Sperm Cryodamage in Ruminants: Understanding the Molecular Changes Induced by the Cryopreservation Process to Optimize Sperm Quality

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
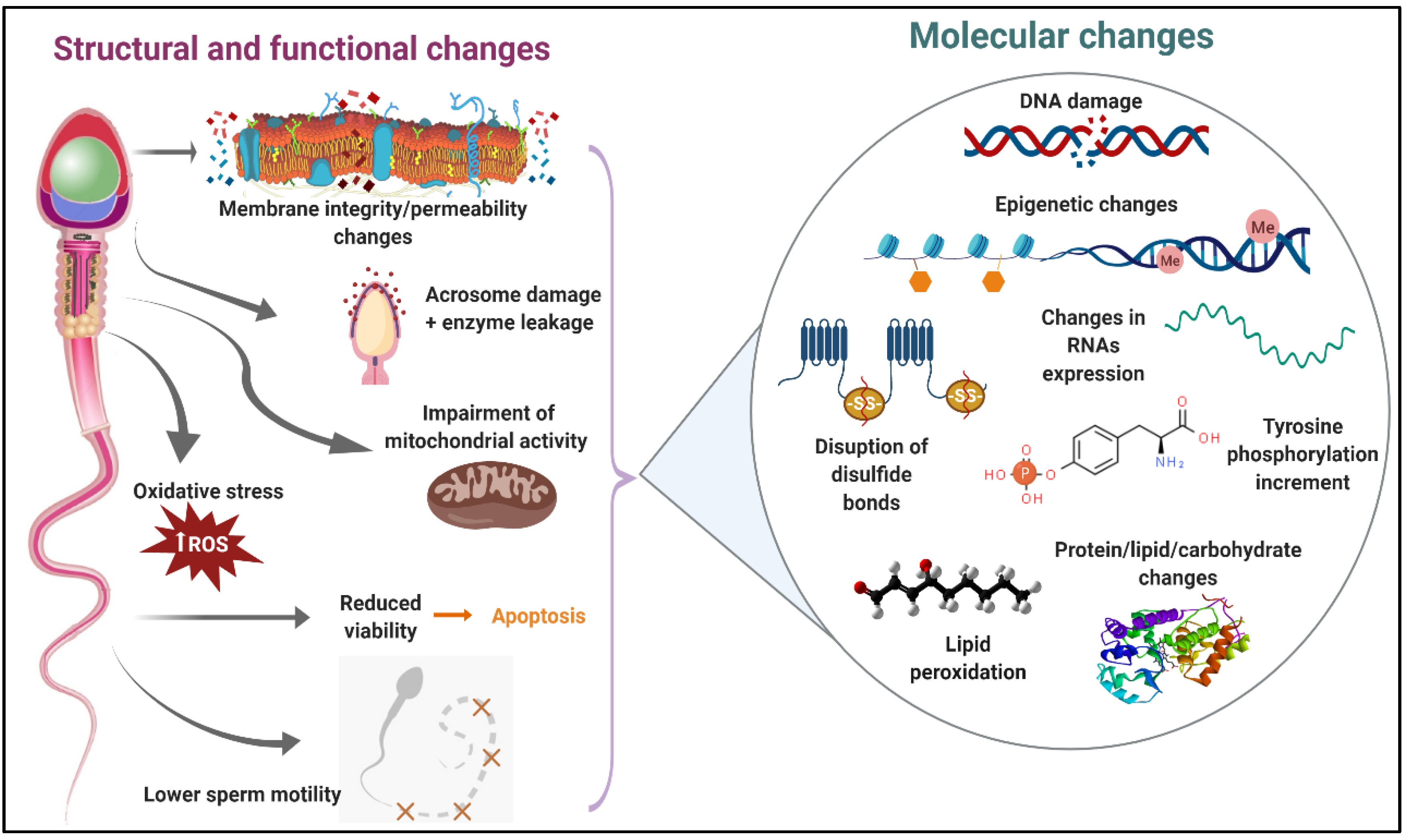
2. Unraveling How the Molecular Damage Caused by the Freezing–Thawing Process Affects Sperm Structure and Function
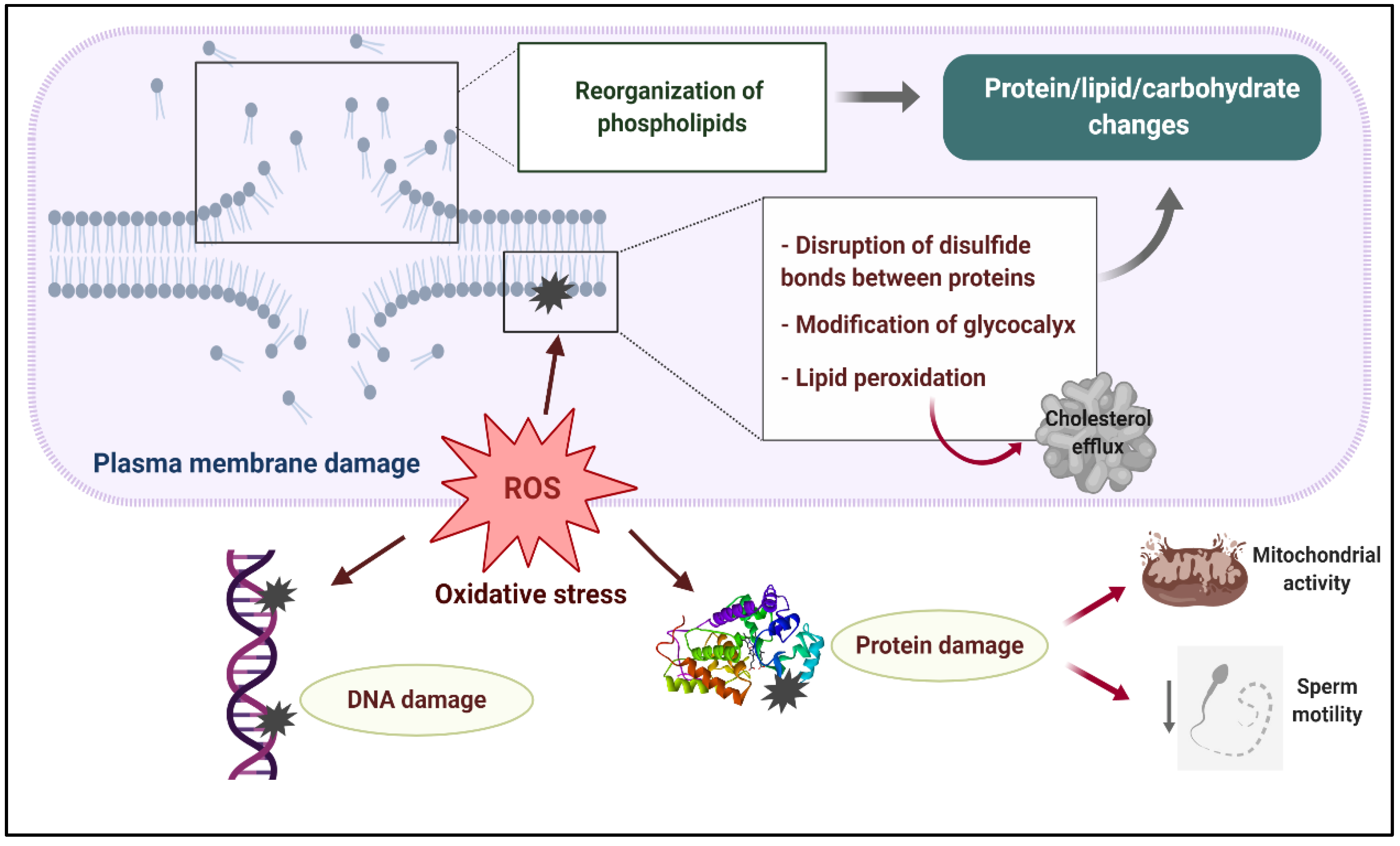
2.1. Molecular Changes in the Sperm Plasma Membrane during Cryopreservation
2.2. Molecular Disturbances in Sperm Energy Metabolism and Motility during Cryopreservation
2.3. Molecular Changes in the Sperm Chromatin during Cryopreservation
3. Molecular Mechanisms Involved in Those Factors That May Affect Sperm Cryotolerance
4. Molecular Aspects of Those Novel Strategies to Reduce Sperm Cryodamage
5. Concluding Remarks and Future Directions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TCP1 | T-complex protein 1 subunit alpha |
| LOC101123268 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase |
| RPN1 | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 1 |
| HEXB | Beta-hexosaminidase subunit beta-like isoform X1 |
| CSNK1G2 | Casein kinase I isoform gamma-2 isoform X2 |
| ICA | Inhibitor of carbonic anhydrase-like isoform X3 |
| LOC101123216 | Disintegrin and metalloproteinase domain-containing protein 20 |
| ADAM2 | Fertilin beta |
| TIMP-2 | Tissue inhibitor of metalloproteinases 2 |
| HSP70 | Heat shock 70 kDa protein |
| GLUT | Glucose transporter |
| CLU | Clusterin |
| BSP5 | Binder of sperm protein 5 |
| BSP1 | Binder of sperm protein 1 |
| aSFP | Acidic seminal fluid protein |
| HSP4AL | Heat shock 70 kDa protein 4 L isoform C1 |
| TRAP1 | Heat shock protein 75 kDa, mitochondrial isoform X3 |
| GPX4 | Phospholipid hydroperoxide glutathione peroxidase |
| GPX5 | Epididymal secretory glutathione peroxidase |
| CSNK2A2 | Casein kinase II subunit alpha |
| SOD2 | Superoxide dismutase 2 |
| PRDX5 | Peroxiredoxin 5 |
| COX5B | Cytochrome c oxidase subunit 5B, mitochondrial |
| AK1 | Adenylate kinase isoenzyme 1 |
| NUDFV2 | NADH dehydrogenase flavoprotein 2 |
| ODPB2 | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial isoform 2 |
| ACO2 | Aconitate hydratase, mitochondrial |
| NDPK7 | Nucleoside diphosphate kinase 7 |
| GPI | Glucose-6-phosphate isomerase |
| ALDOA | Fructose-bisphosphate aldolase |
| GAPDH5 | Glyceraldehyde-3-phosphate dehydrogenase, testis-specific |
| PGK2 | Phosphoglycerate kinase 2 |
| PGAM2 | Phosphoglycerate mutase 2 |
| PKM2 | Pyruvate kinase M2 |
| TPI | Triosephosphate isomerase |
| TEKT4 | Tektin 4 |
| ACTL7B | Actin-like protein 7B |
| HSP90 | Heat shock 90 kDa protein |
| DNMT3A | DNA (cytosine-5-)-methyltransferase 3 alpha |
| DNMT3B | DNA (cytosine-5-)-methyltransferase 3 beta |
| JHDM2A | JmjC domain-containing histone demethylation protein 2A |
| KAT8 | K(lysine) acetyltransferase 8 |
| IGF2 | Insulin-like growth factor 2 |
| ALB | Bovine serum albumin |
| SPZ1 | Spermadhesin 1 |
| PLA2G7 | Platelet-activating factor ace- tylhydrolase precursor |
| RSVP14 | Ram seminal vesicle protein 14 kDa |
| RSVP20 | Ram seminal vesicle protein 20 kDa |
| PRNT | Prion protein testis specific |
| CFTR | Cystic fibrosis transmembrane conductance regulator |
| EDIL3 | EGF-like repeat and discoidon 1-like domain-containing protein 3 |
| LEG1 | Liver enriched gene 1 |
| SPADH2 | Spermadhesin Z13-like |
| PPP1R7 | Protein phosphatase 1 regulatory subunit 7 trifunctional |
| BDH2 | Bodhesin-2 |
| RNASE9 | Inactive ribonuclease-like protein 9 |
| IZUMO4 | Izumo sperm–egg fusion protein 4 |
| PDB1 | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial precursor |
| HSP90A | Heat shock 90 kDa protein alpha |
References
- Bailey, J.; Morrier, A.; Cormier, N. Semen cryopreservation: Successes and persistent problems in farm species. Can. J. Anim. Sci. 2003, 83, 393–401. [Google Scholar] [CrossRef]
- Bailey, J.L.; Bilodeau, J.F.; Cormier, N. Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon. J. Androl. 2000, 21, 1–7. [Google Scholar]
- Kumar, A.; Prasad, J.K.; Srivastava, N.; Ghosh, S.K. Strategies to minimize various stress-related freeze–thaw damages during conventional cryopreservation of mammalian spermatozoa. Biopreserv. Biobank. 2019, 17, 603–612. [Google Scholar] [CrossRef]
- Ezzati, M.; Shanehbandi, D.; Hamdi, K.; Rahbar, S.; Pashaiasl, M. Influence of cryopreservation on structure and function of mammalian spermatozoa: An overview. Cell Tissue Bank. 2020, 21, 1–15. [Google Scholar] [CrossRef]
- Amidi, F.; Pazhohan, A.; Shabani Nashtaei, M.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Watson, P.F. Recent developments and concepts in the cryopreservation of spermatozoa and the assessment of their post-thawing function. Reprod. Fertil. Dev. 1995, 7, 871–891. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Wu, G.; Hong, Q.; Quan, G. Spermatozoa cryopreservation: State of art and future in small ruminants. Biopreserv. Biobank. 2018, 17, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Grötter, L.G.; Cattaneo, L.; Estela, P.; Kjelland, M.E.; Ferré, L.B. Recent advances in bovine sperm cryopreservation techniques with a focus on sperm post—Thaw quality optimization. Reprod. Domest. Anim. 2019, 54, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Lemma, A. Effect of Cryopreservation on sperm quality and fertility. Artif. Insemin. Farm Anim. 2011, 191–216. [Google Scholar]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef]
- Ugur, M.R.; Abdelrahman, A.S.; Evans, H.C.; Gilmore, A.A.; Hitit, M.; Arifiantini, R.I.; Purwantara, B.; Kaya, A.; Memili, E. Advances in cryopreservation of bull sperm. Front. Vet. Sci. 2019, 6, 268. [Google Scholar] [CrossRef] [Green Version]
- Pini, T.; Leahy, T.; de Graaf, S.P. Sublethal sperm freezing damage: Manifestations and solutions. Theriogenology 2018, 118, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Hezavehei, M.; Sharafi, M.; Kouchesfahani, H.M.; Henkel, R.; Agarwal, A.; Esmaeili, V.; Shahverdi, A. Sperm cryopreservation: A review on current molecular cryobiology and advanced approaches. Reprod. Biomed. Online 2018, 37, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Pulina, G.; Helena, A.; Francesconi, D.; Stefanon, B.; Calamari, L.; Lacetera, N.; Orto, V.D.; Pilla, F.; Marsan, A.; Mele, M.; et al. Sustainable ruminant production to help feed the planet. Ital. J. Anim. Sci. 2017, 16, 140–171. [Google Scholar] [CrossRef] [Green Version]
- Khalil, W.A.; El-harairy, M.A.; Zeidan, A.E.B.; Hassan, M.A.E.; Mohey-Elsaeed, O. Evaluation of bull spermatozoa during and after cryopreservation: Structural and ultrastructural insights. Int. J. Vet. Sci. Med. 2018, 6, S49–S56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gürler, H.; Malama, E.; Heppelmann, M.; Calisici, O.; Leiding, C.; Kastelic, J.P.; Bollwein, H. Effects of cryopreservation on sperm viability, synthesis of reactive oxygen species, and DNA damage of bovine sperm. Theriogenology 2016, 86, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Gagnon, C. Production of reactive oxygen species by spermatozoa undergoing cooling, freezing, and thawing. Mol. Reprod. Dev. 2001, 59, 451–458. [Google Scholar] [CrossRef]
- Pons-Rejraji, H.; Bailey, J.L.; Leclerc, P. Cryopreservation affects bovine sperm intracellular parameters associated with capacitation and acrosome exocytosis. Reprod. Fertil. Dev. 2009, 21, 525–537. [Google Scholar] [CrossRef]
- Yoon, S.J.; Kwon, W.S.; Rahman, M.S.; Lee, J.S.; Pang, M.G. A novel approach to identifying physical markers of cryo-damage in bull spermatozoa. PLoS ONE 2015, 10, e0126232. [Google Scholar] [CrossRef] [Green Version]
- Rasul, Z.; Ahmad, N.; Anzar, M. Changes in motion characteristics, plasma membrane integrity, and acrosome morphology during cryopreservation of buffalo spermatozoa. J. Androl. 2001, 22, 278–283. [Google Scholar]
- Kadirvel, G.; Kumar, S.; Kumaresan, A. Lipid peroxidation, mitochondrial membrane potential and DNA integrity of spermatozoa in relation to intracellular reactive oxygen species in liquid and frozen-thawed buffalo semen. Anim. Reprod. Sci. 2009, 114, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, P.; Singh, P.; Yadav, S.P.; Yadav, P.S. Assessment of sperm damages during different stages of cryopreservation in water buffalo by fluorescent probes. Cytotechnology 2016, 68, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Nasrullah, R.; Riaz, H.; Sattar, A.; Ahmad, N. Changes in motility, morphology, plasma membrane and acrosome integrity during stages of cryopreservation of buck sperm. J. S. Afr. Vet. Assoc. 2014, 85, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Dorado, J.; Hidalgo, M.; Muñoz, A.; Rodríguez, I. Assessment of goat semen freezability according to the spermatozoa characteristics from fresh and frozen samples. Anim. Reprod. Sci. 2009, 112, 150–157. [Google Scholar] [CrossRef]
- Chauhan, M.S.; Kapila, R.; Gandhi, K.K.; Anand, S.R. Acrosome damage and enzyme leakage of goat spermatozoa during dilution, cooling and freezing. Andrologia 1994, 26, 21–26. [Google Scholar] [CrossRef]
- Nur, Z.; Zik, B.; Ustuner, B.; Sagirkaya, H.; Ozguden, C.G. Effects of different cryoprotective agents on ram sperm morphology and DNA integrity. Theriogenology 2010, 73, 1267–1275. [Google Scholar] [CrossRef]
- Peris-Frau, P.; Martín-Maestro, A.; Iniesta-Cuerda, M.; Sánchez-Ajofrín, I.; Cesari, A.; Garde, J.J.; Villar, M.; Soler, A.J. Cryopreservation of ram sperm alters the dynamic changes associated with in vitro capacitation. Theriogenology 2020, 145, 100–108. [Google Scholar] [CrossRef]
- Da Silva Maia, M.; Bicudo, S.D.; Sicherle, C.C.; Rodello, L.; Gallego, I.C.S. Lipid peroxidation and generation of hydrogen peroxide in frozen-thawed ram semen cryopreserved in extenders with antioxidants. Anim. Reprod. Sci. 2010, 122, 118–123. [Google Scholar] [CrossRef]
- Peris, S.I.; Morrier, A.; Dufour, M.; Bailey, J.L. Cryopreservation of ram semen facilitates sperm DNA damage: Relationship between sperm andrological parameters and the sperm chromatin structure Assay. J. Androl. 2004, 25, 224–233. [Google Scholar] [CrossRef]
- García-Álvarez, O.; Maroto-Morales, A.; Martínez-Pastor, F.; Garde, J.J.; Ramón, M.; Fernández-Santos, M.R.; Esteso, M.C.; Pérez-Guzmán, M.D.; Soler, A.J. Sperm characteristics and in vitro fertilization ability of thawed spermatozoa from black manchega ram: Electroejaculation and postmortem collection. Theriogenology 2009, 72, 160–168. [Google Scholar] [CrossRef]
- Fernández-Santos, M.R.; Esteso, M.C.; Montoro, V.; Soler, A.J.; Garde, J.J. Cryopreservation of Iberian red deer (Cervus elaphus hispanicus) epididymal spermatozoa: Effects of egg yolk, glycerol and cooling rate. Theriogenology 2006, 66, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Gur, Y.; Breitbart, H. Mammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomes. Genes Dev. 2006, 20, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, M.A.; Nixon, B.; Naumovski, N.; Aitken, R.J. Proteomic insights into the maturation and capacitation of mammalian spermatozoa. Syst. Biol. Reprod. Med. 2012, 58, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; De Mateo, S.; Estanyol, J.M. Sperm cell proteomics. Proteomics 2009, 9, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, G.; Swain, N.; Samanta, L. Sperm proteome: What is on the horizon? Reprod. Sci. 2014, 22, 638–653. [Google Scholar] [CrossRef]
- Dai, D.; Qazi, I.H.; Ran, M.; Liang, K.; Zhang, Y. Exploration of miRNA and mRNA profiles in fresh and frozen-thawed boar sperm by transcriptome and small RNA sequencing. Int. J. Mol. Sci. 2019, 20, 802. [Google Scholar] [CrossRef] [Green Version]
- Darin-Bennett, A.; White, I.G. Influence of the cholesterol content of mammalian spermatozoa on susceptibility to cold-shock. Cryobiology 1977, 14, 466–470. [Google Scholar] [CrossRef]
- Wojtusik, J.; Wang, Y.; Pukazhenthi, B.S. Pretreatment with cholesterol-loaded cyclodextrins prevents loss of motility associated proteins during cryopreservation of addra gazelle (Nanger dama ruficollis) spermatozoa. Cryobiology 2018, 81, 74–80. [Google Scholar] [CrossRef]
- Lessard, C.; Parent, S.; Leclerc, P.; Bailey, J.L.; Sullivan, R. Cryopreservation alters the levels of the bull sperm surface protein P25b. J. Androl. 2000, 21, 700–707. [Google Scholar]
- Pini, T.; Rickard, J.P.; Leahy, T.; Crossett, B.; Druart, X.; De Graaf, S.P. Cryopreservation and egg yolk medium alter the proteome of ram spermatozoa. J. Proteom. 2018, 181, 73–82. [Google Scholar] [CrossRef]
- Peris-Frau, P.; Martín-Maestro, A.; Iniesta-Cuerda, M.; Sánchez-Ajofrín, I.; Mateos-Hernández, L.; Garde, J.J.; Villar, M.; Soler, A.J. Freezing—Thawing procedures remodel the proteome of ram sperm before and after in vitro capacitation. Int. J. Mol. Sci. 2019, 20, 4596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, T.; Divyashree, B.C.; Roy, S.C.; Roy, K.S. Loss of heat shock protein 70 from apical region of buffalo (Bubalus bubalis) sperm head after freezing and thawing. Theriogenology 2016, 85, 828–834. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, K.; Zhao, X.; Zhang, Y.; Ma, Y.; Hu, J. Differential proteome association study of freeze-thaw damage in ram sperm. Cryobiology 2016, 72, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Westfalewicz, B.; Dietrich, M.A.; Ciereszko, A. Impact of cryopreservation on bull (Bos taurus) semen proteome. J. Anim. Sci. 2015, 93, 5240–5253. [Google Scholar] [CrossRef]
- Marti, E.; Marti, J.I.; Muiño-Blanco, T.; Cebrián-Pérez, J.A. Effect of the cryopreservation process on the activity and immunolocalization of antioxidant enzymes in ram spermatozoa. J. Androl. 2008, 29, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Bilodeau, J.-F.; Chatterjee, S.; Sirard, M.A.; Gagnon, C. Levels of antioxidant defenses are decreased in bovine spermatozoa after a cycle of freezing. Mol. Hum. Reprod. 2000, 55, 282–288. [Google Scholar] [CrossRef]
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Ryu, D.Y.; Park, Y.J.; Pang, M.G. Proteomic identification of cryostress in epididymal spermatozoa. J. Anim. Sci. Biotechnol. 2016, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Park, Y.J.; Pang, M.G. Addition of cryoprotectant significantly alters the epididymal sperm proteome. PLoS ONE 2016, 11, e0152690. [Google Scholar] [CrossRef] [Green Version]
- Martin-Hidalgo, D.; Bragado, M.J.; Batista, A.R.; Oliveira, P.F.; Alves, M.G. Antioxidants and male fertility: From molecular studies to clinical evidence. Antioxidants 2019, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Chatterjee, S.; De Lamirande, E.; Gagnon, C. Cryopreservation alters membrane sulfhydryl status of bull spermatozoa: Protection by oxidized glutathione. Mol. Reprod. Dev. 2001, 60, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Free radicals, lipid peroxidation and sperm function. Reprod. Fertil. Dev. 1995, 7, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Baker, M.A.; Nixon, B. Are sperm capacitation and apoptosis the opposite ends of a continuum driven by oxidative stress? Asian J. Androl. 2015, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mostek, A.; Dietrich, M.A.; Słowińska, M.; Ciereszko, A. Cryopreservation of bull semen is associated with carbonylation of sperm proteins. Theriogenology 2017, 92, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Losano, J.; Angrimani, D.; Dalmazzo, A.; Rui, B.; Brito, M.; Mendes, C.; Kawai, G.; Vannucchi, C.; Assumpção, M.; Barnabe, V.; et al. Effect of mitochondrial uncoupling and glycolysis inhibition on ram sperm functionality. Reprod. Domest. Anim. 2017, 52, 289–297. [Google Scholar] [CrossRef]
- Naresh, S. Effect of cooling and cryopreservation on cytoskeleton actin and protein tyrosine phosphorylation in buffalo spermatozoa. Cryobiology 2016, 72, 7–13. [Google Scholar] [CrossRef]
- Felipe-Pérez, Y.E.; Valencia, J.; Juárez-Mosqueda, M. de L.; Pescador, N.; Roa-Espitia, A.L.; Hernández-González, E.O. Cytoskeletal proteins F-actin and β-dystrobrevin are altered by the cryopreservation process in bull sperm. Cryobiology 2012, 64, 103–109. [Google Scholar] [CrossRef]
- Zhang, X.G.; Hu, S.; Han, C.; Zhu, Q.C.; Yan, G.J.; Hu, J.H. Association of heat shock protein 90 with motility of post-thawed sperm in bulls. Cryobiology 2015, 70, 164–169. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, K.; Jain, S.; Hassan, T.; Dada, R. Novel insights into the genetic and epigenetic paternal contribution to the human embryo. Clinics 2013, 68, 5–14. [Google Scholar] [CrossRef]
- Lewis, S.E.M.; Aitken, R.J. DNA damage to spermatozoa has impacts on fertilization and pregnancy. Cell Tissue Res. 2005, 322, 33–41. [Google Scholar] [CrossRef]
- Khalifa, T.A.A.; Rekkas, C.A.; Lymberopoulos, A.G.; Sioga, A.; Dimitriadis, I.; Papanikolaou, T. Factors affecting chromatin stability of bovine spermatozoa. Anim. Reprod. Sci. 2008, 104, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Kadirvel, G.; Periasamy, S.; Kumar, S. Effect of cryopreservation on apoptotic-like events and its relationship with cryocapacitation of buffalo (Bubalus bubalis) sperm. Reprod. Domest. Anim. 2012, 47, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.; Sabido, O.; Durand, P.; Levy, R. Cryopreservation induces an apoptosis-like mechanism in bull sperm. Biol. Reprod. 2004, 71, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Schans, G.P.; Haring, R.; van Dijk-Knijnenburg, H.C.M.; Bruijnzeel, P.L.B.; Den Daas, N.H.G. An immunochemical assay to detect DNA damage in bovine sperm. J. Androl. 2000, 21, 250–257. [Google Scholar]
- Peris, S.I.; Bilodeau, J.-F.; Dufour, M.; Bailey, J.L. Impact of cryopreservation and reactive oxygen species on DNA integrity, lipid peroxidation, and functional parameters in ram sperm. Mol. Reprod. Dev. 2007, 74, 878–892. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P. The sperm chromatin structure assay (SCSA®) and other sperm DNA fragmentation tests for evaluation of sperm nuclear DNA integrity as related to fertility. Anim. Reprod. Sci. 2016, 169, 56–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosálvez, J.; López-Fernández, C.; Fernández, J.L.; Gouraud, A.; Holt, W.V. Relationships between the dynamics of iatrogenic DNA damage and genomic design in mammalian spermatozoa from eleven species. Mol. Reprod. Dev. 2011, 78, 951–961. [Google Scholar]
- Cortés-Gutiérrez, E.I.; López-Fernández, C.; Fernández, J.L.; Dávila-Rodríguez, M.I.; Johnston, S.D.; Gosálvez, J. Interpreting sperm DNA damage in a diverse range of mammalian sperm by means of the two-tailed comet assay. Front. Genet. 2014, 5, 1–11. [Google Scholar]
- Domínguez-Rebolledo, Á.E.; Fernández-Santos, M.R.; García-Álvarez, O.; Maroto-Morales, A.; Garde, J.J.; Martínez-Pastor, F. Washing increases the susceptibility to exogenous oxidative stress in red deer spermatozoa. Theriogenology 2009, 72, 1073–1084. [Google Scholar] [CrossRef]
- Martinez-Pastor, F.; Johannisson, A.; Gil, J.; Kaabi, M.; Anel, L.; Paz, P.; Rodriguez-Martinez, H. Use of chromatin stability assay, mitochondrial stain JC-1, and fluorometric assessment of plasma membrane to evaluate frozen-thawed ram semen. Anim. Reprod. Sci. 2004, 84, 121–133. [Google Scholar] [CrossRef]
- Peris-Frau, P.; Álvarez-Rodríguez, M.; Martín-Maestro, A.; Iniesta-Cuerda, M.; Sánchez-Ajofrín, I.; Garde, J.J.; Rodriguez-Martinez, H.; Soler, A.J. Comparative evaluation of DNA integrity using sperm chromatin structure assay and Sperm-Ovis-Halomax during in vitro capacitation of cryopreserved ram spermatozoa. Reprod. Domest. Anim. 2019, 54, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Shaman, J.A.; Ward, W.S. Non-genetic contributions of the sperm nucleus to embryonic development. Asian J. Androl. 2011, 13, 31–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Rajput, S.; De, S.; Kumar, R.; Chakravarty, A.K.; Datta, T.K. Genome-wide pro fi ling of sperm DNA methylation in relation to buffalo (Bubalus bubalis) bull fertility. Theriogenology 2014, 82, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Lin, S.; Zhao, Z.; Sun, Q. Epigenetic dynamics and interplay during spermatogenesis and embryogenesis: Implications for male fertility and offspring health. Oncotarget 2017, 8, 53804–53818. [Google Scholar] [CrossRef] [Green Version]
- Collins, L.J.; Schönfeld, B.; Xiaowei, S.C. The epigenetics of non-coding RNA. In Handbook of Epigenetics; Elsevier Inc.: Amsterdam, The Netherlands, 2011; pp. 49–61. [Google Scholar]
- Chen, X.; Wang, Y.; Zhu, H.; Hao, H.; Zhao, X.; Qin, T.; Wang, D. Comparative transcript profiling of gene expression of fresh and frozen-thawed bull sperm. Theriogenology 2015, 83, 504–511. [Google Scholar] [CrossRef]
- Aurich, C.; Schreiner, B.; Ille, N.; Alvarenga, M.; Scarlet, D. Cytosine methylation of sperm DNA in horse semen after cryopreservation. Theriogenology 2016, 86, 1347–1352. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Peng, W.; Ding, L.; He, L.; Zhang, Y.; Fang, D.; Tang, K. A preliminary study on epigenetic changes during boar spermatozoa cryopreservation. Cryobiology 2014, 69, 119–127. [Google Scholar] [CrossRef]
- Ortiz-Rodriguez, J.M.; Ortega-Ferrusola, C.; Gil, M.C.; Martín-Cano, F.E.; Gaitskell-Phillips, G.; Rodríguez-Martínez, H.; Hinrichs, K.; Álvarez-Barrientos, A.; Román, Á.; Peña, F.J. Transcriptome analysis reveals that fertilization with cryopreserved sperm downregulates genes relevant for early embryo development in the horse. PLoS ONE 2019, 14, e0213420. [Google Scholar] [CrossRef] [Green Version]
- Perumal, P.; Savino, N.; Ctr, S.; Mh, K.; Ezung, E.; Chang, S.; Tzt, S. Seasonal effect on physiological, reproductive and fertility profiles in breeding mithun bulls. Asian Pac. J. Reprod. 2017, 6, 268–278. [Google Scholar] [CrossRef]
- De Rensis, F.; John, R. Heat stress and seasonal effects on reproduction in the dairy cow—A review. Theriogenology 2003, 60, 1139–1151. [Google Scholar] [CrossRef]
- Al-Dawood, A. Towards heat stress management in small Ruminants—A review. Ann. Anim. Sci. 2017, 17, 59–88. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Moreno, J.; Gómez-Brunet, A.; Toledano-Díaz, A.; Picazo, R.; Gonzalez-Bulnes, A.; López-Sebastián, A. Seasonal endocrine changes and breeding activity in mediterranean wild ruminants. Reprod. Domest. Anim. 2006, 41, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fresneda, L.; O’Brien, E.; Velázquez, R.; Toledano-Díaz, A.; Martínez-Cáceres, C.M.; Tesfaye, D.; Schellander, K.; García-Vázquez, F.A.; Santiago-Moreno, J. Seasonal variation in sperm freezability associated with changes in testicular germinal epithelium in domestic (Ovis aries) and wild (Ovis musimon) sheep. Reprod. Fertil. Dev. 2019, 31, 1545–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argov-Argaman, N.; Mahgrefthe, K.; Zeron, Y.; Roth, Z. Season-induced variation in lipid composition is associated with semen quality in Holstein bulls. Reproduction 2013, 145, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westfalewicz, B.; Dietrich, M.; Słowińska, M.; Judycka, S.; Ciereszko, A. Seasonal changes in the proteome of cryopreserved bull semen supernatant. Theriogenology 2019, 126, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Muiño-Blanco, T.; Pérez-Pé, R.; Cebrián-Pérez, J.A. Seminal plasma proteins and sperm resistance to stress. Reprod. Domest. Anim. 2008, 43, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, M.P.; Falcinelli, A.; Hozbor, F.; Sánchez, E.; Cesari, A.; Alberio, R.H. Seasonal variations in the composition of ram seminal plasma and its effect on frozen-thawed ram sperm. Theriogenology 2008, 69, 564–573. [Google Scholar] [CrossRef]
- Leahy, T.; Marti, J.I.; Evans, G.; Maxwell, W.M.C. Seasonal variation in the protective effect of seminal plasma on frozen-thawed ram spermatozoa. Anim. Reprod. Sci. 2010, 119, 147–153. [Google Scholar] [CrossRef]
- Morató, R.; Prieto-Martínez, N.; Muiño, R.; Hidalgo, C.O.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaporin 11 is related to cryotolerance and fertilising ability of frozen-thawed bull spermatozoa. Reprod. Fertil. Dev. 2018, 30, 1099–1108. [Google Scholar] [CrossRef]
- Ryu, D.Y.; Song, W.H.; Pang, W.K.; Yoon, S.J.; Rahman, M.S.; Pang, M.G. Freezability biomarkers in bull epididymal spermatozoa. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Wang, Y.F.; Wang, H.; Wang, C.W.; Zan, L.S.; Hu, J.H.; Li, Q.W.; Jia, Y.H.; Ma, G.J. HSP90 expression correlation with the freezing resistance of bull sperm. Zygote 2014, 22, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Rickard, J.P.; Leahy, T.; Soleilhavoup, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Lynch, G.W.; Druart, X.; de Graaf, S.P. The identification of proteomic markers of sperm freezing resilience in ram seminal plasma. J. Proteom. 2015, 126, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Rego, J.P.A.; Martins, J.M.; Wolf, C.A.; Van Tilburg, M.; Moreno, F.; Monteiro-Moreira, A.C.; Moreira, R.A.; Santos, D.O.; Moura, A.A. Proteomic analysis of seminal plasma and sperm cells and their associations with semen freezability in Guzerat bulls. J. Anim. Sci. 2016, 94, 5308–5320. [Google Scholar] [CrossRef] [PubMed]
- Song, W.H.; Ryu, D.Y.; Pang, W.K.; Yoon, S.J.; Rahman, M.S.; Pang, M.G. NT5C1B and FH are closely associated with cryoprotectant tolerance in spermatozoa. Andrology 2019, 8, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, M.; Gajare, V.C.; Mandal, A.; Mondal, M.; Das, S.K.; Ghosh, M.K.; Rai, S.; Behera, R. Electrophoretic profile of seminal proteins and their correlation with in vitro sperm characters in Black Bengal buck semen. Vet. World 2019, 12, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Rickard, J.P.; Schmidt, R.E.; Maddison, J.W.; Bathgate, R.; Lynch, G.W.; Druart, X.; De Graaf, S.P. Variation in seminal plasma alters the ability of ram spermatozoa to survive cryopreservation. Reprod. Fertil. Dev. 2016, 28, 516–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soler, A.; García, A.; Fernández-Santos, M.; Esteso, M.; Garde, J. Effects of thawing procedure on postthawed in vitro viability and in vivo fertility of Red Deer epididymal. J. Androl. 2003, 24, 746–756. [Google Scholar] [CrossRef]
- Fujii, T.; Hirayama, H.; Fukuda, S.; Kageyama, S.; Naito, A.; Yoshino, H.; Moriyasu, S.; Yamazaki, T.; Sakamoto, K.; Hayakawa, H.; et al. Expression and localization of aquaporins 3 and 7 in bull spermatozoa and their relevance to sperm motility after cryopreservation. J. Reprod. Dev. 2018, 64, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Martínez, N.; Morató, R.; Muiño, R.; Hidalgo, C.O.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaglyceroporins 3 and 7 in bull spermatozoa: Identification, localisation and their relationship with sperm cryotolerance. Reprod. Fertil. Dev. 2017, 29, 1249–1259. [Google Scholar] [CrossRef]
- Jobim, M.I.M.; Oberst, E.R.; Salbego, C.; Souza, D.; Wald, V.B.; Tramontina, F.; Mattos, R.C. Two-dimensional polyacrylamide gel electrophoresis of bovine seminal plasma proteins and their relation with semen freezability. Theriogenology 2004, 61, 255–266. [Google Scholar] [CrossRef]
- Pereira, R.M.; Mesquita, P.; Pires, V.M.R.; Baptista, M.C.; Barbas, J.P.; Pimenta, J.; Horta, A.E.M.; Prates, J.A.M.; Marques, C.C. Prion protein testis specific (PRNT) gene polymorphisms and transcript level in ovine spermatozoa: Implications in freezability, fertilization and embryo production. Theriogenology 2018, 115, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, S.; Hering, D.M.; Kordan, W.; Lecewicz, M. Missense mutation within cystic fibrosis transmembrane conductance regulator (CFTR) gene is associated with selected parameters of the frozen-thawed sperm in Holstein-Friesian bulls. Pol. J. Vet. Sci. 2019, 22, 221–225. [Google Scholar] [PubMed]
- Nikbin, S.; Panandam, J.M.; Yaakub, H.; Murugaiyah, M.; Sazili, A.Q. Novel SNPs in heat shock protein 70 gene and their association with sperm quality traits of Boer goats and Boer crosses. Anim. Reprod. Sci. 2014, 146, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Petrunkina, A.M.; Waberski, D.; Günzel-Apel, A.R.; Töpfer-Petersen, E. Determinants of sperm quality and fertility in domestic species. Reproduction 2007, 134, 3–17. [Google Scholar] [CrossRef]
- Maroto-Morales, A.; Ramón, M.; García-Álvarez, O.; Soler, A.J.; Fernández-Santos, M.R.; Roldan, E.R.S.; Gomendio, M.; Pérez-Guzmán, M.D.; Garde, J.J. Morphometrically-distinct sperm subpopulations defined by a multistep statistical procedure in Ram ejaculates: Intra—And interindividual variation. Theriogenology 2012, 77, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Esteso, M.C.; Fernández-Santos, M.R.; Soler, A.J.; Montoro, V.; Quintero-Moreno, A.; Garde, J.J. The effects of cryopreservation on the morphometric dimensions of Iberian red deer (Cervus elaphus hispanicus) epididymal sperm heads. Reprod. Domest. Anim. 2006, 41, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Muiño, R.; Peña, A.I.; Rodríguez, A.; Tamargo, C.; Hidalgo, C.O. Effects of cryopreservation on the motile sperm subpopulations in semen from Asturiana de los Valles bulls. Theriogenology 2009, 72, 860–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbas, J.P.; Leahy, T.; Horta, A.E.; García-Herreros, M. Sperm kinematics and subpopulational responses during the cryopreservation process in caprine ejaculates. Cryobiology 2018, 82, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Ramón, M.; Pérez-Guzmán, M.D.; Jiménez-Rabadán, P.; Esteso, M.C.; García-Álvarez, O.; Maroto-Morales, A.; Anel-López, L.; Soler, A.J.; Fernández-Santos, M.R.; Garde, J.J. Sperm cell population dynamics in ram semen during the cryopreservation process. PLoS ONE 2013, 8, e59189. [Google Scholar] [CrossRef]
- D’Amours, O.; Frenette, G.; Bourassa, S.; Calvo, É.; Blondin, P.; Sullivan, R. Proteomic markers of functional sperm population in bovines: Comparison of low- and high-density spermatozoa following cryopreservation. J. Proteome Res. 2018, 17, 177–188. [Google Scholar] [CrossRef]
- Ortega-Ferrusola, C.; Gil, M.C.; Rodríguez-Martínez, H.; Anel, L.; Peña, F.J.; Martín-Muñoz, P. Flow cytometry in spermatology: A bright future ahead. Reprod. Domest. Anim. 2017, 52, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fresneda, L.; Castaño, C.; Bóveda, P.; Tesfaye, D.; Schellander, K.; Santiago-Moreno, J.; García-Vázquez, F.A. Epididymal and ejaculated sperm differ on their response to the cryopreservation and capacitation processes in mouflon (Ovis musimon). Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Quinn, P.J.; White, I.G. Phospholipid and cholesterol content of epididymal and ejaculated ram spermatozoa and seminal plasma in relation to cold shock. Aust. J. Biol. Sci. 1967, 20, 1205–1215. [Google Scholar] [CrossRef] [Green Version]
- Pini, T.; Leahy, T.; Soleilhavoup, C.; Tsikis, G.; Labas, V.; Combes-Soia, L.; Harichaux, G.; Rickard, J.P.; Druart, X.; De Graaf, S.P. Proteomic investigation of ram spermatozoa and the proteins conferred by seminal plasma. J. Proteome Res. 2016, 15, 3700–3711. [Google Scholar] [CrossRef] [PubMed]
- Perez-Patiño, C.; Barranco, I.; Li, J.; Padilla, L.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J.; Parrilla, I. Cryopreservation differentially alters the proteome of epididymal and ejaculated pig spermatozoa. Int. J. Mol. Sci. 2019, 20, 1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardini, A.; Hozbor, F.; Sanchez, E.; Fornés, M.W.; Alberio, R.H.; Cesari, A. Conserved ram seminal plasma proteins bind to the sperm membrane and repair cryopreservation damage. Theriogenology 2011, 76, 436–447. [Google Scholar] [CrossRef]
- Leahy, T.; Rickard, J.P.; Bernecic, N.C.; Druart, X.; De Graaf, S. Ram seminal plasma and its functional proteomic assessment. Reproduction 2019, 157, 243–256. [Google Scholar] [CrossRef]
- Manjunath, P.; Bergeron, A.; Lefebvre, J.; Fan, J. Seminal plasma proteins: Functions and interaction with protective agents during semen preservation. Soc. Reprod. Fertil. Suppl. 2007, 65, 217–228. [Google Scholar]
- Pini, T.; Farmer, K.; Druart, X.; Teixeira-gomes, A.P.; Tsikis, G.; Labas, V.; Leahy, T.; Graaf, S.P. De Binder of Sperm Proteins protect ram spermatozoa from freeze-thaw damage. Cryobiology 2018, 82, 78–87. [Google Scholar] [CrossRef]
- Layek, S.S.; Mohanty, T.K.; Kumaresan, A.; Parks, J.E. Cryopreservation of bull semen: Evolution from egg yolk based to soybean based extenders. Anim. Reprod. Sci. 2016, 172, 1–9. [Google Scholar] [CrossRef]
- Barbas, J.P.; Mascarenhas, R.D. Cryopreservation of domestic animal sperm cells. Cell Tissue Bank 2009, 10, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Allai, L.; Benmoula, A.; Marciane da Silva, M.; Nasser, B.; El Amiri, B. Supplementation of ram semen extender to improve seminal quality and fertility rate. Anim. Reprod. Sci. 2018, 192, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Purdy, P.H. A review on goat sperm cryopreservation. Small Rumin. Res. 2006, 63, 215–225. [Google Scholar] [CrossRef]
- Mata-Campuzano, M.; Álvarez-Rodríguez, M.; Álvarez, M.; Tamayo-Canul, J.; Anel, L.; De Paz, P.; Martínez-Pastor, F. Post-thawing quality and incubation resilience of cryopreserved ram spermatozoa are affected by antioxidant supplementation and choice of extender. Theriogenology 2015, 83, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Santos, M.R.; Martínez-Pastor, F.; García-Macías, V.; Esteso, M.C.; Soler, A.J.; de Paz, P.; Anel, L.; Garde, J.J. Extender osmolality and sugar supplementation exert a complex effect on the cryopreservation of Iberian red deer (Cervus elaphus hispanicus) epididymal spermatozoa. Theriogenology 2007, 67, 738–753. [Google Scholar] [CrossRef]
- Iqbal, S.; Naz, S.; Ahmed, H.; Andrabi, S.M.H. Cryoprotectant effect of trehalose in extender on post-thaw quality and in vivo fertility of water buffalo (Bubalus bubalis) bull spermatozoa. Andrologia 2018, 50, 1–6. [Google Scholar] [CrossRef]
- Hu, J.H.; Zan, L.S.; Zhao, X.L.; Li, Q.W.; Jiang, Z.L.; Li, Y.K.; Li, X. Effects of trehalose supplementation on semen quality and oxidative stress variables in frozen-thawed bovine semen. J. Anim. Sci. 2010, 88, 1657–1662. [Google Scholar] [CrossRef]
- Aboagla, E.M.E.; Terada, T. Trehalose-enhanced fluidity of the goat sperm membrane and its protection during freezing. Biol. Reprod. 2003, 69, 1245–1250. [Google Scholar] [CrossRef]
- Rostami, B.; Ebrahimi, D.; Sadeghipanah, H.; Masoumi, R.; Shahir, M.H. Effects of supplementation of tris-egg yolk extender with different sugars and antioxidants on freezability of ram semen. Cryobiology 2019. [Google Scholar] [CrossRef]
- Fujikawa, T.; Imamura, S.; Tokumaru, M.; Ando, T.; Gen, Y.; Hyon, S.H.; Kubota, C. Cryoprotective effect of antifreeze polyamino-acid (Carboxylated Poly-L-Lysine) on bovine sperm: A technical note. Cryobiology 2018, 82, 159–162. [Google Scholar] [CrossRef]
- Tariq, A.; Ahmad, M.; Iqbal, S.; Riaz, M.I.; Tahir, M.Z.; Ghafoor, A.; Riaz, A. Effect of carboxylated poly L-Lysine as a cryoprotectant on post-thaw quality and in vivo fertility of Nili Ravi buffalo (Bubalus bubalis) bull semen. Theriogenology 2020, 144, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Longobardi, V.; Albero, G.; De Canditiis, C.; Salzano, A.; Natale, A.; Balestrieri, A.; Neglia, G.; Campanile, G.; Gasparrini, B. Cholesterol-loaded cyclodextrins prevent cryocapacitation damages in buffalo (Bubalus bubalis) cryopreserved sperm. Theriogenology 2017, 89, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Mocé, E.; Purdy, P.H.; Graham, J.K. Treating ram sperm with cholesterol-loaded cyclodextrins improves cryosurvival. Anim. Reprod. Sci. 2010, 118, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Purdy, P.H.; Graham, J.K. Effect of cholesterol-loaded cyclodextrin on the cryosurvival of bull sperm. Cryobiology 2004, 48, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Salmon, V.M.; Leclerc, P.; Bailey, J.L.; Animales, S.; Al, S.E.T. Cholesterol-loaded cyclodextrin increases the cholesterol content of goat sperm to improve cold and osmotic resistance and maintain sperm function after cryopreservation. Biol. Reprod. 2016, 94, 85. [Google Scholar] [CrossRef]
- Batissaco, L.; De Arruda, R.P.; Alves, M.B.R.; Torres, M.A.; Lemes, K.M.; Prado-Filho, R.R.; De Almeida, T.G.; De Andrade, A.F.C.; Celeghini, E.C.C. Cholesterol-loaded cyclodextrin is efficient in preserving sperm quality of cryopreserved ram semen with low freezability. Reprod. Biol. 2020, 20, 14–24. [Google Scholar] [CrossRef]
- Fernández-Santos, M.R.; Martínez-Pastor, F.; García-Macías, V.; Esteso, M.C.; Soler, A.J.; Paz, P.; Anel, L.; Garde, J.J. Sperm characteristics and DNA integrity of Iberian red deer (Cervus elaphus hispanicus) epididymal spermatozoa frozen in the presence of enzymatic and nonenzymatic antioxidants. J. Androl. 2007, 28, 294–305. [Google Scholar] [CrossRef]
- Sánchez-Rubio, F.; Fernández-Santos, M.R.; Castro-Vázquez, L.; García-Álvarez, O.; Maroto-Morales, A.; Soler, A.J.; Martínez-Pastor, F.; Garde, J.J. Cinnamtannin B-1, a novel antioxidant for sperm in red deer. Anim. Reprod. Sci. 2018, 195, 44–52. [Google Scholar] [CrossRef]
- Khalil, W.A.; El-Harairy, M.A.; Zeidan, A.E.B.; Hassan, M.A.E. Impact of selenium nano-particles in semen extender on bull sperm quality after cryopreservation. Theriogenology 2019, 126, 121–127. [Google Scholar] [CrossRef]
- Hozyen, H.F.; Shamy, A.A.E.; Farghali, A.A. In vitro Supplementation of nano selenium minimizes freeze-thaw induced damage to ram spermatozoa. Int. J. Vet. Sci. 2019, 8, 249–254. [Google Scholar]
- Shahin, M.A.; Khalil, W.A.; Saadeldin, I.M.; Swelum, A.A.-A.; El-harairy, M.A. Comparison between the effects of adding vitamins, trace elements, and nanoparticles to SHOTOR extender on the cryopreservation of dromedary camel epididymal spermatozoa. Animals 2020, 10, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashtestani, F.; Ghourchian, H.; Najafi, A. Silver-gold-apoferritin nanozyme for suppressing oxidative stress during cryopreservation. Mater. Sci. Eng. C 2019, 94, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Cebrián-Pérez, J.A.; González-Arto, M.; Dos Santos Hamilton, T.R.; Pérez-Pé, R.; Muiño-Blanco, T. Melatonin in sperm biology: Breaking paradigms. Reprod. Domest. Anim. 2014, 49, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, I.; Kohram, H.; Ardabili, F.F. Antioxidative effects of melatonin on kinetics, microscopic and oxidative parameters of cryopreserved bull spermatozoa. Anim. Reprod. Sci. 2013, 139, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhao, C.; Xiang, H.; Zhao, X.; Zhong, R. Melatonin inhibits formation of mitochondrial permeability transition pores and improves oxidative phosphorylation of frozen-thawed ram sperm. Front. Endocrinol. 2020, 10, 896. [Google Scholar] [CrossRef] [Green Version]
- Ferrusola, C.O.; Fernández, L.G.; Sandoval, C.S.; García, B.M.; Martínez, H.R.; Tapia, J.A.; Peña, F.J. Inhibition of the mitochondrial permeability transition pore reduces “apoptosis like” changes during cryopreservation of stallion spermatozoa. Theriogenology 2010, 74, 458–465. [Google Scholar] [CrossRef]
- Pillai, H.; Parmar, M.S.; Shende, A.M.; Thomas, J.; Kartha, H.S.; Sharma, T.; Ghosh, G.S.K.; Kumaresan, A.; Bhure, S.K. Effect of supplementation of recombinant regucalcin in extender on cryopreservation of spermatozoa of water buffalo (Bubalus bubalis). Mol. Reprod. Dev. 2017, 84, 1133–1139. [Google Scholar] [CrossRef]
- Ledesma, A.; Zalazar, L.; Buchelly, F.; Ignacio, J.; Brown, P.; Mitch, E.; Hozbor, F.; Cesari, A. Recombinant peptide reverses cryo-capacitation in ram sperm and improves in vitro fertilization. Anim. Reprod. Sci. 2019, 207, 61–72. [Google Scholar] [CrossRef]
- Zalazar, L.; Iniesta-Cuerda, M.; Sánchez-Ajofrín, I.; Soler, A.J.; Garde, J.J.; Cesari, A. Recombinant SPINK3 improves ram sperm quality and in vitro fertility after cryopreservation. Theriogenology 2020, 144, 45–55. [Google Scholar] [CrossRef]
- Ramírez-Vasquez, R.; Cesari, A.; Hozbor, F.; Greco, M.B.; Cano, A. Extenders modify the seminal plasma ability to minimize freeze-thaw damage on ram sperm. Reprod. Domest. Anim. 2019, 54, 1621–1629. [Google Scholar] [CrossRef]
- Robles, V.; Valcarce, D.G.; Riesco, M.F. The use of antifreeze proteins in the cryopreservation of gametes and embryos. Biomolecules 2019, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, S.R.; Oliver, J.E.; Upreti, G.C. Effect of antifreeze proteins on the motility of ram spermatozoa. Cryobiology 1994, 31, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Prathalingam, N.S.; Holt, W.V.; Revell, S.G.; Mirczuk, S.; Fleck, R.A.; Watson, P.F. Impact of antifreeze proteins and antifreeze glycoproteins on bovine sperm during freeze-thaw. Theriogenology 2006, 66, 1894–1900. [Google Scholar] [CrossRef]
- Qadeer, S.; Khan, M.A.; Ansari, M.S.; Rakha, B.A.; Ejaz, R.; Husna, A.U.; Ashiq, M.; Iqbal, R.; Ullah, N.; Akhter, S. Evaluation of antifreeze protein III for cryopreservation of Nili-Ravi (Bubalus bubalis) buffalo bull sperm. Anim. Reprod. Sci. 2014, 148, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Qadeer, S.; Khan, M.A.; Ansari, M.S.; Rakha, B.A.; Ejaz, R.; Iqbal, R.; Younis, M.; Ullah, N.; DeVries, A.L.; Akhter, S. Efficiency of antifreeze glycoproteins for cryopreservation of Nili-Ravi (Bubalus bubalis) buffalo bull sperm. Anim. Reprod. Sci. 2015, 157, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Qadeer, S.; Khan, M.A.; Shahzad, Q.; Azam, A.; Ansari, M.S.; Rakha, B.A.; Ejaz, R.; Husna, A.U.; Duman, J.G.; Akhter, S. Efficiency of beetle (Dendroides canadensis) recombinant antifreeze protein for buffalo semen freezability and fertility. Theriogenology 2016, 86, 1662–1669. [Google Scholar] [CrossRef] [PubMed]
- Arando, A.; Gonzalez, A.; Delgado, J.V.; Arrebola, F.A.; Perez-Marín, C.C. Storage temperature and sucrose concentrations affect ram sperm quality after vitrification. Anim. Reprod. Sci. 2017, 181, 175–185. [Google Scholar] [CrossRef]
- Jiménez-Rabadán, P.; García-Álvarez, O.; Vidal, A.; Maroto-Morales, A.; Iniesta-Cuerda, M.; Ramón, M.; del Olmo, E.; Fernández-Santos, R.; Garde, J.J.; Soler, A.J. Effects of vitrification on ram spermatozoa using free-egg yolk extenders. Cryobiology 2015, 71, 85–90. [Google Scholar] [CrossRef]
- Pradiee, J.; Esteso, M.C.; Castaño, C.; Toledano-Díaz, A.; Lopez-Sebastián, A.; Guerra, R.; Santiago-Moreno, J. Conventional slow freezing cryopreserves mouflon spermatozoa better than vitrification. Andrologia 2017, 49, 1–7. [Google Scholar] [CrossRef]
- Pradieé, J.; Sánchez-Calabuig, M.J.; Castaño, C.; O’Brien, E.; Esteso, M.C.; Beltrán-Breña, P.; Maillo, V.; Santiago-Moreno, J.; Rizos, D. Fertilizing capacity of vitrified epididymal sperm from Iberian ibex (Capra pyrenaica). Theriogenology 2018, 108, 314–320. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Protein Name | Species | Origin/Source | Function during Cryopreservation | References |
|---|---|---|---|---|
| Seminal plasma protein PDC-109 (BSP-A1/A2) | Bull | Seminal plasma | Sperm membrane protection. | [101] |
| Acidic seminal fluid protein (aSFP) | Bull | Seminal plasma | Sperm membrane protection against lipid peroxidation. | [101] |
| Clusterin (CLU) | Bull | Seminal plasma | Sperm membrane protection. | [101] |
| Heat-shock protein 90 (HSP90) | Bull | Ejaculated sperm | Sperm motility regulation and protection against oxidative stress, thermal stress and apoptosis. | [92] |
| Ram | Seminal plasma | [93] | ||
| 26S proteasome non-ATPase regulatory subunit 2 (PSMD2) | Ram | Seminal plasma | Optimal cell organization by removing misfolded or damaged proteins. | [93] |
| Tripeptidyl-peptidase 2 (TPP2) | Ram | Seminal plasma | Protection from cryo-capacitation, increasing sperm longevity and fertility. | [93] |
| Transitional endoplasmic reticulum ATPase (VCP) | Ram | Seminal plasma | Positive regulation of mitochondrial membrane potential. | [93] |
| Sorbitol dehydrogenase (SORD) | Ram | Seminal plasma | Sperm motility regulation. | [93] |
| Chaperonin-containing t-complex polypeptide 1 (CCT) | Ram | Seminal plasma | Sperm membrane stabilization through an efficient protein folding. | [93] |
| Acrosome formation-associated factor isoform 2 (AFAF) | Bull | Ejaculated sperm | Preservation of acrosome integrity and viability. | [94] |
| Disintegrin and metalloproteinase domain-containing protein 2 (ADAM2) | Bull | Ejaculated sperm | Sperm membrane stabilization. | [94] |
| Aquaporin 3 (AQP3) | Bull | Ejaculated sperm | Protection against osmotic changes by controlling efficiently the flux of water and glycerol in the sperm membrane. Also involved in sperm motility. | [99,100] |
| Aquaporin 7 (AQP7) | Bull | Ejaculated sperm | Protection against osmotic changes by controlling efficiently the flux of water and glycerol in the sperm membrane. Also involved in sperm motility. | [99] |
| Aquaporin 11 (AQP11) | Bull | Ejaculated sperm | Protection against osmotic changes by controlling efficiently the flux of water and glycerol in the sperm membrane. | [90] |
| ATP synthase subunit beta, mitochondrial (ATP1B1) | Bull | Epididymal sperm | ATP synthesis through the electron transport chain, which explains the higher mitochondrial activity and motility of GF. | [91] |
| Fumarate hydratase, mitochondrial (FH) | Bull | Ejaculated sperm | Involved in energy metabolism, which explains the higher motility of GF. | [95] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peris-Frau, P.; Soler, A.J.; Iniesta-Cuerda, M.; Martín-Maestro, A.; Sánchez-Ajofrín, I.; Medina-Chávez, D.A.; Fernández-Santos, M.R.; García-Álvarez, O.; Maroto-Morales, A.; Montoro, V.; et al. Sperm Cryodamage in Ruminants: Understanding the Molecular Changes Induced by the Cryopreservation Process to Optimize Sperm Quality. Int. J. Mol. Sci. 2020, 21, 2781. https://doi.org/10.3390/ijms21082781
Peris-Frau P, Soler AJ, Iniesta-Cuerda M, Martín-Maestro A, Sánchez-Ajofrín I, Medina-Chávez DA, Fernández-Santos MR, García-Álvarez O, Maroto-Morales A, Montoro V, et al. Sperm Cryodamage in Ruminants: Understanding the Molecular Changes Induced by the Cryopreservation Process to Optimize Sperm Quality. International Journal of Molecular Sciences. 2020; 21(8):2781. https://doi.org/10.3390/ijms21082781
Chicago/Turabian StylePeris-Frau, Patricia, Ana Josefa Soler, María Iniesta-Cuerda, Alicia Martín-Maestro, Irene Sánchez-Ajofrín, Daniela Alejandra Medina-Chávez, María Rocío Fernández-Santos, Olga García-Álvarez, Alejandro Maroto-Morales, Vidal Montoro, and et al. 2020. "Sperm Cryodamage in Ruminants: Understanding the Molecular Changes Induced by the Cryopreservation Process to Optimize Sperm Quality" International Journal of Molecular Sciences 21, no. 8: 2781. https://doi.org/10.3390/ijms21082781
APA StylePeris-Frau, P., Soler, A. J., Iniesta-Cuerda, M., Martín-Maestro, A., Sánchez-Ajofrín, I., Medina-Chávez, D. A., Fernández-Santos, M. R., García-Álvarez, O., Maroto-Morales, A., Montoro, V., & Garde, J. J. (2020). Sperm Cryodamage in Ruminants: Understanding the Molecular Changes Induced by the Cryopreservation Process to Optimize Sperm Quality. International Journal of Molecular Sciences, 21(8), 2781. https://doi.org/10.3390/ijms21082781