
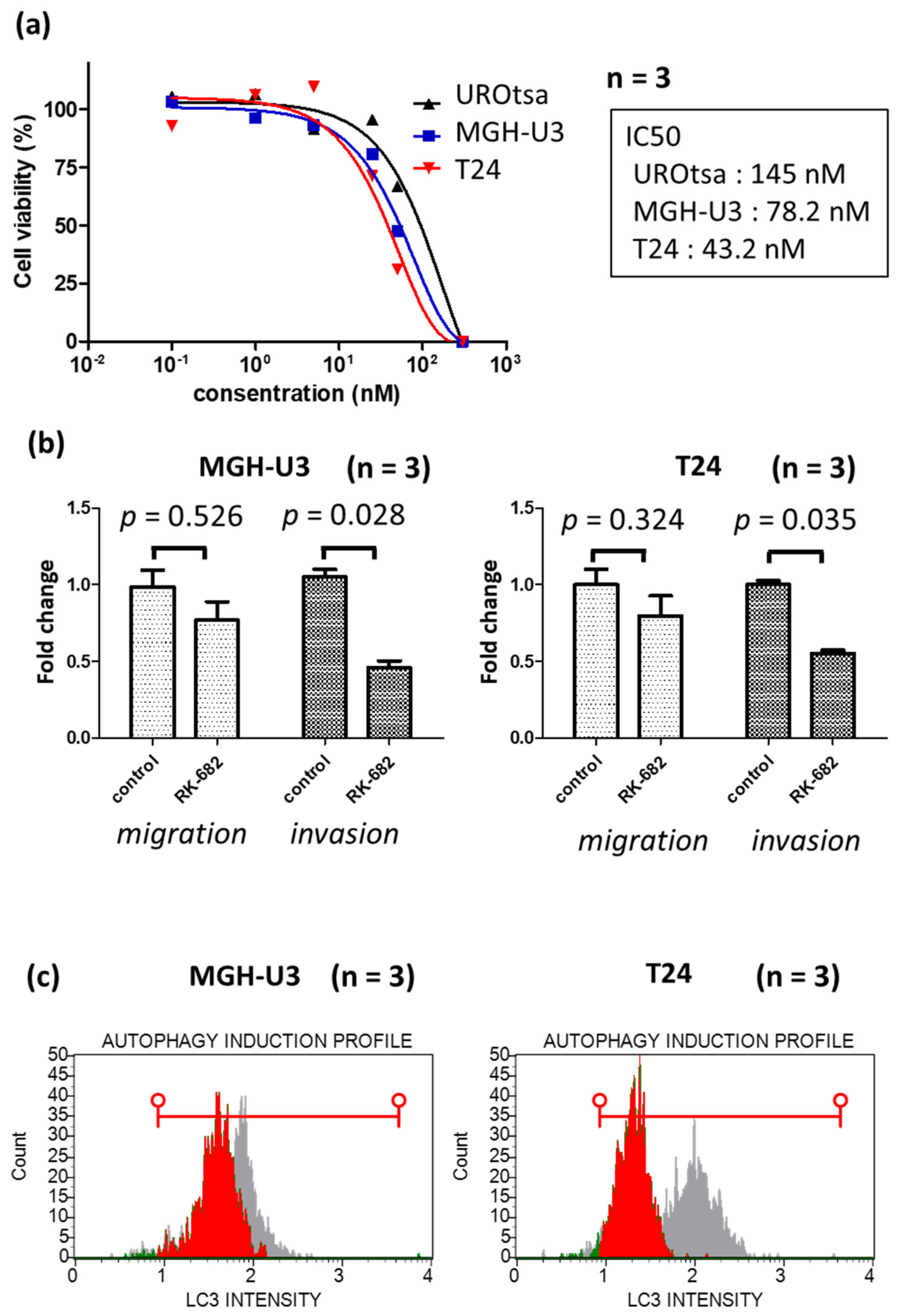
2.3. The Multi Enzyme Inhibitor RK-682, Which Is Also a Heparanase Inhibitor, Suppresses Cell Proliferation and Autophagy in Human Urothelial Cancer Cell Lines
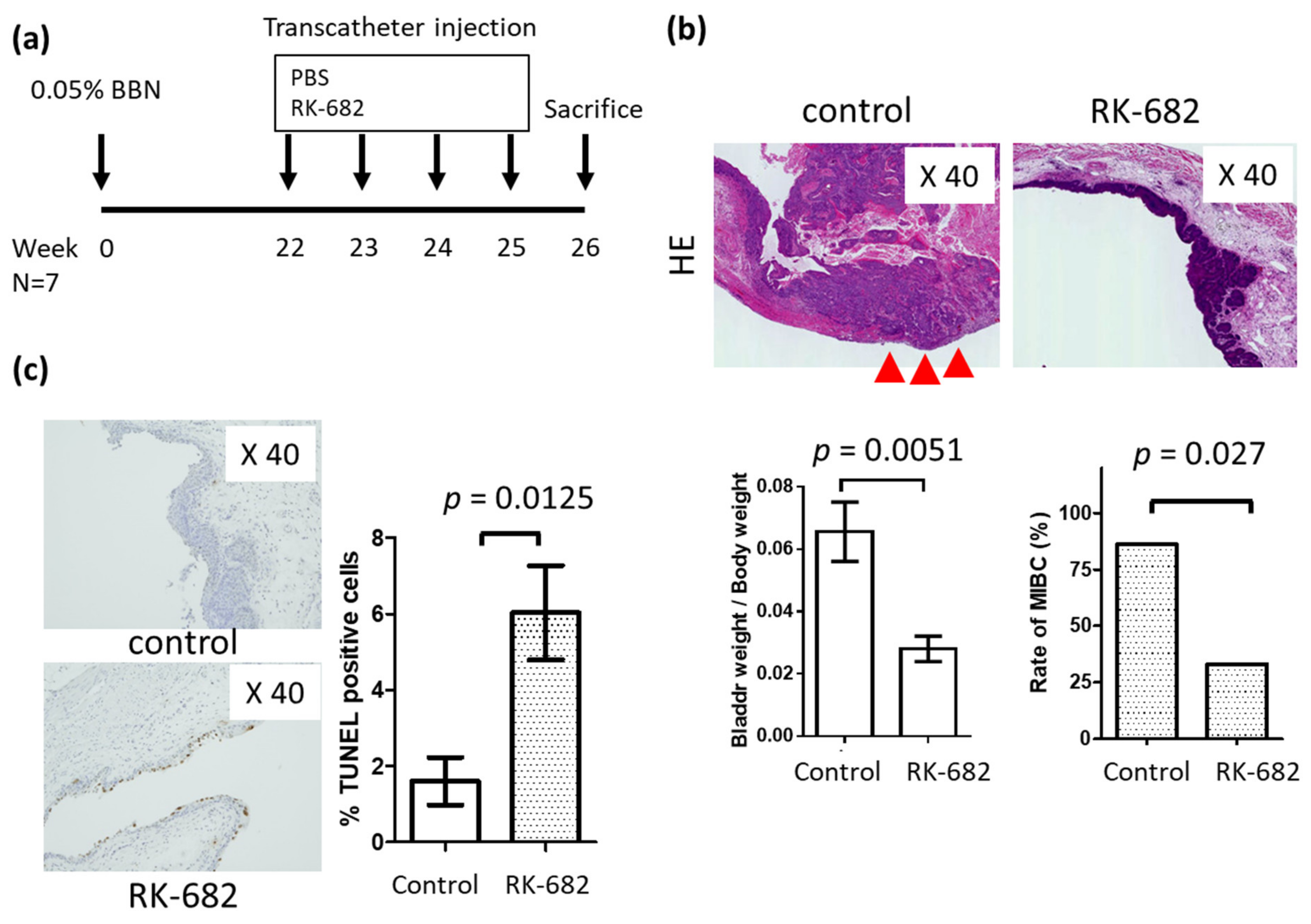
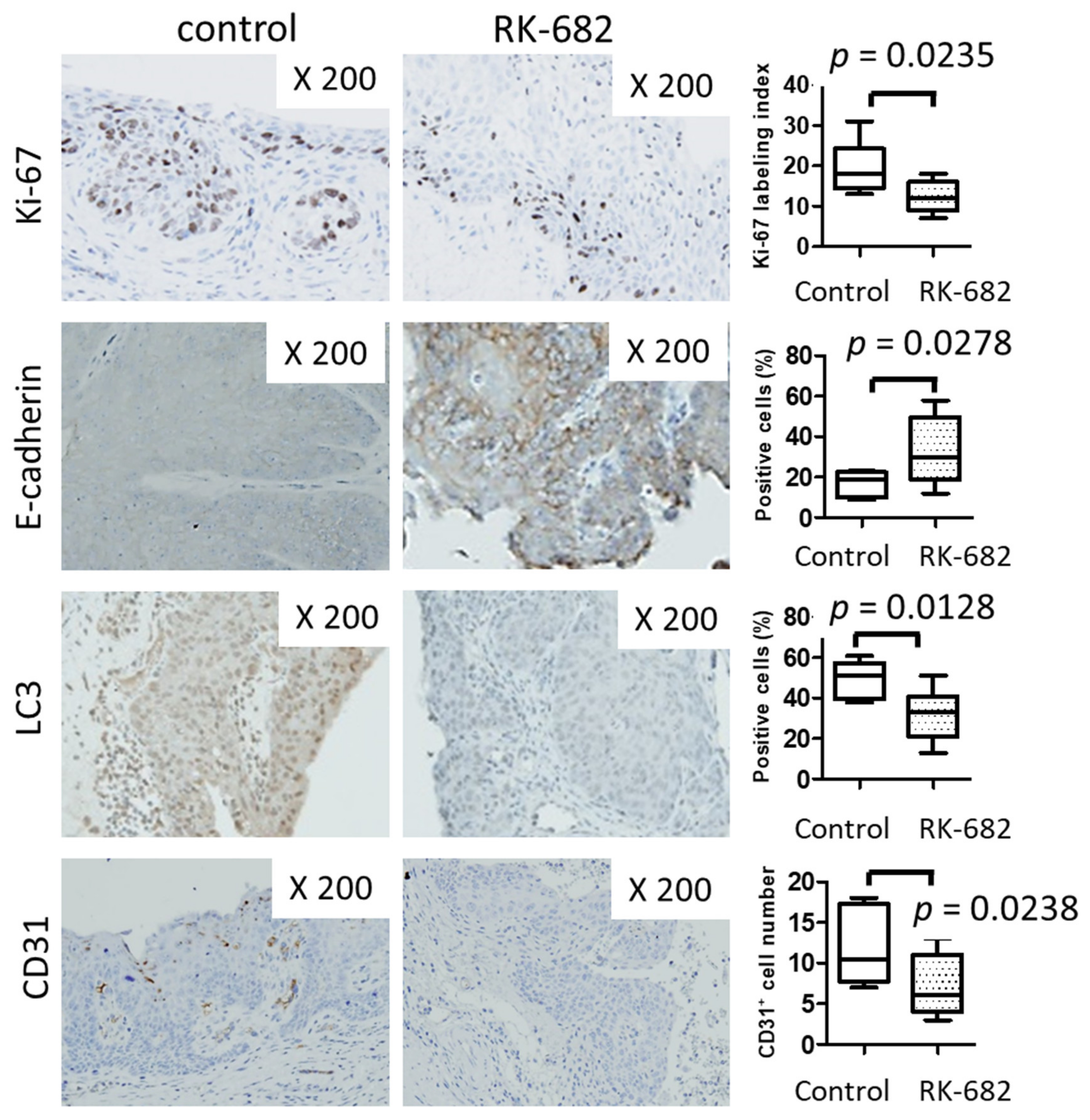
2.4. In Vivo Growth of Urothelial Carcinoma Is Suppressed by RK-682 in the BBN-Induced Mouse Bladder Cancer Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Plasmids and Chemicals
4.2. siRNA Transfection of Heparanase
4.3. Tissue Samples and Immunohistochemistry (IHC)
4.4. Cell Proliferation Assay
4.5. Apoptosis Detection Assay
4.6. TdT-Mediated dUTP Nick End Labeling (TUNEL) Assay
4.7. Cell Viability Assay
4.8. Migration Assay
4.9. Autophagy Assay
4.10. BBN-Induced Mouse Bladder Cancer Model
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AV | Annexin V |
| FITC | Fluorescein-5-isothiocyanate |
| IC50 | half-maximal inhibitory concentration |
| IHC | Immunohistochemistry |
| PBS | phosphate-buffered saline |
| PI | propidium iodide |
| siRNA | Small Interfering Ribonucleic Acid |
| qPCR | Quantitative Polymerase Chain Reaction |
| TUNEL | TdT-mediated dUTP nick end labeling |
| TURBT | Transurethral resection of the bladder tumor |
References
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3, a004952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, E.; Khurana, A.; Shridhar, V.; Dredge, K. The Role of heparanase and sulfatases in the modification of heparan sulfate proteoglycans within the tumor microenvironment and opportunities for novel cancer therapeutics. Front. Oncol. 2014, 4, 195. [Google Scholar] [CrossRef] [PubMed]
- Barash, U.; Cohen-Kaplan, V.; Dowek, I.; Sanderson, R.D.; Ilan, N.; Vlodavsky, I. Proteoglycans in health and disease: New concepts for heparanase function in tumor progression and metastasis. FEBS J. 2010, 277, 3890–3903. [Google Scholar] [CrossRef] [Green Version]
- Dhodapkar, M.V.; Abe, E.; Theus, A.; Lacy, M.; Langford, J.K.; Barlogie, B.; Sanderson, R.D. Syndecan-1 is a multifunctional regulator of myeloma pathobiology: Control of tumor cell survival, growth, and bone cell differentiation. Blood 1998, 91, 2679–2688. [Google Scholar] [CrossRef]
- Mahtouk, K.; Cremer, F.W.; Rème, T.; Jourdan, M.; Baudard, M.; Moreaux, J.; Requirand, G.; Fiol, G.; De Vos, J.; Moos, M.; et al. Heparan sulphate proteoglycans are essential for the myeloma cell growth activity of EGF-family ligands in multiple myeloma. Oncogene 2006, 25, 7180–7191. [Google Scholar] [CrossRef] [Green Version]
- Derksen, P.W.; Keehnen, R.M.; Evers, L.M.; Van Oers, M.H.; Spaargaren, M.; Pals, S.T. Cell surface proteoglycan syndecan-1 mediates hepatocyte growth factor binding and promotes Met signaling in multiple myeloma. Blood 2002, 99, 1405–1410. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; MacLeod, V.; Dai, Y.; Khotskaya-Sample, Y.; Shriver, Z.; Venkataraman, G.; Sasisekharan, R.; Naggi, A.; Torri, G.; Casu, B.; et al. The syndecan-1 heparan sulfate proteoglycan is a viable target for myeloma therapy. Blood 2007, 110, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, V.; Koo, C.Y.; Ibrahim, S.A.; Wang, Z.; Spillmann, D.; Dreier, R.; Kelsch, R.; Fischgrabe, J.; Smollich, M.; Rossi, L.H.; et al. Differential roles for membrane-bound and soluble syndecan-1 (CD138) in breast cancer progression. Carcinogenesis 2009, 30, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, K.; Nakamura, M.; Anai, S.; De Velasco, M.; Tanaka, M.; Tsujikawa, K.; Ouji, Y.; Konishi, N. A novel human AlkB homologue, ALKBH8, contributes to human bladder cancer progression. Cancer Res. 2009, 69, 3157–3164. [Google Scholar] [CrossRef] [Green Version]
- Shimada, K.; Matsuyoshi, S.; Nakamura, M.; Ishida, E.; Konishi, N. Phosphorylation status of Fas-associated death domain-containing protein (FADD) is associated with prostate cancer progression. J. Pathol. 2005, 206, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Nakamura, M.; De Velasco, M.A.; Tanaka, M.; Ouji, Y.; Konishi, N. Syndecan-1, a new target molecule involved in progression of androgen-independent prostate cancer. Cancer Sci. 2009, 100, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- Vlodavsky, I.; Ilan, N.; Naggi, A.; Casu, B. Heparanase: Structure, biological functions, and inhibition by heparin-derived mimetics of heparan sulfate. Curr. Pharm. Des. 2007, 13, 2057–2073. [Google Scholar] [CrossRef] [PubMed]
- Vlodavsky, I.; Elkin, M.; Abboud-Jarrous, G.; Levi-Adam, F.; Fuks, L.; Shafat, I.; Ilan, N. Heparanase: One molecule with multiple functions in cancer progression. Connect. Tissue Res. 2008, 49, 207–210. [Google Scholar] [CrossRef]
- Ilan, N.; Elkin, M.; Vlodavsky, I. Regulation, function and clinical significance of heparanase in cancer metastasis and angiogenesis. Int. J. Biochem. Cell Biol. 2006, 38, 2018–2039. [Google Scholar] [CrossRef]
- Whitelock, J.M.; Iozzo, R.V. Heparan sulfate: A complex polymer charged with biological activity. Chem. Rev. 2005, 105, 2745–2764. [Google Scholar] [CrossRef]
- Maeda, T.; Desouky, J.; Friedl, A. Syndecan-1 expression by stromal fibroblasts promotes breast carcinoma growth in vivo and stimulates tumor angiogenesis. Oncogene 2006, 25, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Weissmann, M.; Arvatz, G.; Horowitz, N.; Feld, S.; Naroditsky, I.; Zhang, Y.; Ng, M.; Hammond, E.; Nevo, E.; Vlodavsky, I.; et al. Heparanase-neutralizing antibodies attenuate lymphoma tumor growth and metastasis. Proc. Natl. Acad. Sci. USA 2016, 113, 704–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.F.; Tang, X.D.; Gao, J.H.; Fang, D.C.; Yang, S.M. Heparanase: A universal immunotherapeutic target in human cancers. Drug Discov. Today 2011, 16, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Tang, X.D.; Wan, Y.; Chen, L.; Yu, S.T.; Xiong, Z.; Fang, D.C.; Liang, G.P.; Yang, S.M. HLA-A2-restricted cytotoxic T lymphocyte epitopes from human heparanase as novel targets for broad-spectrum tumor immunotherapy. Neoplasia (N. Y. NY) 2008, 10, 977–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baburajeev, C.P.; Mohan, C.D.; Rangappa, S.; Mason, D.J.; Fuchs, J.E.; Bender, A.; Barash, U.; Vlodavsky, I.; Basappa; Rangappa, K.S. Identification of novel class of triazolo-thiadiazoles as potent inhibitors of human heparanase and their anticancer activity. BMC Cancer 2017, 17, 235. [Google Scholar] [CrossRef] [Green Version]
- Miao, H.Q.; Liu, H.; Navarro, E.; Kussie, P.; Zhu, Z. Development of heparanase inhibitors for anti-cancer therapy. Curr. Med. Chem. 2006, 13, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.R.; Freeman, C.; Brown, K.J.; Francis, D.J.; Cowden, W.B. Identification of sulfated oligosaccharide-based inhibitors of tumor growth and metastasis using novel in vitro assays for angiogenesis and heparanase activity. Cancer Res. 1999, 59, 3433–3441. [Google Scholar] [PubMed]
- Borsig, L. Antimetastatic activities of modified heparins: Selectin inhibition by heparin attenuates metastasis. Semin. Thromb. Hemost. 2007, 33, 540–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlodavsky, I.; Friedmann, Y.; Elkin, M.; Aingorn, H.; Atzmon, R.; Ishai-Michaeli, R.; Bitan, M.; Pappo, O.; Peretz, T.; Michal, I.; et al. Mammalian heparanase: Gene cloning, expression and function in tumor progression and metastasis. Nat. Med. 1999, 5, 793–802. [Google Scholar] [CrossRef]
- Hulett, M.D.; Freeman, C.; Hamdorf, B.J.; Baker, R.T.; Harris, M.J.; Parish, C.R. Cloning of mammalian heparanase, an important enzyme in tumor invasion and metastasis. Nat. Med. 1999, 5, 803–809. [Google Scholar] [CrossRef]
- Parish, C.R.; Freeman, C.; Hulett, M.D. Heparanase: A key enzyme involved in cell invasion. Biochim. Biophys. Acta 2001, 1471, M99–M108. [Google Scholar] [CrossRef]
- Vlodavsky, I.; Friedmann, Y. Molecular properties and involvement of heparanase in cancer metastasis and angiogenesis. J. Clin. Investig. 2001, 108, 341–347. [Google Scholar] [CrossRef]
- Vlodavsky, I.; Beckhove, P.; Lerner, I.; Pisano, C.; Meirovitz, A.; Ilan, N.; Elkin, M. Significance of heparanase in cancer and inflammation. Cancer Microenviron. 2012, 5, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Jung, O.; Trapp-Stamborski, V.; Purushothaman, A.; Jin, H.; Wang, H.; Sanderson, R.D.; Rapraeger, A.C. Heparanase-induced shedding of syndecan-1/CD138 in myeloma and endothelial cells activates VEGFR2 and an invasive phenotype: Prevention by novel synstatins. Oncogenesis 2016, 5, e202. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-P.; Luo, J.-S.; Tian, Y.; Nie, C.-L.; Cui, W.; Zhang, W.-D. Downregulation of heparanase expression results in suppression of invasion, migration, and adhesion abilities of hepatocellular carcinoma cells. BioMed Res. Int. 2015, 2015, 241983. [Google Scholar] [CrossRef]
- Comperat, E.M.; Burger, M.; Gontero, P.; Mostafid, A.H.; Palou, J.; Roupret, M.; van Rhijn, B.W.G.; Shariat, S.F.; Sylvester, R.J.; Zigeuner, R.; et al. Grading of urothelial carcinoma and the new “world health organisation classification of tumours of the urinary system and male genital organs 2016”. Eur. Urol. Focus 2019, 5, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Gohji, K.; Okamoto, M.; Kitazawa, S.; Toyoshima, M.; Dong, J.; Katsuoka, Y.; Nakajima, M. Heparanase protein and gene expression in bladder cancer. J. Urol. 2001, 166, 1286–1290. [Google Scholar] [CrossRef]
- Shteingauz, A.; Boyango, I.; Naroditsky, I.; Hammond, E.; Gruber, M.; Doweck, I.; Ilan, N.; Vlodavsky, I. Heparanase enhances tumor growth and chemoresistance by promoting autophagy. Cancer Res. 2015, 75, 3946–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabichi, A.; Keyhani, A.; Tanaka, N.; Delacerda, J.; Lee, I.L.; Zou, C.; Zhou, J.H.; Benedict, W.F.; Grossman, H.B. Characterization of a panel of cell lines derived from urothelial neoplasms: Genetic alterations, growth in vivo and the relationship of adenoviral mediated gene transfer to coxsackie adenovirus receptor expression. J. Urol. 2006, 175, 1133–1137. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pTa (n = 20) | pT1 (n = 17) | pTis (n = 10) | |
|---|---|---|---|
| Age | 71.3 (61–82) | 72.9 (5–80) | 72.4 (62–86) |
| Gender (M:F) | 16:4 | 14:3 | 8:2 |
| Grade | |||
| Low grade | 13 | 3 | 0 |
| High grade | 7 | 14 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tatsumi, Y.; Miyake, M.; Shimada, K.; Fujii, T.; Hori, S.; Morizawa, Y.; Nakai, Y.; Anai, S.; Tanaka, N.; Konishi, N.; et al. Inhibition of Heparanase Expression Results in Suppression of Invasion, Migration and Adhesion Abilities of Bladder Cancer Cells. Int. J. Mol. Sci. 2020, 21, 3789. https://doi.org/10.3390/ijms21113789
Tatsumi Y, Miyake M, Shimada K, Fujii T, Hori S, Morizawa Y, Nakai Y, Anai S, Tanaka N, Konishi N, et al. Inhibition of Heparanase Expression Results in Suppression of Invasion, Migration and Adhesion Abilities of Bladder Cancer Cells. International Journal of Molecular Sciences. 2020; 21(11):3789. https://doi.org/10.3390/ijms21113789
Chicago/Turabian StyleTatsumi, Yoshihiro, Makito Miyake, Keiji Shimada, Tomomi Fujii, Shunta Hori, Yosuke Morizawa, Yasushi Nakai, Satoshi Anai, Nobumichi Tanaka, Noboru Konishi, and et al. 2020. "Inhibition of Heparanase Expression Results in Suppression of Invasion, Migration and Adhesion Abilities of Bladder Cancer Cells" International Journal of Molecular Sciences 21, no. 11: 3789. https://doi.org/10.3390/ijms21113789
APA StyleTatsumi, Y., Miyake, M., Shimada, K., Fujii, T., Hori, S., Morizawa, Y., Nakai, Y., Anai, S., Tanaka, N., Konishi, N., & Fujimoto, K. (2020). Inhibition of Heparanase Expression Results in Suppression of Invasion, Migration and Adhesion Abilities of Bladder Cancer Cells. International Journal of Molecular Sciences, 21(11), 3789. https://doi.org/10.3390/ijms21113789