3D Culture of MSCs on a Gelatin Microsphere in a Dynamic Culture System Enhances Chondrogenesis

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
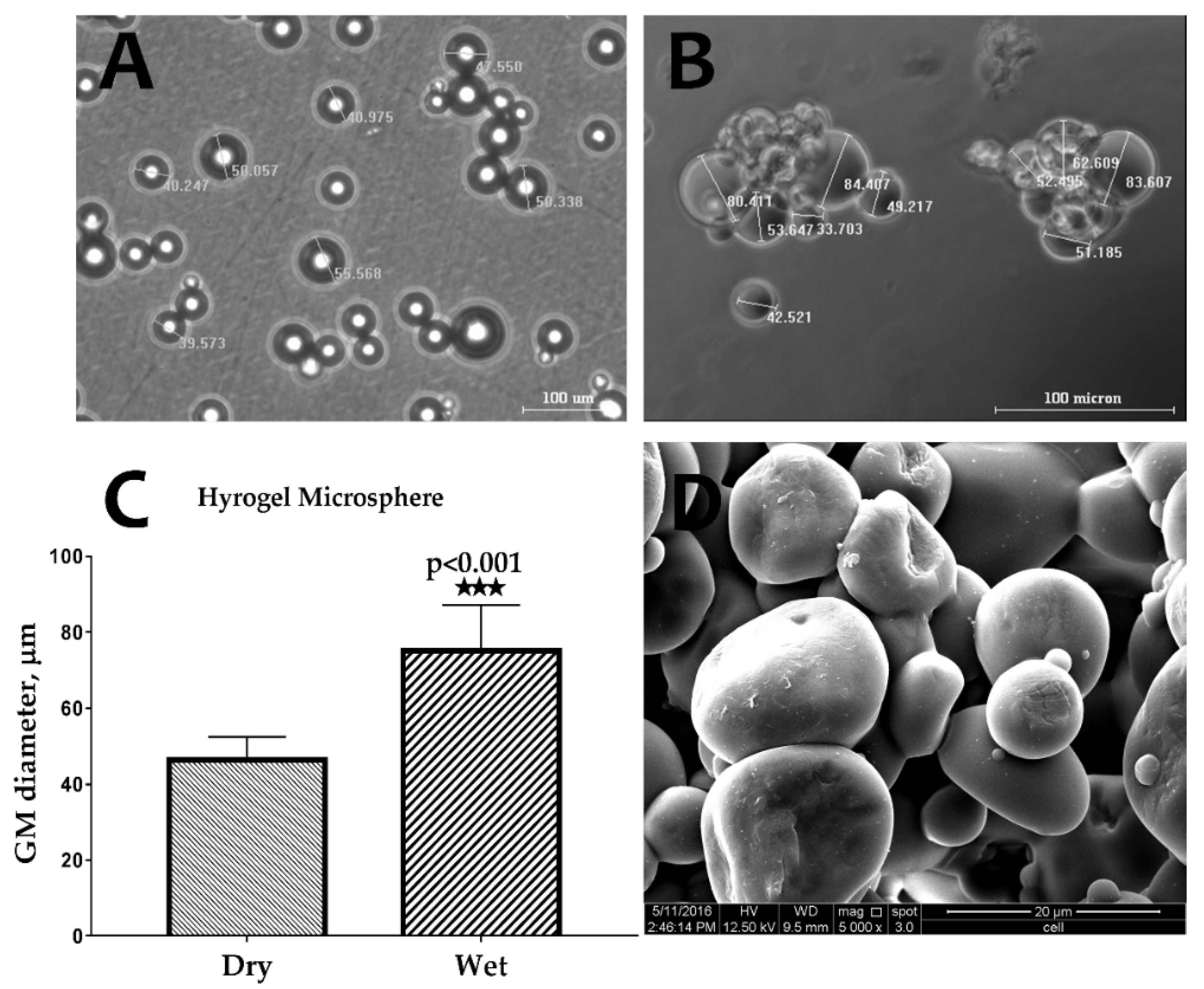
2.1. Characterization of Gelatin Microspheres
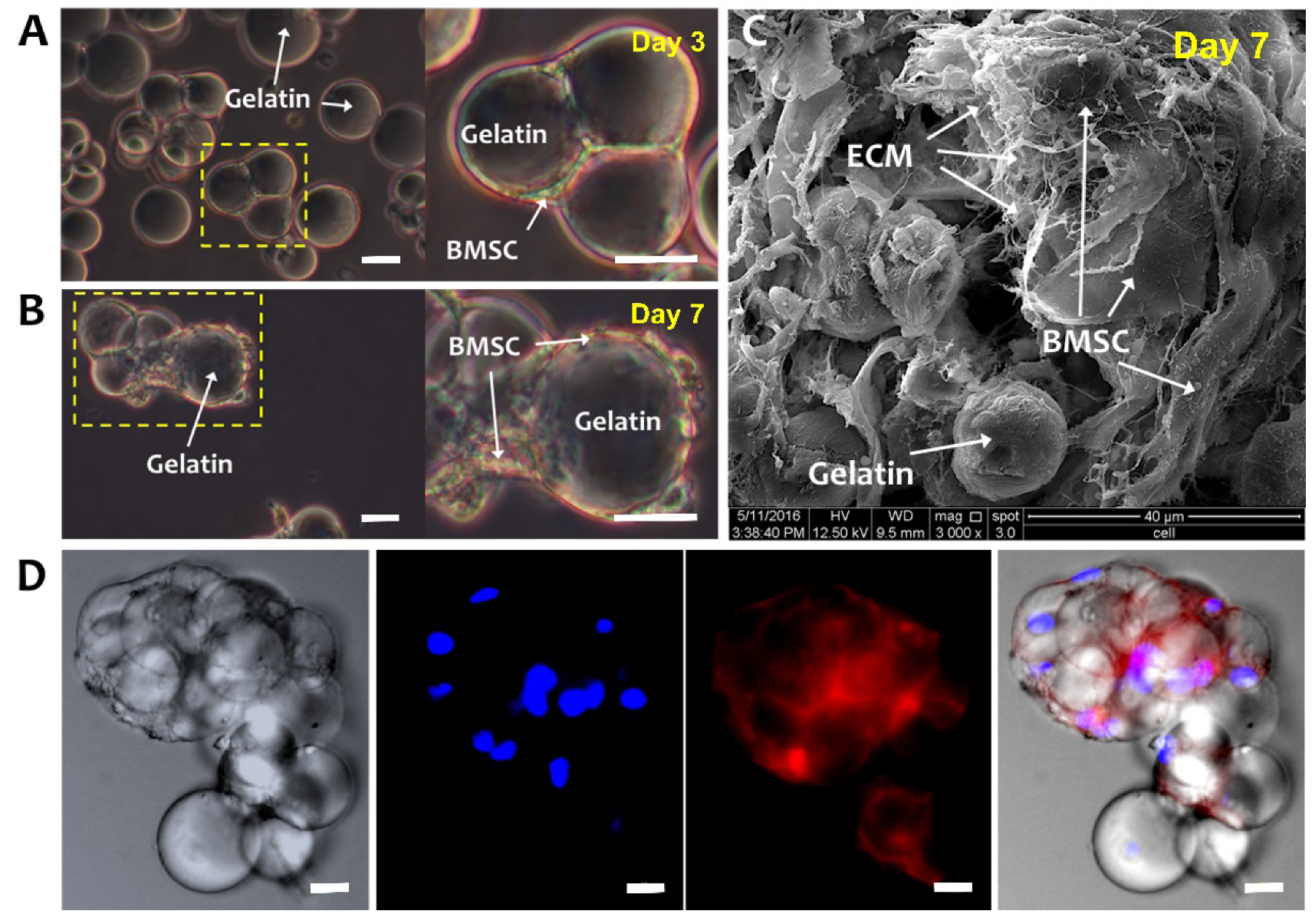
2.2. Morphology of BMSCs on a Gelatin Microsphere
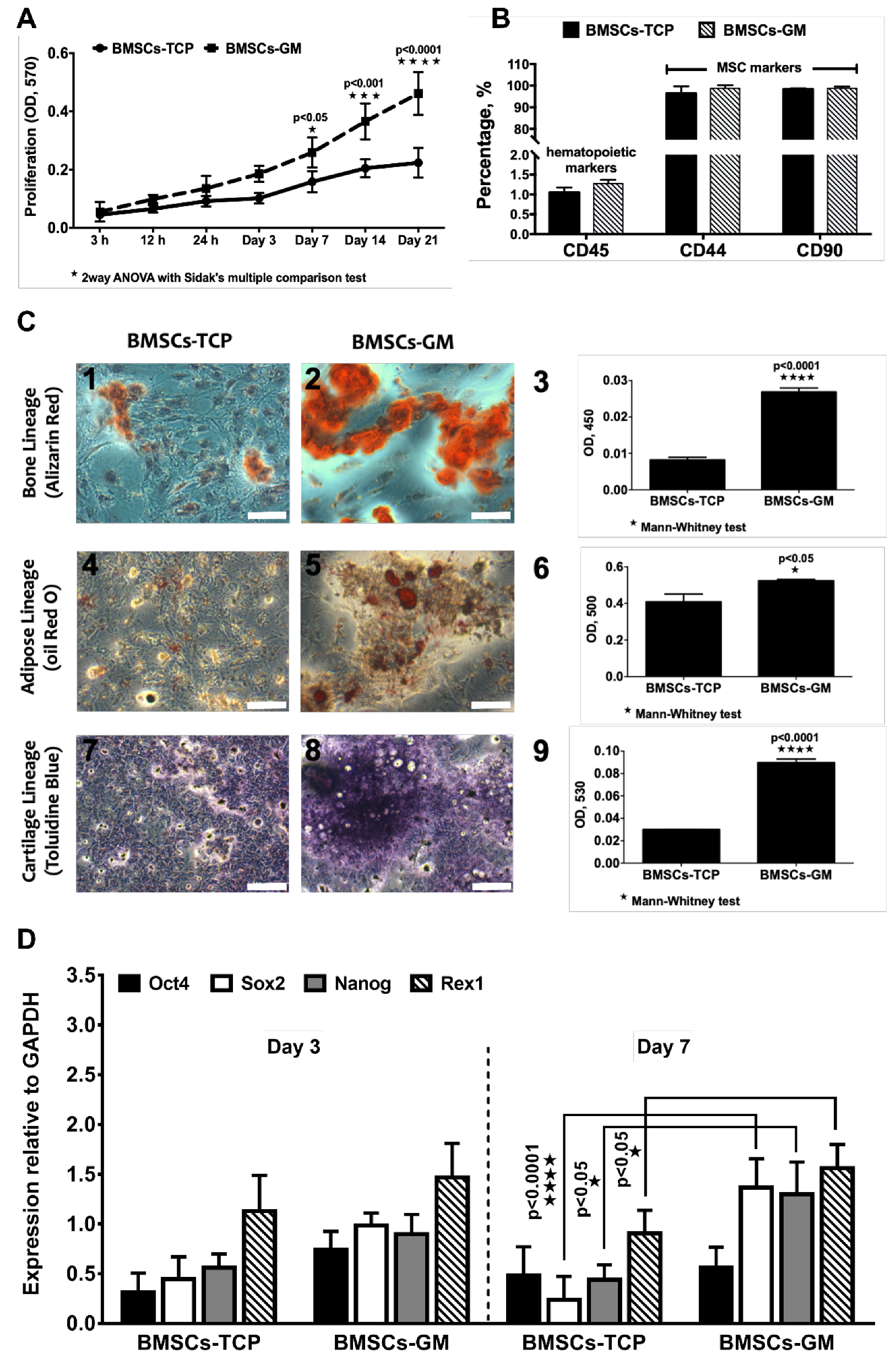
2.3. Proliferation, Characterization, and Differentiation of BMSCs-TCP vs. BMSCs-GM
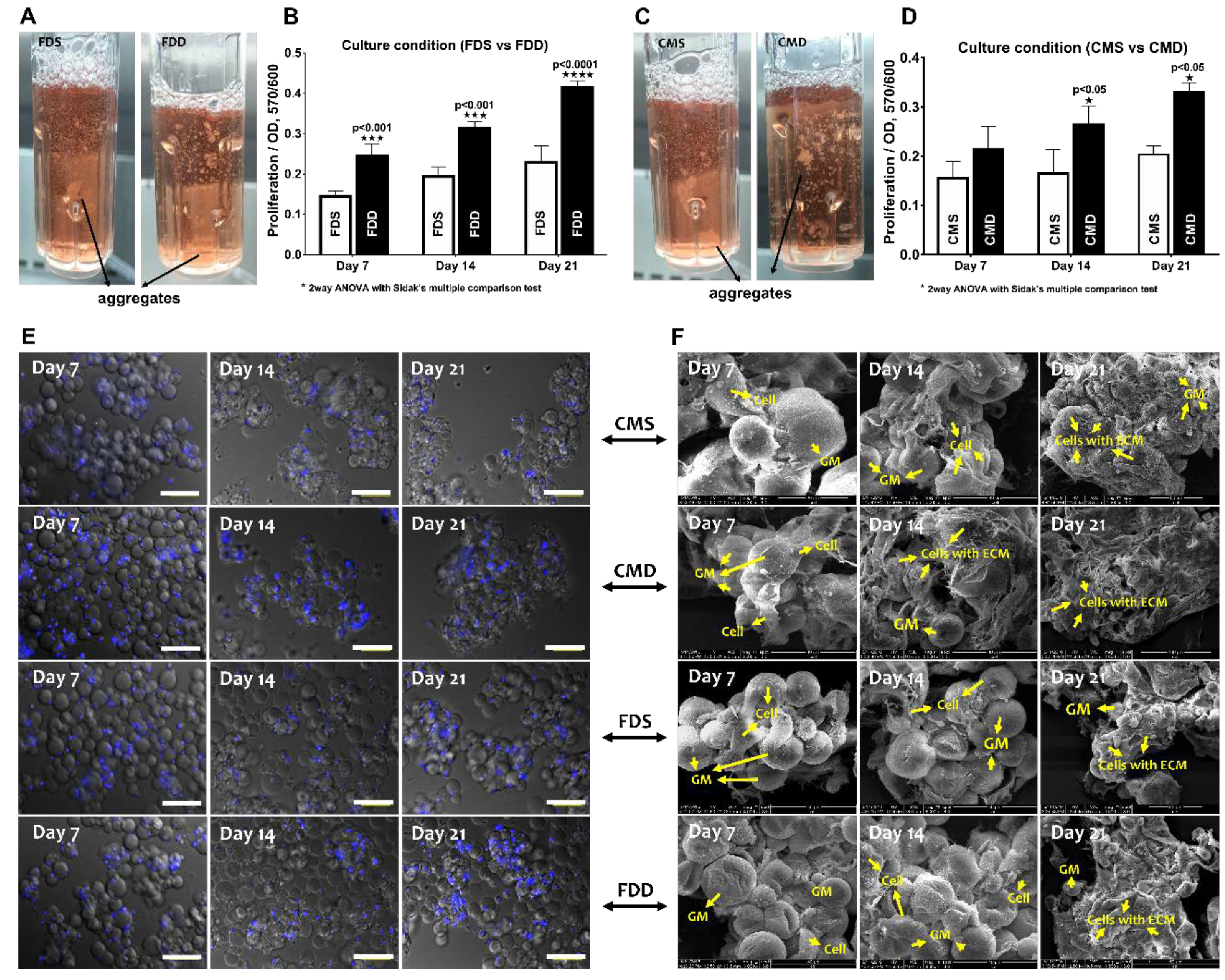
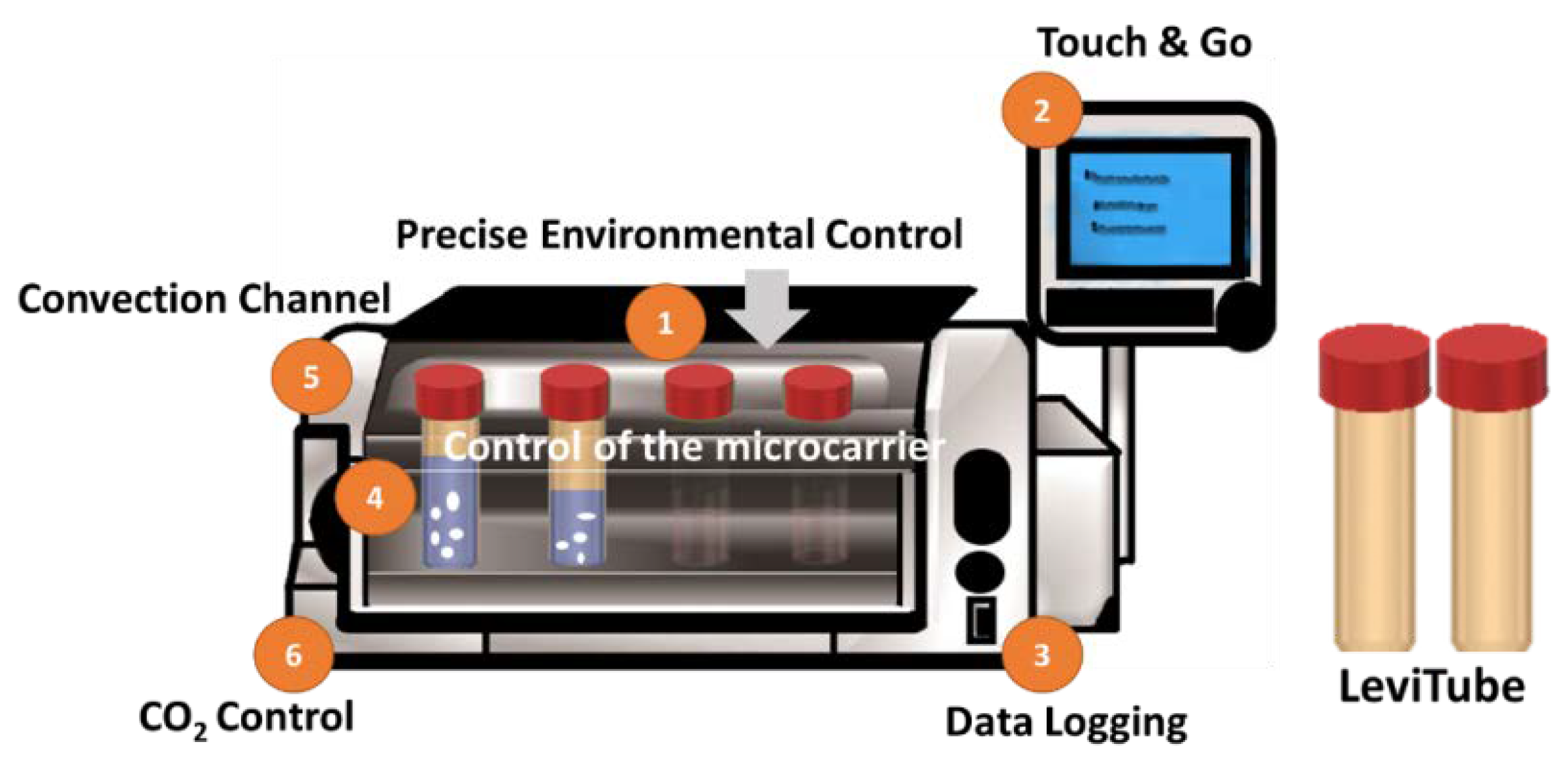
2.4. Attachment and Proliferation of BMSCs-GM in a Static vs. Dynamic Culture Environment
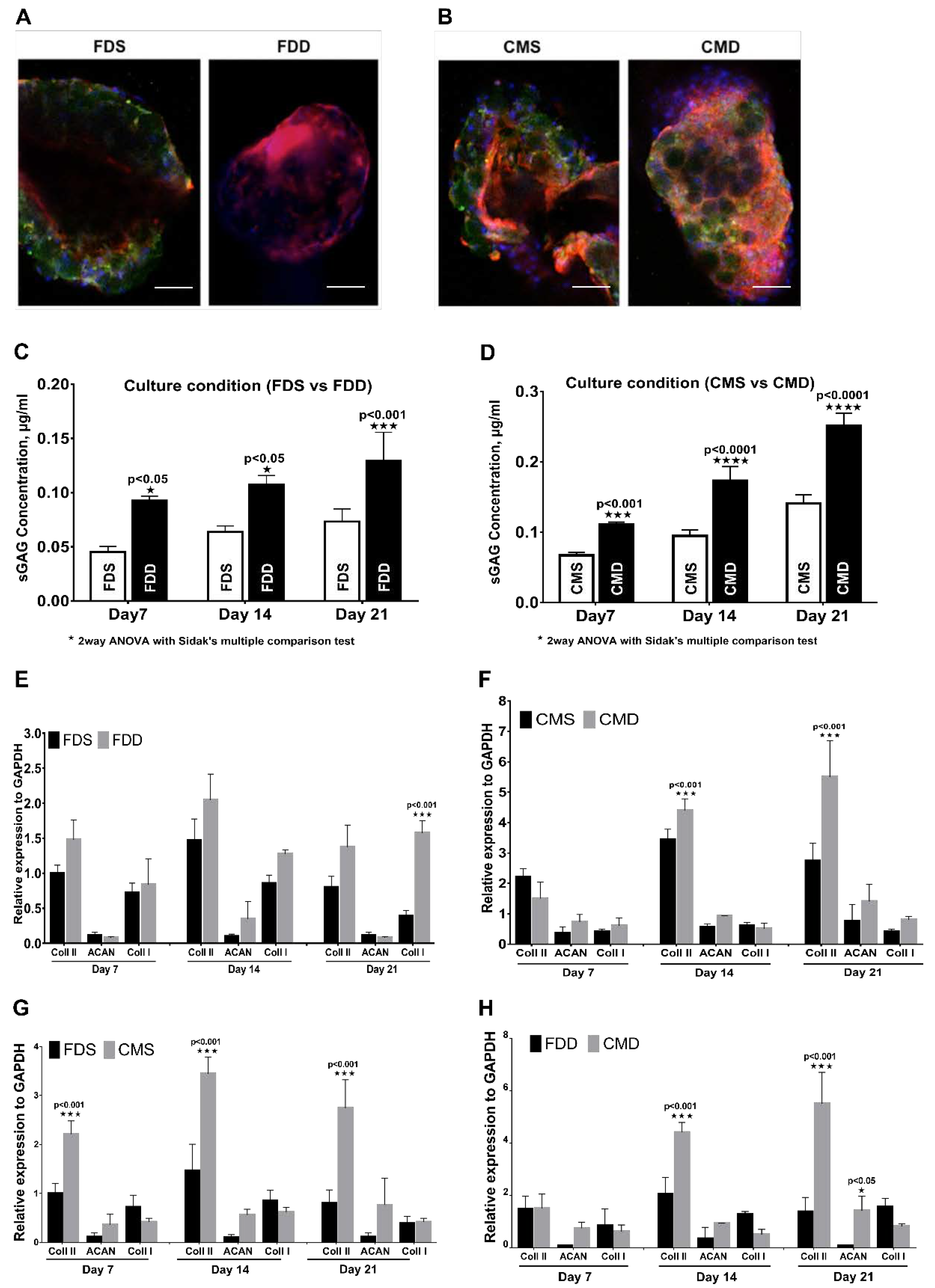
2.5. Chondrogenic Differentiation of BMSCs-GM in a Static vs. Dynamic Culture Environment
3. Discussion
4. Materials and Methods
4.1. Isolation of BMSCs and Cell Culture
4.2. Preparation and Characterization of Gelatin Microspheres (GM)
4.3. Cell Seeding on Gelatin Microspheres and Tissue Culture Plate (TCP)
4.4. Immunophenotype Analysis of BMSCs-TCP vs. BMSCs-GM by Flow Cytometry
4.5. Real-Time Polymerase Chain Reaction (qPCR)
4.6. Proliferation Assay on BMSCs-TCP vs. BMSCs-GM
4.7. Trilineage Differentiation of BMSCs-TCP vs. BMSCs-GM
4.8. Chondrogenic Differentiation of BMSCs-GM in a Static vs. Dynamic Culture Environment
4.9. Immunofluorescence Staining
4.10. Sulfated Glycosaminoglycan (sGAG) Production Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3D | Three-Dimensional |
| 2D | Two-Dimensional |
| BMSCs | Bone-Marrow-Derived Mesenchymal Stem Cells |
| GMs | Gelatin Microspheres |
| MSC | Mesenchymal Stem Cells |
| FBS | Fetal Bovine Serum |
| SEM | Scanning Electron Microscope |
| TCP | Tissue Culture Plate |
| PVA | Polyvinyl Alcohol |
| PBS | Phosphate Buffer Saline |
| BSA | Bovine Serum Albumin |
| CIM | Chondrogenic Induction Medium |
| ITS | Insulin Transferring Selenium |
| TGF-β3 | Transforming Growth Factor Beta 3 |
| IGF-1 | Insulin-Like Growth Factor 1 |
| DMMB | 1,9-Dimethyl Methylene Blue |
| CLSM | Confocal Laser Scanning Microscopy |
| DAPI | 4′,6-diamidino-2-phenylindole |
References
- Dzobo, K.; Thomford, N.E.; Senthebane, D.A.; Shipanga, H.; Rowe, A.; Dandara, C.; Pillay, M.; Shirley, K.; Motaung, C.M. Innovation and Transformation of Medicine. Stem Cells Int. 2018, 2018, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Leduc, T.; Hervieu, M.; Legendre, F.; Bouyoucef, M.; Gruchy, N.; Poulain, L.; De Vienne, C.; Herlicoviez, M.; Demoor, M.; Galéra, P. Chondrogenic commitment of human umbilical cord blood-derived mesenchymal stem cells in collagen matrices for cartilage engineering. Sci. Rep. 2016, 6, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arslan, E.; Guler, M.O.; Tekinay, A.B. Glycosaminoglycan-Mimetic Signals Direct the Osteo/Chondrogenic Differentiation of Mesenchymal Stem Cells in a Three-Dimensional Peptide Nanofiber Extracellular Matrix Mimetic Environment. Biomacromolecules 2016, 17, 1280–1291. [Google Scholar] [CrossRef] [PubMed]
- Fong, C.Y.; Subramanian, A.; Gauthaman, K.; Venugopal, J.; Biswas, A.; Ramakrishna, S.; Bongso, A. Human Umbilical Cord Wharton’s Jelly Stem Cells Undergo Enhanced Chondrogenic Differentiation when Grown on Nanofibrous Scaffolds and in a Sequential Two-stage Culture Medium Environment. Stem Cell Rev. Reports 2012, 8, 195–209. [Google Scholar] [CrossRef]
- Negoro, T.; Takagaki, Y.; Okura, H.; Matsuyama, A. Trends in clinical trials for articular cartilage repair by cell therapy. NPJ Regen. Med. 2018, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Najar, M.; Bouhtit, F.; Melki, R.; Afif, H.; Hamal, A.; Fahmi, H.; Merimi, M.; Lagneaux, L. Mesenchymal Stromal Cell-Based Therapy: New Perspectives and Challenges. J. Clin. Med. 2019, 8, 626. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.T.; Ting, C.H.; Yen, M.L.; Liu, K.J.; Sytwu, H.K.; Wu, K.K.; Yen, B.L. Human mesenchymal stem cells (MSCs) for treatment towards immune- and inflammation-mediated diseases: Review of current clinical trials. J. Biomed. Sci. 2016, 23, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Emadedin, M.; Liastani, M.G.; Fazeli, R.; Mohseni, F.; Moghadasali, R.; Mardpour, S.; Hosseini, S.E.; Niknejadi, M.; Moeininia, F.; Fanni, A.A.; et al. Long-term follow-up of intra-articular injection of autologous mesenchymal stem cells in patients with knee, ankle, or hip osteoarthritis. Arch. Iran. Med. 2015, 18, 336–344. [Google Scholar]
- Jo, C.H.; Chai, J.W.; Jeong, E.C.; Oh, S.; Shin, J.S.; Shim, H.; Yoon, K.S. Intra-articular Injection of Mesenchymal Stem Cells for the Treatment of Osteoarthritis of the Knee: A 2-Year Follow-up Study. Am. J. Sports Med. 2017, 45, 2774–2783. [Google Scholar] [CrossRef]
- Orozco, L.; Munar, A.; Soler, R.; Alberca, M.; Soler, F.; Huguet, M.; Sentís, J.; Sánchez, A.; García-Sancho, J. Treatment of knee osteoarthritis with autologous mesenchymal stem cells: A pilot study. Transplantation 2013, 95, 1535–1541. [Google Scholar] [CrossRef]
- Banfi, A.; Muraglia, A.; Dozin, B. Proliferation kinetics and differentiation potential of ex vivo expanded human bone marrow stromal cells: Implications for their use in cell therapy. Exp. Hematol. 2000, 28, 707–715. [Google Scholar] [CrossRef]
- Justice, B.A.; Badr, N.A.; Felder, R.A. 3D cell culture opens new dimensions in cell-based assays. Drug Discov. Today 2009, 14, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Reiser, J.; Zhang, X.Y.; Hemenway, C.S.; Mondal, D.; Pradhan, L.; La Russa, V.F. Potential of mesenchymal stem cells in gene therapy approaches for inherited and acquired diseases. Expert Opin. Biol. Ther. 2005, 5, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Chang, N.J.; Jhung, Y.R.; Issariyakul, N.; Yao, C.K.; Yeh, M.L. Synergistic stimuli by hydrodynamic pressure and hydrophilic coating on PLGA scaffolds for extracellular matrix synthesis of engineered cartilage. J. Biomater. Sci. Polym. Ed. 2012, 23, 2133–2151. [Google Scholar] [CrossRef]
- Cukierman, E.; Pankov, R.; Stevens, D.R.; Yamada, K.M. Taking Cell-Matrix Adhesions to the Third Dimension. Science 2016, 294, 1708–1712. [Google Scholar] [CrossRef]
- Nakaguchi, K.; Jinnou, H.; Kaneko, N.; Sawada, M.; Hikita, T.; Saitoh, S.; Tabata, Y.; Sawamoto, K. Growth factors released from gelatin hydrogel microspheres increase new neurons in the adult mouse brain. Stem Cells Int. 2012, 2012, 915160. [Google Scholar] [CrossRef] [Green Version]
- Ozeki, M.; Tabata, Y. In vivo degradability of hydrogels prepared from different gelatins by various cross-linking methods. J. Biomater. Sci. Polym. Ed. 2005, 16, 549–561. [Google Scholar] [CrossRef]
- Shamsul, B.S.; Tan, K.K.; Chen, H.C.; Aminuddin, B.S.; Ruszymah, B.H.I. Posterolateral spinal fusion with ostegenesis induced BMSC seeded TCP/HA in a sheep model. Tissue Cell 2014, 46, 152–158. [Google Scholar] [CrossRef]
- Sulaiman, S.B.; Keong, T.K.; Cheng, C.H.; Saim, A.B.; Idrus, R.B.H. Tricalcium phosphate/hydroxyapatite (TCP-HA) bone scaffold as potential candidate for the formation of tissue engineered bone. Indian J. Med. Res. 2013, 137, 1093–1101. [Google Scholar]
- Csd, L.; Am, N.; Ea, W.; Oa, B.; Bd, B.; Schwartz, Z. Adipose Stem Cell Microbeads as Production Sources for Chondrogenic Growth Factors. J. Stem Cells Regen. Med. 2014, 10, 38–48. [Google Scholar]
- Meyer, C.; Stenberg, L.; Gonzalez-Perez, F.; Wrobel, S.; Ronchi, G.; Udina, E.; Suganuma, S.; Geuna, S.; Navarro, X.; Dahlin, L.B.; et al. Chitosan-film enhanced chitosan nerve guides for long-distance regeneration of peripheral nerves. Biomaterials 2016, 76, 33–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Kallos, M.S.; Christopher, H.; Arindom, S. Improved expansion of human bone marrow-derived mesenchymal stem cells in microcarrier-based suspension culture. J. Tissue Eng. Regen. Med. 2012, 8, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Tharmalingam, T.; Sunley, K.; Spearman, M.; Butler, M. Enhanced production of human recombinant proteins from CHO cells grown to high densities in macroporous microcarriers. Mol. Biotechnol. 2011, 49, 263–276. [Google Scholar] [CrossRef]
- Leong, W.; Wang, D.A. Cell-laden Polymeric Microspheres for Biomedical Applications. Trends Biotechnol. 2015, 33, 653–666. [Google Scholar] [CrossRef]
- Tsai, A.C.; Ma, T. Expansion of Human Mesenchymal Stem Cells in a Microcarrier Bioreactor. Methods Mol. Biol. 2016, 257–284. [Google Scholar]
- Merten, O.W. Advances in cell culture: Anchorage dependence. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140040. [Google Scholar] [CrossRef] [Green Version]
- Malda, J.; Frondoza, C.G. Microcarriers in the engineering of cartilage and bone. Trends Biotechnol. 2006, 24, 299–304. [Google Scholar] [CrossRef]
- Leong, W.; Lau, T.T.; Wang, D.A. A temperature-cured dissolvable gelatin microsphere-based cell carrier for chondrocyte delivery in a hydrogel scaffolding system. Acta Biomater. 2013, 9, 6459–6467. [Google Scholar] [CrossRef]
- Hayashi, K.; Tabata, Y. Preparation of stem cell aggregates with gelatin microspheres to enhance biological functions. Acta Biomater. 2011, 7, 2797–2803. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Akazawa, T.; Tabata, Y. In vitro proliferation and chondrogenic differentiation of rat bone marrow stem cells cultured with gelatin hydrogel microspheres for TGF-β1 release. J. Biomater. Sci. Polym. Ed. 2010, 21, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Gohi, B.F.C.A.; Liu, X.Y.; Zeng, H.Y.; Xu, S.; Ake, K.M.H.; Cao, X.J.; Zou, K.M.; Namulondo, S. Enhanced efficiency in isolation and expansion of hAMSCs via dual enzyme digestion and micro-carrier. Cell Biosci. 2020, 10, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Yu, L.; Lim, C.G.; Goodley, A.S.; Xiao, X.; Placone, J.K.; Ferlin, K.M.; Nguyen, B.N.B.; Hsieh, A.H.; Fisher, J.P. Effect of Dynamic Culture and Periodic Compression on Human Mesenchymal Stem Cell Proliferation and Chondrogenesis. Ann. Biomed. Eng. 2016, 44, 2103–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, A.; Mehta, S.; Gupta, D.; Sheikh, S.; Pallagatti, S.; Singh, R.; Singla, I. Clinical & immunological erythematosus patients characteristics in systemic lupus Maryam. J. Dent. Educ. 2012, 76, 1532–1539. [Google Scholar] [PubMed]
- Perez, R.A.; Riccardi, K.; Altankov, G.; Ginebra, M.P. Dynamic cell culture on calcium phosphate microcarriers for bone tissue engineering applications. J. Tissue Eng. 2014, 5, 2041731414543965. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, P.; Pramanik, K.; Biswas, A.; Ku Patra, R. In vitro cartilage construct generation from silk fibroin- chitosan porous scaffold and umbilical cord blood derived human mesenchymal stem cells in dynamic culture condition. J. Biomed. Mater. Res. Part A 2018, 106, 397–407. [Google Scholar] [CrossRef]
- Schop, D.; Janssen, F.W.; Borgart, E.; de Bruijn, J.D.; van Dijkhuizen-Radersma, R. Expansion of mesenchymal stem cells using a microcarrier-based cultivation system: Growth and metabolism. J. Tissue Eng. Regen. Med. 2008, 2, 126–135. [Google Scholar] [CrossRef]
- Dexheimer, V.; Frank, S.; Richter, W. Proliferation as a requirement for in vitro chondrogenesis of human mesenchymal stem cells. Stem Cells Dev. 2012, 21, 2160–2169. [Google Scholar] [CrossRef] [Green Version]
- Ratner, B.D.; Hoffman, A.S.; Schoen, F.J.; Lemons, J.E. Biomaterials Science: An Introduction to Materials in Medicine; Elsevier: Amsterdam, The Netherlands, 2013; pp. 209–222. [Google Scholar]
- Lee, K.C.; Wong, W.K.; Feng, B. Decoding the pluripotency network: The emergence of new transcription factors. Biomedicines 2013, 1, 49–78. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Wang, H.; Pan, G.; Geng, Y.; Guo, Y.; Pei, D. Regulation of the Pluripotency Marker Rex-1 by Nanog and Sox2. J. Biol. Chem. 2006, 281, 23319–23325. [Google Scholar] [CrossRef] [Green Version]
- Cheng, N.C.; Wang, S.; Young, T.H. The influence of spheroid formation of human adipose-derived stem cells on chitosan films on stemness and differentiation capabilities. Biomaterials 2012, 33, 1748–1758. [Google Scholar] [CrossRef]
- Bhandari, D.R.; Seo, K.-W.; Roh, K.-H.; Jung, J.-W.; Kang, S.-K.; Kang, K.-S. REX-1 expression and p38 MAPK activation status can determine proliferation/differentiation fates in human mesenchymal stem cells. PLoS ONE 2010, 5, e10493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drela, K.; Stanaszek, L.; Nowakowski, A.; Kuczynska, Z.; Lukomska, B. Experimental strategies of mesenchymal stem cell propagation: Adverse events and potential risk of functional changes. Stem Cells Int. 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.J.; Tuli, R.; Huang, X.; Laquerriere, P.; Tuan, R.S. Multilineage differentiation of human mesenchymal stem cells in a three-dimensional nanofibrous scaffold. Biomaterials 2005, 26, 5158–5166. [Google Scholar] [CrossRef] [PubMed]
- Leong, D.T.; Abraham, M.C.; Gupta, A.; Lim, T.-C.; Chew, F.T.; Hutmacher, D.W. ATF5, a possible regulator of osteogenic differentiation in human adipose-derived stem cells. J. Cell. Biochem. 2012, 113, 2744–2753. [Google Scholar] [CrossRef]
- Boo, L.; Selvaratnam, L.; Tai, C. Expansion and preservation of multipotentiality of rabbit bone-marrow derived mesenchymal stem cells in dextran-based microcarrier spin culture. J. Mater. Sci. Mater. Med. 2011, 22, 1343–1356. [Google Scholar] [CrossRef]
- Yang, Y.; Rossi, F.M.; Putnins, E.E. Ex vivo expansion of rat bone marrow mesenchymal stromal cells on microcarrier beads in spin culture. Biomaterials 2007, 28, 3110–3120. [Google Scholar] [CrossRef]
- Freed, L.E.; Vunjak-Novakovic, G.; Langer, R. Cultivation of cell-polymer cartilage implants in bioreactors. J. Cell. Biochem. 1993, 51, 257–264. [Google Scholar] [CrossRef]
- Li, W.J.; Jiang, Y.J.; Tuan, R.S. Cell-nanofiber-based cartilage tissue engineering using improved cell seeding, growth factor, and bioreactor technologies. Tissue Eng. Part. A 2008, 14, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Shim, G.; Lee, S.; Han, J.; Kim, G.; Jin, H.; Miao, W.; Yi, T.G.; Cho, Y.K.; Song, S.U.; Oh, Y.K. Pharmacokinetics and in Vivo Fate of Intra-Articularly Transplanted Human Bone Marrow-Derived Clonal Mesenchymal Stem Cells. Stem Cells Dev. 2015, 24, 1124–1132. [Google Scholar] [CrossRef]
- Malda, J.; van den Brink, P.; Meeuwse, P.; Grojec, M.; Martens, D.E.; Tramper, J.; Riesle, J.; van Blitterswijk, C.A. Effect of oxygen tension on adult articular chondrocytes in microcarrier bioreactor culture. Tissue Eng. 2004, 10, 987–994. [Google Scholar] [CrossRef]
- Frondoza, C.; Sohrabi, A.; Hungerford, D. Human chondrocytes proliferate and produce matrix components in microcarrier suspension culture. Biomaterials 1996, 17, 879–888. [Google Scholar] [CrossRef]
- Fischer, B.; Meier, A.; Dehne, A.; Salhotra, A.; Tran, T.A.; Neumann, S.; Schmidt, K.; Meiser, I.; Neubauer, J.C.; Zimmermann, H.; et al. A complete workflow for the differentiation and the dissociation of hiPSC-derived cardiospheres. Stem Cell Res. 2018, 32, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ude, C.C.; Seet, W.T.; Aini, S.S.; Aminuddin, B.S.; Ruszymah, B.H.I. Shelf Life Evaluation of Clinical Grade Chondrogenic Induced Aged Adult Stem Cells for Cartilage Regeneration. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ude, C.C.; Sulaiman, S.B.; Min-Hwei, N.; Hui-Cheng, C.; Ahmad, J.; Yahaya, N.M.; Saim, A.B.; Idrus, R.B.H. Cartilage regeneration by chondrogenic induced adult stem cells in osteoarthritic sheep model. PLoS ONE 2014, 9, e98770. [Google Scholar] [CrossRef]
- Shamsul, B.S.; Chowdhury, S.R.; Norhamdan, M.Y.; Ruszymah, B.H.I. Effect of cell density on formation of three-dimensional cartilaginous constructs using fibrin & human osteoarthritic chondrocytes. Indian J. Med. Res. 2019, 149, 641–649. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward 5′-3′ | Reverse 5′-3′ |
|---|---|---|
| Type 1 Col | AAGGCTTCAAGGTCCCCCTGGTG | CAGCACCAGTAGCACCATCATTTC |
| Type 2 Col | GGCAATAGCAGGTTCACGTACA | CGATAACAGTCTTGCCCCACTT |
| Aggrecan | ACTTCCGCTGGTCAGATGGA | TCTCGTGCCAGATCATCACC |
| GAPDH | GGCGATGCTGGCGCTGAGTAC | TGGTTCACACCCATGACGA |
| Oct4 | GCAGCGACTATGCACAACGA | CCAGAGTGGTGACGGAGACA |
| Sox2 | CATCACCCACAGCAAATGACA | GCTCCTACCGTACCACTAGAACTT |
| Nanog | CCTGTGATTTGTGGGCCTG | GACAGTCTCCGTGTGAGGCAT |
| Rex1 | TGAAAGCCCACATCCTAACG | TATAACCGCTTTTGGGGTTG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulaiman, S.; Chowdhury, S.R.; Fauzi, M.B.; Rani, R.A.; Yahaya, N.H.M.; Tabata, Y.; Hiraoka, Y.; Binti Haji Idrus, R.; Min Hwei, N. 3D Culture of MSCs on a Gelatin Microsphere in a Dynamic Culture System Enhances Chondrogenesis. Int. J. Mol. Sci. 2020, 21, 2688. https://doi.org/10.3390/ijms21082688
Sulaiman S, Chowdhury SR, Fauzi MB, Rani RA, Yahaya NHM, Tabata Y, Hiraoka Y, Binti Haji Idrus R, Min Hwei N. 3D Culture of MSCs on a Gelatin Microsphere in a Dynamic Culture System Enhances Chondrogenesis. International Journal of Molecular Sciences. 2020; 21(8):2688. https://doi.org/10.3390/ijms21082688
Chicago/Turabian StyleSulaiman, Shamsul, Shiplu Roy Chowdhury, Mh Busra Fauzi, Rizal Abdul Rani, Nor Hamdan Mohamad Yahaya, Yasuhiko Tabata, Yosuke Hiraoka, Ruszymah Binti Haji Idrus, and Ng Min Hwei. 2020. "3D Culture of MSCs on a Gelatin Microsphere in a Dynamic Culture System Enhances Chondrogenesis" International Journal of Molecular Sciences 21, no. 8: 2688. https://doi.org/10.3390/ijms21082688
APA StyleSulaiman, S., Chowdhury, S. R., Fauzi, M. B., Rani, R. A., Yahaya, N. H. M., Tabata, Y., Hiraoka, Y., Binti Haji Idrus, R., & Min Hwei, N. (2020). 3D Culture of MSCs on a Gelatin Microsphere in a Dynamic Culture System Enhances Chondrogenesis. International Journal of Molecular Sciences, 21(8), 2688. https://doi.org/10.3390/ijms21082688