Changes in the Cellular Distribution of Tyrosine Phosphorylation and Its Relationship with the Acrosomal Exocytosis and Plasma Membrane Integrity during In Vitro Capacitation of Frozen/Thawed Bull Spermatozoa



Abstract
:1. Introduction
2. Results
2.1. Overall Effects of Processing and Incubation on Analyzed Semen Characteristics
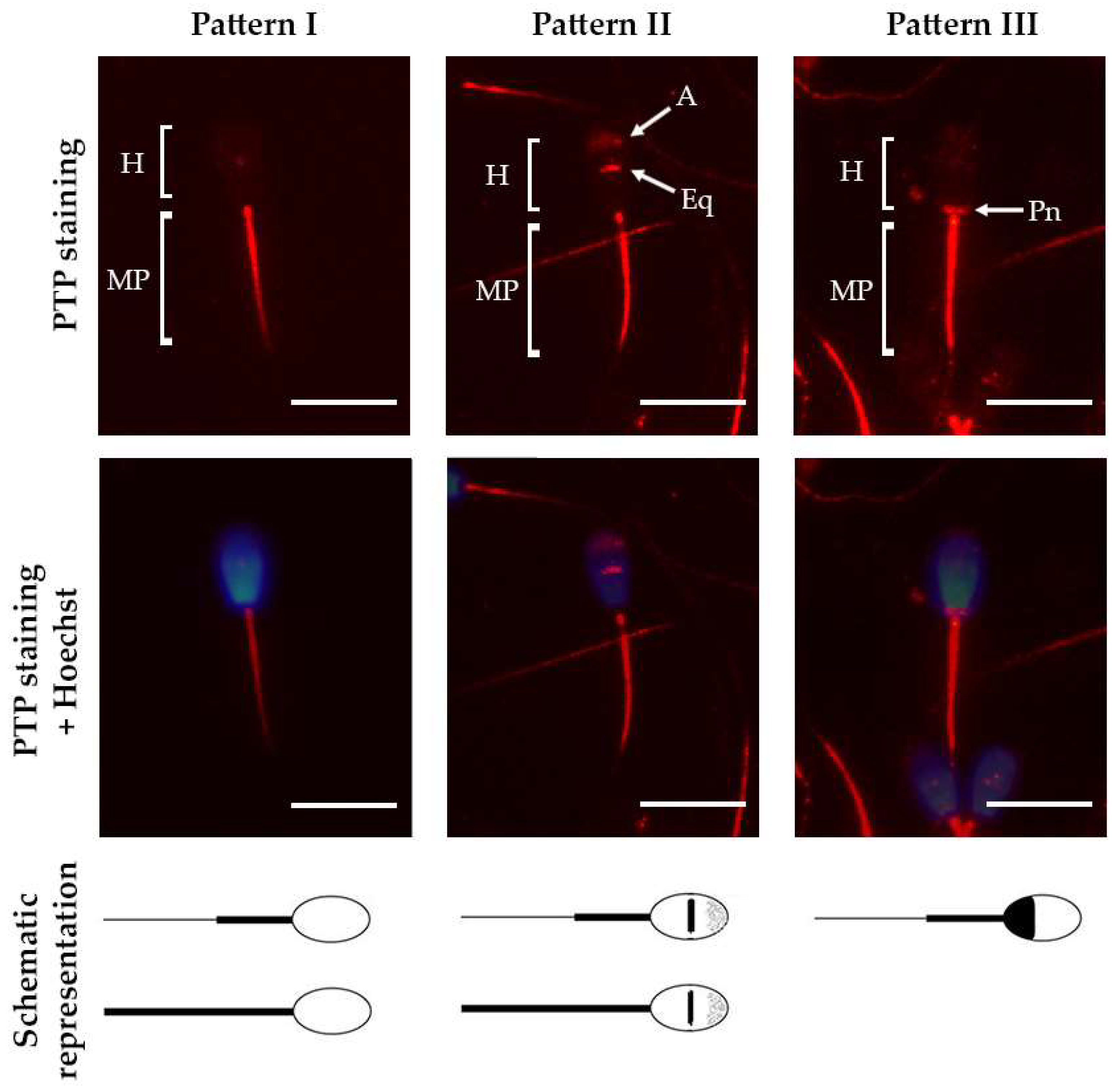
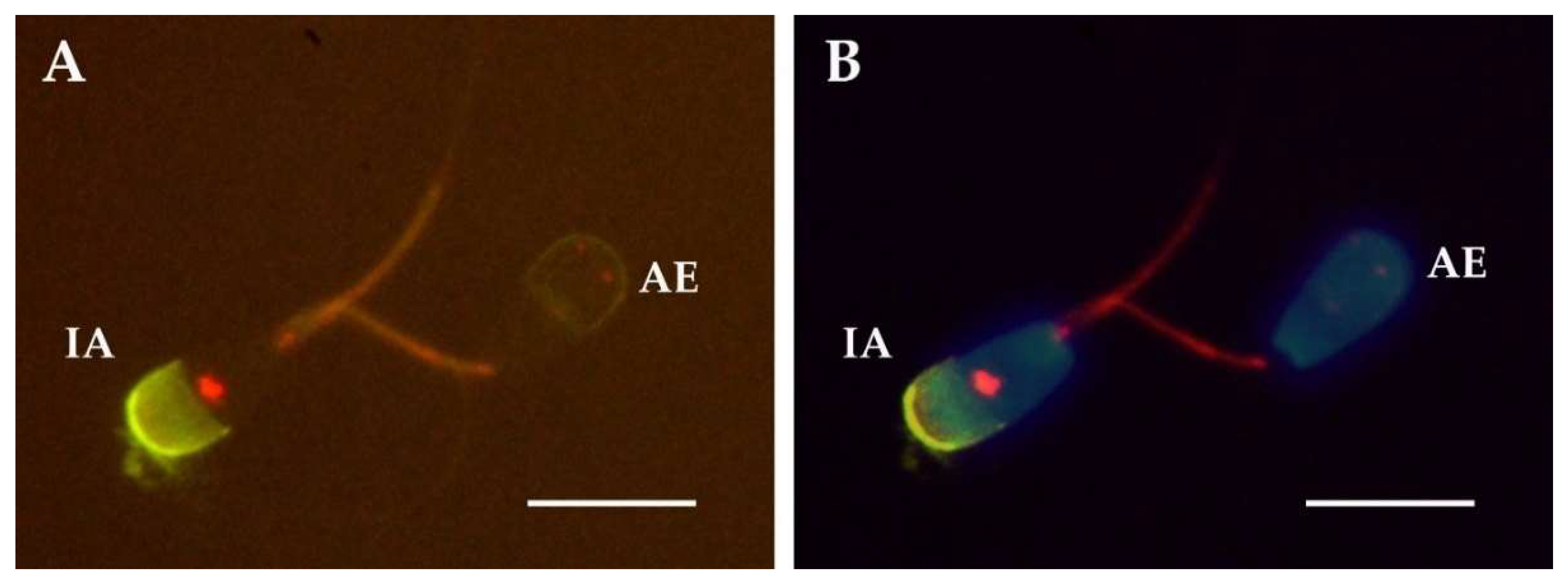
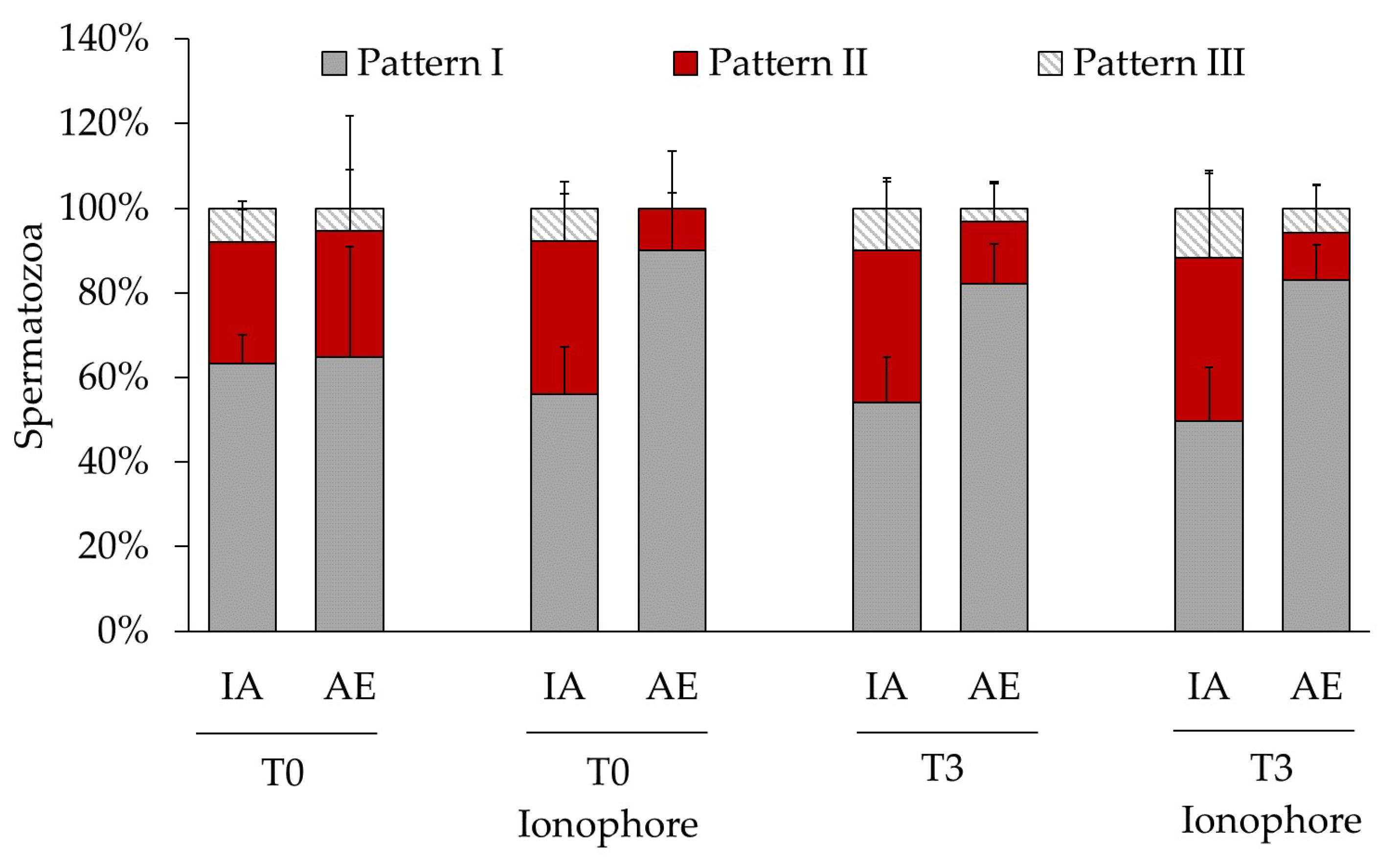
2.2. Relationship between Acrosomal Exocytosis and Cellular Distribution of Protein Tyrosine Phosphorylation
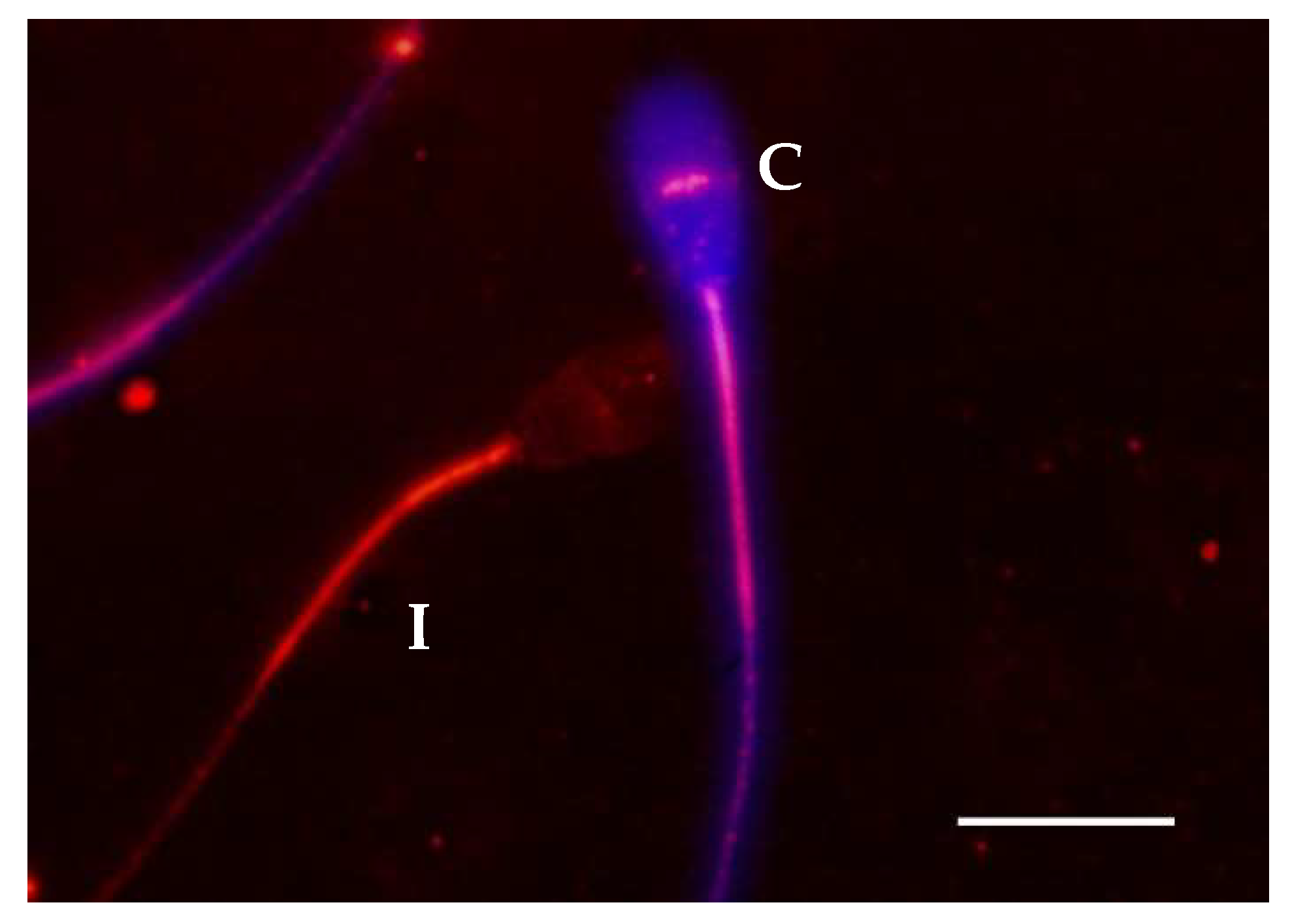
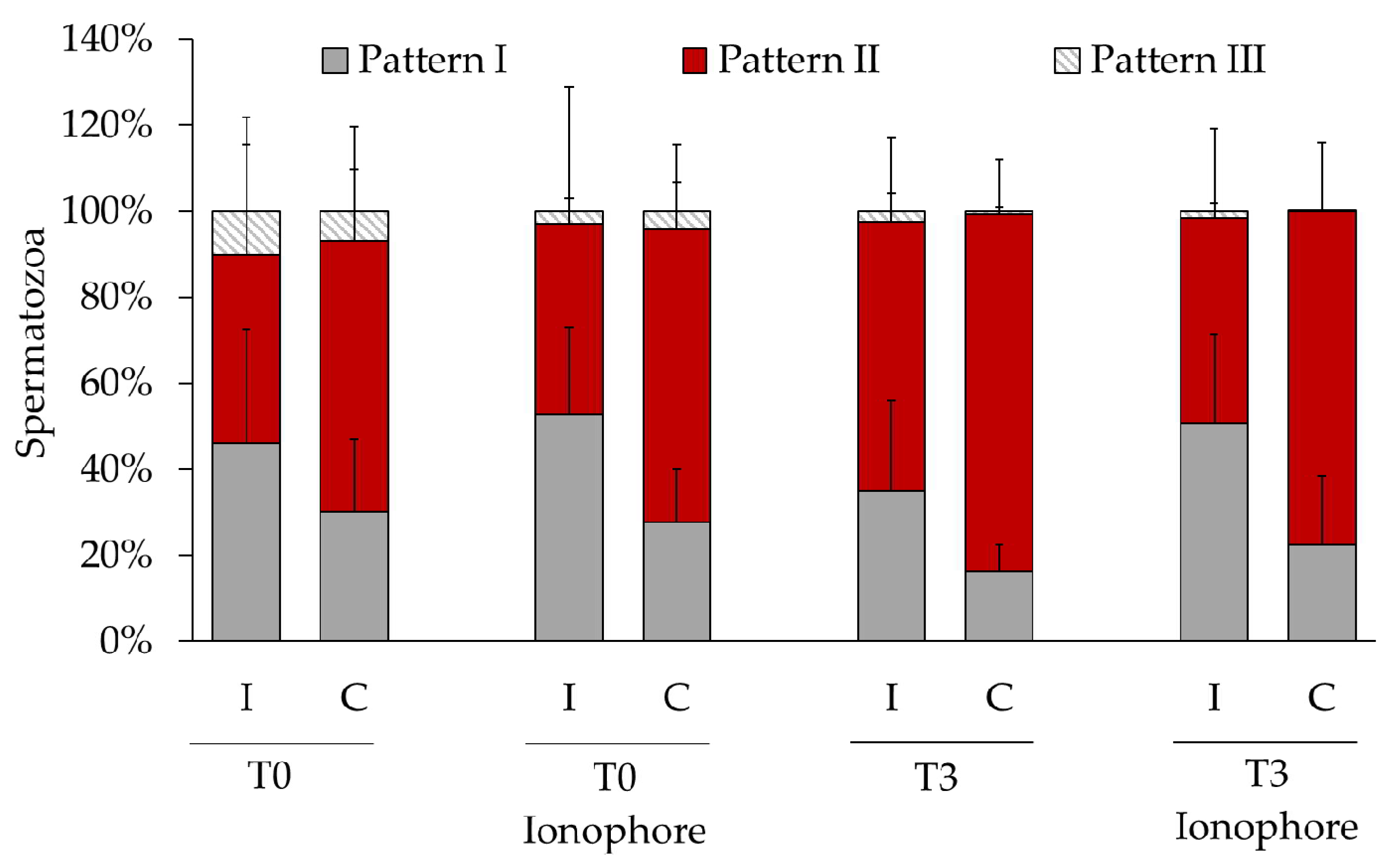
2.3. Relationship between Membrane Integrity and Cellular Distribution of Protein Tyrosine Phosphorylation
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Sperm Processing
4.3. Experimental Design
4.4. Motility Analysis
4.5. Plasma Membrane Integrity Analysis
4.6. Immunofluorescence Analysis of Protein Tyrosine Phosphorylation
4.7. Acrosomal Exocytosis
4.8. Statistics
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PTP | Protein tyrosine phosphorylation |
| I | Intact sperm plasma membrane |
| C | Compromised sperm plasma membrane |
| AE | Acrosomal exocytosis |
| IA | Intact acrosome |
| DGC | Density gradient centrifugation |
| PMI | Plasma membrane integrity |
References
- Visconti, P.E.; Krapf, D.; De La Vega-Beltrán, J.L.; Acevedo, J.J.; Darszon, A. Ion channels, phosphorylation and mammalian sperm capacitation. Asian J. Androl. 2011, 13, 395–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neild, D.N.; Gadella, B.M.; Agüero, A.; Stout, T.A.E.; Colenbrander, B. Capacitation, acrosome function and chromatin structure in stallion sperm. Anim. Reprod. Sci. 2005, 89, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.A.I.; Osycka-salut, C.E.; Castellano, L.; Cesari, A.; Di Siervi, N.; Mutto, A.; Johannisson, A.; Morrell, J.M.; Davio, C.; Perez-Martínez, S. Extracellular cAMP activates molecular signalling pathways associated with sperm capacitation in bovines. MHR: Basic Sci. Reprod. Med. 2017, 23, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Visconti, P.E.; Bailey, J.L.; Moore, G.D.; Pan, D.; Olds-Clarke, P.; Kopf, G.S. Capacitation of mouse spermatozoa. 1. Correlation between the capacitation state and protein tyrosine phosphorylation. Development 1995, 121, 1129–1137. [Google Scholar]
- Gangwar, D.K.; Atreja, S.K. Signalling Events and Associated Pathways Related to the Mammalian Sperm Capacitation. Reprod. Domest. Anim. 2015, 50, 705–711. [Google Scholar] [CrossRef]
- Galantino-Homer, H.L.; Visconti, P.E.; Kopf, G.S. Regulation of protein tyrosine phosphorylation during bovine sperm capacitation by a cyclic adenosine 3′5′-monophosphate-dependent pathway. Biol. Reprod. 1997, 56, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, M.; Doncel, G.F. Involvement of tyrosine kinase and cAMP-dependent kinase cross-talk in the regulation of human sperm motility. Reproduction 2003, 126, 183–195. [Google Scholar] [CrossRef]
- Naresh, S.; Atreja, S.K. The protein tyrosine phosphorylation during in vitro capacitation and cryopreservation of mammalian spermatozoa. Cryobiology 2015, 70, 211–216. [Google Scholar] [CrossRef]
- Grasa, P.; Colas, C.; Gallego, M.; Monteagudo, L.; Muiño-Blanco, T.; Cebrián-Pérez, J.Á. Changes in content and localization of proteins phosphorylated at tyrosine, serine and threonine residues during ram sperm capacitation and acrosome reaction. Reproduction 2009, 137, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Peris-Frau, P.; Martín-Maestro, A.; Iniesta-Cuerda, M.; Sánchez-Ajofrín, I.; Cesari, A.; Garde, J.J.; Villar, M.; Soler, A.J. Cryopreservation of ram sperm alters the dynamic changes associated with in vitro capacitation. Theriogenology 2020, 145, 100–108. [Google Scholar] [CrossRef]
- Urner, F.; Leppens-Luisier, G.; Sakkas, D. Protein Tyrosine Phosphorylation in Sperm During Gamete Interaction in the Mouse: The Influence of Glucose. Biol. Reprod. 2001, 64, 1350–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakkas, D.; Leppens-Luisier, G.; Lucas, H.; Chardonnens, D.; Campana, A.; Franken, D.R.; Urner, F. Localization of Tyrosine Phosphorylated Proteins in Human Sperm and Relation to Capacitation and Zona Pellucida Binding. Biol. Reprod. 2003, 68, 1463–1469. [Google Scholar] [CrossRef] [Green Version]
- Sati, L.; Cayli, S.; Delpiano, E.; Sakkas, D.; Huszar, G. The pattern of tyrosine phosphorylation in human sperm in response to binding to zona pellucida or hyaluronic acid. Reprod. Sci. 2014, 21, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grasa, P.; Cebrián-Pérez, J.Á.; Muiño-Blanco, T. Signal transduction mechanisms involved in in vitro ram sperm capacitation. Reproduction 2006, 132, 721–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luño, V.; López-Úbeda, R.; García-Vázquez, F.A.; Gil, L.; Matás, C. Boar sperm tyrosine phosphorylation patterns in the presence of oviductal epithelial cells: In vitro, ex vivo, and in vivo models. Reproduction 2013, 146, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Pommer, A.C.; Rutllant, J.; Meyers, S.A. Phosphorylation of Protein Tyrosine Residues in Fresh and Cryopreserved Stallion Spermatozoa under Capacitating Conditions. Biol. Reprod. 2003, 68, 1208–1214. [Google Scholar] [CrossRef]
- Kadirvel, G.; Kathiravan, P.; Kumar, S. Protein tyrosine phosphorylation and zona binding ability of in vitro capacitated and cryopreserved buffalo spermatozoa. Theriogenology 2011, 75, 1630–1639. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, V.; Chhillar, S.; Atreja, S. Effect of supplementation of taurine or trehalose in extender on immunolocalization of tyrosine phosphoproteins in buffalo and cattle (karan fries) cryopreserved spermatozoa. Reprod. Domest. Anim. 2013, 48, 407–415. [Google Scholar] [CrossRef]
- Jankovičová, J.; Michalková, K.; Sečová, P.; Horovská, Ľ.; Maňásková-Postlerová, P.; Antalíková, J. Evaluation of protein phosphorylation in bull sperm during their maturation in the epididymis. Cell Tissue Res. 2018, 371, 365–373. [Google Scholar] [CrossRef]
- Bucci, D.; Galeati, G.; Tamanini, C.; Vallorani, C.; Rodriguez-Gil, J.E.; Spinaci, M. Effect of sex sorting on CTC staining, actin cytoskeleton and tyrosine phosphorylation in bull and boar spermatozoa. Theriogenology 2012, 77, 1206–1216. [Google Scholar] [CrossRef]
- Cormier, N.; Bailey, J.L. A Differential Mechanism Is Involved During Heparin- and Cryopreservation-Induced Capacitation of Bovine Spermatozoa. Biol. Reprod. 2003, 69, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Singh, V.; Yadav, H.P.; Verma, M.; Chauhan, D.S.; Saxena, A.; Yadav, S.; Swain, D.K. Effect of reduced glutathione supplementation in semen extender on tyrosine phosphorylation and apoptosis like changes in frozen thawed Hariana bull spermatozoa. Anim. Reprod. Sci. 2017, 182, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.M.; Minami, K.; Ogura, Y.; Otsuka, N.; Hama, S.; Harayama, H.; Sakase, M.; Fukushima, M. Variation among individual bulls in the distribution of acrosomal tyrosine-phosphorylated proteins in epididymal and ejaculated spermatozoa. Reprod. Fertil. Dev. 2017, 29, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Gadella, B.M.; Harrison, R.A.P. The capacitating agent bicarbonate induces protein kinase A-dependent changes in phospholipid transbilayer behavior in the sperm plasma membrane. Development 2000, 127, 2407–2420. [Google Scholar]
- Watson, P.F. Recent developments and concepts in the cryopreservation of spermatozoa and the assessment of their post-thawing function. Reprod. Fertil. Dev. 1995, 7, 871–891. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A.; Nixon, B. Are sperm capacitation and apoptosis the opposite ends of a continuum driven by oxidative stress? Asian J. Androl. 2015, 17, 633–639. [Google Scholar] [CrossRef]
- Leclerc, P.; De Lamirande, E.; Gagnon, C. Regulation of protein-tyrosine phosphorylation and human sperm capacitation by reactive oxygen derivatives. Free Radic. Biol. Med. 1997, 22, 643–656. [Google Scholar] [CrossRef]
- Yanagimachi, R. Mammalian Sperm Acrosome Reaction: Where Does It Begin Before Fertilization? Biol. Reprod. 2011, 85, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Yamashita, M.; Kimura, M.; Honda, A.; Kashiwabara, S. Sperm penetration through cumulus mass and zona pellucida. Int. J. Dev. Biol. 2008, 52, 677–682. [Google Scholar] [CrossRef]
- Hino, T.; Muro, Y.; Tamura-Nakano, M.; Okabe, M.; Tateno, H.; Yanagimachi, R. The behavior and acrosomal status of mouse spermatozoa in vitro, and within the oviduct during fertilization after natural mating. Biol. Reprod. 2016, 95, 50. [Google Scholar] [CrossRef]
- Grippo, A.A.; Way, A.L.; Killian, G.J. Effect of bovine ampullary and isthmic oviductal fluid on motility, acrosome reaction and fertility of bull spermatozoa. J. Reprod. Fertil. 1995, 105, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herz, Z.; Northey, D.; Lawyer, M.; First, M.L. Acrosome reaction of bovine spermatozoa in vivo: Sites and effects of stages of the estrous cycle. Biol. Reprod. 1985, 32, 1163–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deppe, M.; Morales, P.; Sánchez, R. Effect of protease inhibitors on the acrosome reaction and sperm-zona pellucida binding in bovine sperm. Reprod. Domest. Anim. 2008, 43, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Bukovsky, A.; Sharma, R.K.; Bansal, P.; Bhandari, B.; Gupta, S.K. In humans, zona pellucida glycoprotein-1 binds to spermatozoa and induces acrosomal exocytosis. Hum. Reprod. 2010, 25, 1643–1656. [Google Scholar] [CrossRef] [Green Version]
- Lishko, P.V.; Botchkina, I.L.; Kirichok, Y. Progesterone activates the principal Ca2+ channel of human sperm. Nature 2011, 471, 387–392. [Google Scholar] [CrossRef]
- Giojalas, L.C.; Rovasio, R.A.; Fabro, G.; Gakamsky, A.; Eisenbach, M. Timing of sperm capacitation appears to be programmed according to egg availability in the female genital tract. Fertil. Steril. 2004, 82, 247–249. [Google Scholar] [CrossRef]
- Whitfield, C.H.; Parkinson, T.J. Assessment of the fertilizing potential of frozen bovine spermatozoa by in vitro induction of acrosome reactions with calcium ionophore (A23187). Theriogenology 1995, 44, 413–422. [Google Scholar] [CrossRef]
- Mortimer, S.T. CASA—practical aspects. J. Androl. 2000, 21, 515–524. [Google Scholar]
- Lybaert, P.; Danguy, A.; Leleux, F.; Meuris, S.; Lebrun, P. Improved methodology for the detection and quantification of the acrosome reaction in mouse spermatozoa. Histol. Histopathol. 2009, 24, 999–1007. [Google Scholar]
- Fraser, L.R.; Abeydeera, L.R.; Niwa, K. Ca2+-Regulating mechanisms that modulate bull sperm capacitation and acrosomal exocytosis as determined by chlortetracycline analysis. Mol. Reprod. Dev. 1995, 40, 233–241. [Google Scholar] [CrossRef]
- Chamberland, A.; Fournier, V.; Tardif, S.; Sirard, M.A.; Sullivan, R.; Bailey, J.L. The effect of heparin on motility parameters and protein phosphorylation during bovine sperm capacitation. Theriogenology 2001, 55, 823–835. [Google Scholar] [CrossRef]
- Nagdas, S.K.; Smith, L.; Medina-Ortiz, I.; Hernandez-Encarnacion, L.; Raychoudhury, S. Identification of bovine sperm acrosomal proteins that interact with a 32-kDa acrosomal matrix protein. Mol. Cell. Biochem. 2016, 414, 153–169. [Google Scholar] [CrossRef] [Green Version]
- Botta, D.; de Arruda, R.P.; Watanabe, Y.F.; de Carvalho Balieiro, J.C.; Romanello, N.; do Nascimento Barreto, A.; de Andrade Pantoja, M.H.; Giro, A.; de Carvalho, C.P.T.; de Sousa Oliveira, A.; et al. Influence of post-thawing thermal environment on bovine sperm characteristics and in vitro fertility. Andrologia 2019, 51, 1–10. [Google Scholar] [CrossRef]
- Fraser, L.R. Minimum and maximum extracellular Ca2+ requirements during mouse sperm capacitation and fertilization in vitro. J. Reprod. Fertil. 1987, 81, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Pons-rejraji, H.; Bailey, J.L.; Leclerc, P. Cryopreservation affects bovine sperm intracellular parameters associated with capacitation and acrosome exocytosis. Reprod. Fertil. Dev. 2009, 21, 525–537. [Google Scholar] [CrossRef]
- Khalil, W.A.; El-Harairy, M.A.; Zeidan, A.E.B.; Hassan, M.A.E.; Mohey-Elsaeed, O. Evaluation of bull spermatozoa during and after cryopreservation: Structural and ultrastructural insights. Int. J. Vet. Sci. Med. 2018, 22, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Purdy, P.H.; Graham, J.K. Effect of Adding Cholesterol to Bull Sperm Membranes on Sperm Capacitation, the Acrosome Reaction, and Fertility. Biol. Reprod. 2004, 71, 522–527. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, Y.; Isono, A.; Kojima, A.; Arai, M.M.; Noda, T.; Sakase, M.; Fukushima, M.; Harayama, H. Distinct segment-specific functions of calyculin A-sensitive protein phosphatases in the regulation of cAMP-triggered events in ejaculated bull spermatozoa. Mol. Reprod. Dev. 2015, 82, 232–250. [Google Scholar] [CrossRef]
- Aitken, R.J.; Paterson, M.; Fisher, H.; Buckingham, D.W.; Van Duin, M. Redox regulation of tyrosine phosphorylation in human spermatozoa and its role in the control of human sperm function. J. Cell Sci. 1995, 108, 2017–2025. [Google Scholar]
- Kerns, K.; Sharif, M.; Zigo, M.; Xu, W.; Hamilton, L.E.; Sutovsky, M.; Ellersieck, M.; Drobnis, E.Z.; Bovin, N.; Oko, R.; et al. Sperm cohort-specific zinc signature acquisition and capacitation-induced zinc flux regulate sperm-oviduct and sperm-zona pellucida interactions. Int. J. Mol. Sci. 2020, 21, 2121. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Giaretta, E.; Munerato, M.; Yeste, M.; Galeati, G.; Spinaci, M.; Tamanini, C.; Mari, G.; Bucci, D. Implementing an open-access CASA software for the assessment of stallion sperm motility: Relationship with other sperm quality parameters. Anim. Reprod. Sci. 2017, 176, 11–19. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motility | Before DGC | T0h | T1h | T2h | T3h |
|---|---|---|---|---|---|
| VCL (µm/s) | 76 ± 9 | 89 ± 11 | 88 ± 3 | 78 ± 10 | 71 ± 12 |
| VAP (µm/s) | 37 ± 4 | 41 ± 4 | 42.9 ± 1 | 40 ± 5 | 38 ± 8 |
| VSL (µm/s) | 27 ± 3 | 32 ± 3 | 37.7 ± 1 | 35 ± 4 | 34 ± 7 |
| LIN (%) | 37 ± 3 a | 38 ± 3 ab | 42 ± 1 | 46 ± 1 bc | 48 ± 2 c |
| STR (%) | 73 ± 7 a | 79 ± 3 ab | 85 ± 1 abc | 88 ± 1 bc | 89 ± 1 c |
| WOB (%) | 48 ± 3 | 46 ± 3 | 48 ± 2 | 51 ± 1 | 53 ± 3 |
| BCF (Hz) | 19 ± 0.7 a | 20.4 ± 0.4 ab | 22.4 ± 0.8 | 22.9 ± 0.8 bc | 24.1 ± 1 c |
| ALH (µm) | 2.9 ± 0.3 | 3.3 ± 0.3 | 3.38 ± 0.07 | 3 ± 0.07 | 2.8 ± 0.5 |
| Motility (%) | 56 ± 11 ab | 63 ± 9 a | 55 ± 11 ab | 46 ± 13 b | 43 ± 10 b |
| Sample | Incubation (h) | Positive Acrosomal Exocytosis (%) | Intact Plasma Membrane (%) | PTP-Positive Sperm | ||
|---|---|---|---|---|---|---|
| Head (%) | Equatorial Region (%) | Whole Flagella (%) | ||||
| Before DGC | 4.2 ± 4.5 a | 58 ± 13 a | 40 ± 15 b | 31 ± 20 b | 1.4 ± 1.4 a | |
| Control | 0 | 3.5 ± 1 a | 59 ± 6 a | 49 ± 18 ab | 38 ± 23 b | 1.4 ± 1.6 a |
| Ionophore | 0 | 4.8 ± 2.5 ac | 57 ± 10 a | 50 ± 15 ab | 43 ± 20 ab | 0.8 ± 1 a |
| Control | 3 | 9.3 ± 2.7 c | 27 ± 16 c | 61 ± 21 ac | 54 ± 28 ac | 9 ± 9.8 c |
| Ionophore | 3 | 12.6 ± 4.6 c* | 29 ± 13 c | 59 ± 17 ac | 52 ± 25 ac | 11 ± 12 c |
| Incubation Time (h) | Ionophore Treatment | Acrosomal Exocytosis | Analyzed Spermatozoa (n) | PTP Patterns | ||
|---|---|---|---|---|---|---|
| I | II | III | ||||
| Before DGC | no | IA | 1356 | 64 ± 9 a | 24 ± 12 b | 12 ± 6 b |
| AE | 60 | 78 ± 23 a | 12 ± 20 b | 12 ± 16 b | ||
| 0 | no | IA | 1269 | 63 ± 7 a | 29 ± 8 b | 8 ± 2 c |
| AE | 44 | 65 ± 26 a | 30 ± 27 ab | 5 ± 9 b | ||
| 0 | yes | IA | 1410 | 56 ± 11 a**** | 36 ± 14 b*** | 8 ± 3 c** |
| AE | 71 | 90 ± 14 a | 10 ± 14 b | 0 | ||
| 3 | no | IA | 1222 | 54 ± 11 a* | 36 ± 16 a** | 10 ± 7 b* |
| AE | 130 | 82 ± 9 a | 15 ± 9 b | 3 ± 6 b | ||
| 3 | yes | IA | 1266 | 50 ± 13 a** | 39 ± 21 b* | 12 ± 8 c** |
| AE | 184 | 83 ± 8 a | 11 ± 11 b | 6 ± 5 b | ||
| Incubation Time (h) | Ionophore Treatment | Plasma Membrane Integrity | Analyzed Spermatozoa (n) | PTP Patterns | ||
|---|---|---|---|---|---|---|
| I | II | III | ||||
| Before DGC | no | I | 711 | 59 ± 26 a | 40 ± 22 a | 0.1 ± 0.4 b* |
| C | 510 | 51 ± 15 a | 47 ± 19 a | 1.4 ± 0.7 b | ||
| 0 | no | I | 688 | 46 ± 26 a | 43 ± 26 a* | 10 ± 22 b |
| C | 474 | 30 ± 17 b | 63 ± 27 a | 7 ± 10 c | ||
| 0 | yes | I | 625 | 53 ± 20 a** | 44 ± 32 a* | 3 ± 3 b |
| C | 451 | 28 ± 12 b | 68 ± 20 a | 4 ± 7 b | ||
| 3 | no | I | 318 | 35 ± 21 a* | 62 ± 20 a* | 2 ± 4 b |
| C | 838 | 16 ± 6 b | 83 ± 13 a | 1 ± 1 c | ||
| 3 | yes | I | 339 | 51 ± 21 a*** | 47 ± 21 a*** | 2 ± 2 b |
| C | 803 | 22 ± 16 b | 77 ± 16 a | 0.1 ± 0.4 c | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Díaz, S.; Grande-Pérez, S.; Arce-López, S.; Tamargo, C.; Olegario Hidalgo, C.; Pérez-Cerezales, S. Changes in the Cellular Distribution of Tyrosine Phosphorylation and Its Relationship with the Acrosomal Exocytosis and Plasma Membrane Integrity during In Vitro Capacitation of Frozen/Thawed Bull Spermatozoa. Int. J. Mol. Sci. 2020, 21, 2725. https://doi.org/10.3390/ijms21082725
Ruiz-Díaz S, Grande-Pérez S, Arce-López S, Tamargo C, Olegario Hidalgo C, Pérez-Cerezales S. Changes in the Cellular Distribution of Tyrosine Phosphorylation and Its Relationship with the Acrosomal Exocytosis and Plasma Membrane Integrity during In Vitro Capacitation of Frozen/Thawed Bull Spermatozoa. International Journal of Molecular Sciences. 2020; 21(8):2725. https://doi.org/10.3390/ijms21082725
Chicago/Turabian StyleRuiz-Díaz, Sara, Sergio Grande-Pérez, Sol Arce-López, Carolina Tamargo, Carlos Olegario Hidalgo, and Serafín Pérez-Cerezales. 2020. "Changes in the Cellular Distribution of Tyrosine Phosphorylation and Its Relationship with the Acrosomal Exocytosis and Plasma Membrane Integrity during In Vitro Capacitation of Frozen/Thawed Bull Spermatozoa" International Journal of Molecular Sciences 21, no. 8: 2725. https://doi.org/10.3390/ijms21082725
APA StyleRuiz-Díaz, S., Grande-Pérez, S., Arce-López, S., Tamargo, C., Olegario Hidalgo, C., & Pérez-Cerezales, S. (2020). Changes in the Cellular Distribution of Tyrosine Phosphorylation and Its Relationship with the Acrosomal Exocytosis and Plasma Membrane Integrity during In Vitro Capacitation of Frozen/Thawed Bull Spermatozoa. International Journal of Molecular Sciences, 21(8), 2725. https://doi.org/10.3390/ijms21082725