The Expression Profile of Dental Pulp-Derived Stromal Cells Supports Their Limited Capacity to Differentiate into Adipogenic Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
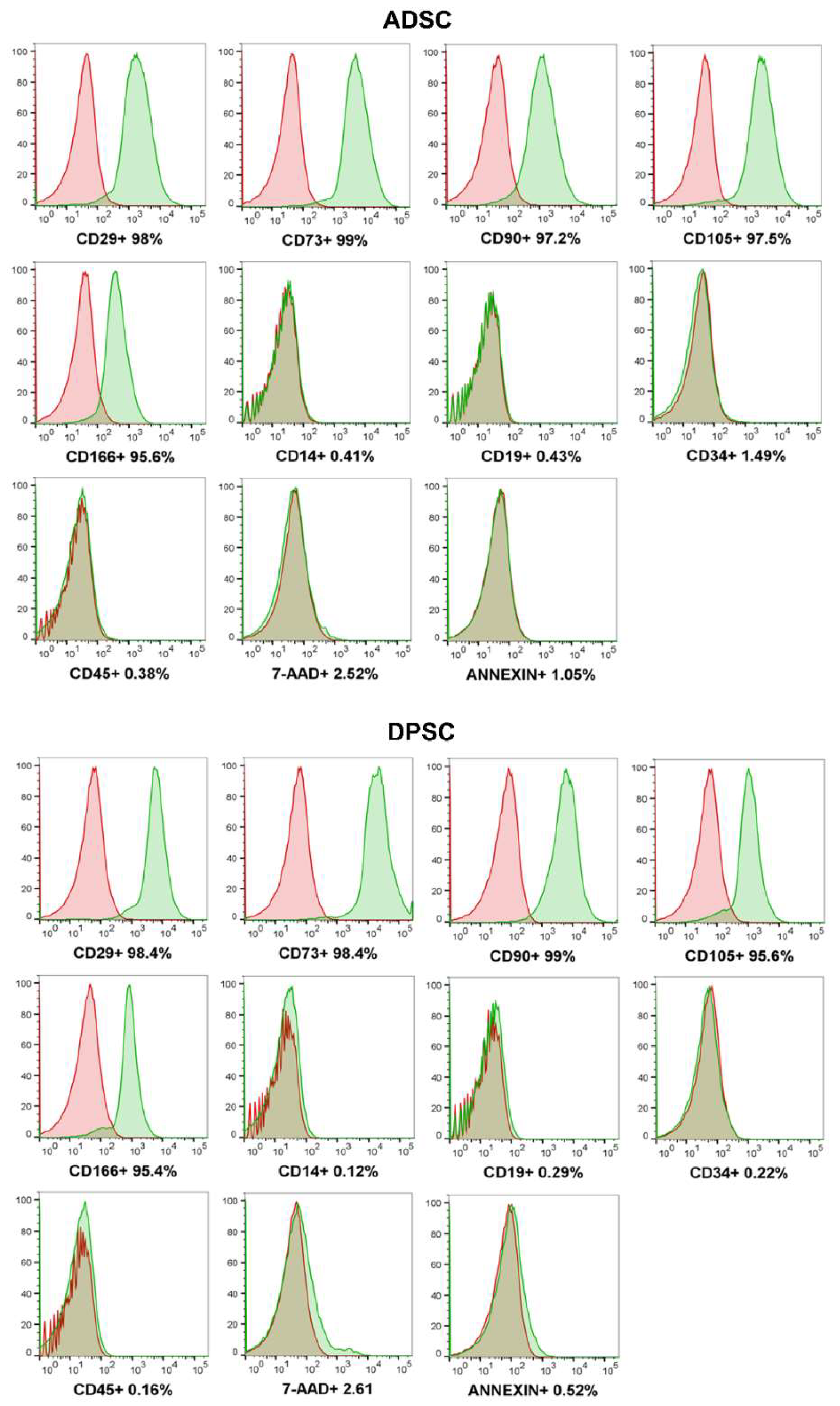
2.1. DPSCs Are Immunophenotypically and Morphologically Similar to ADSCs and at Least Bipotent
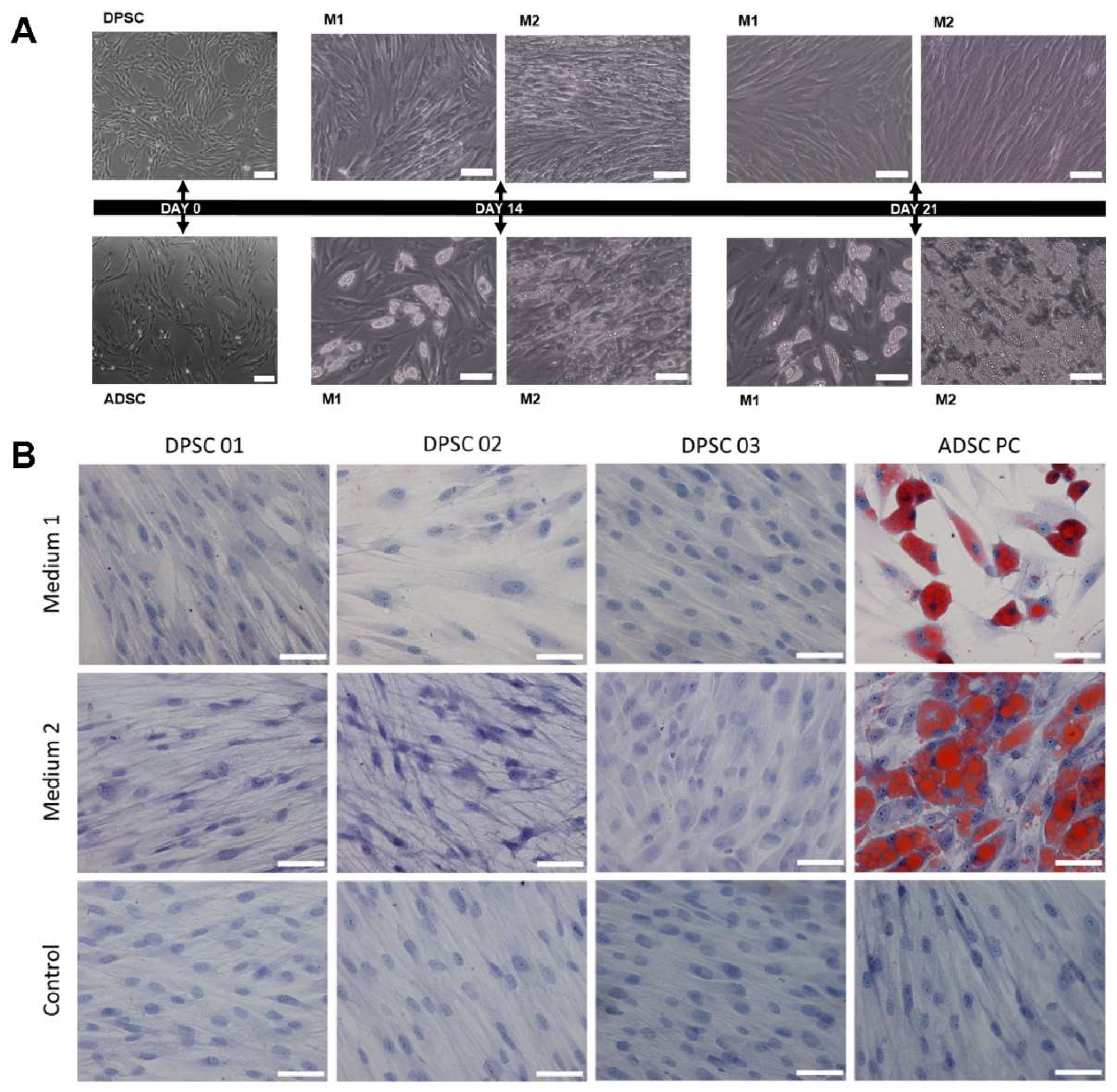
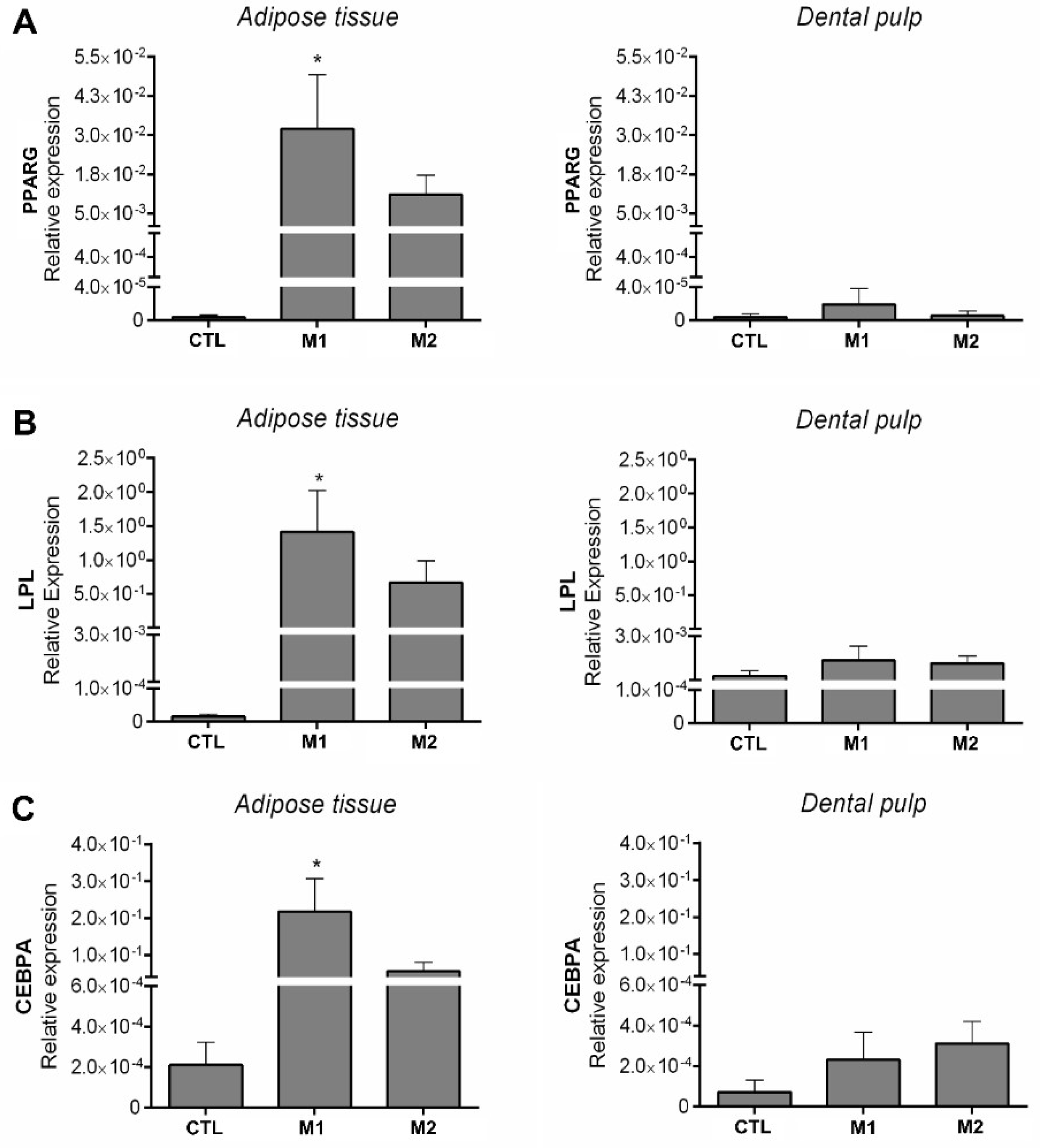
2.2. DPSCs do not Differentiate into Adipocytes After 21 Days of Induction Using Two Different Protocols
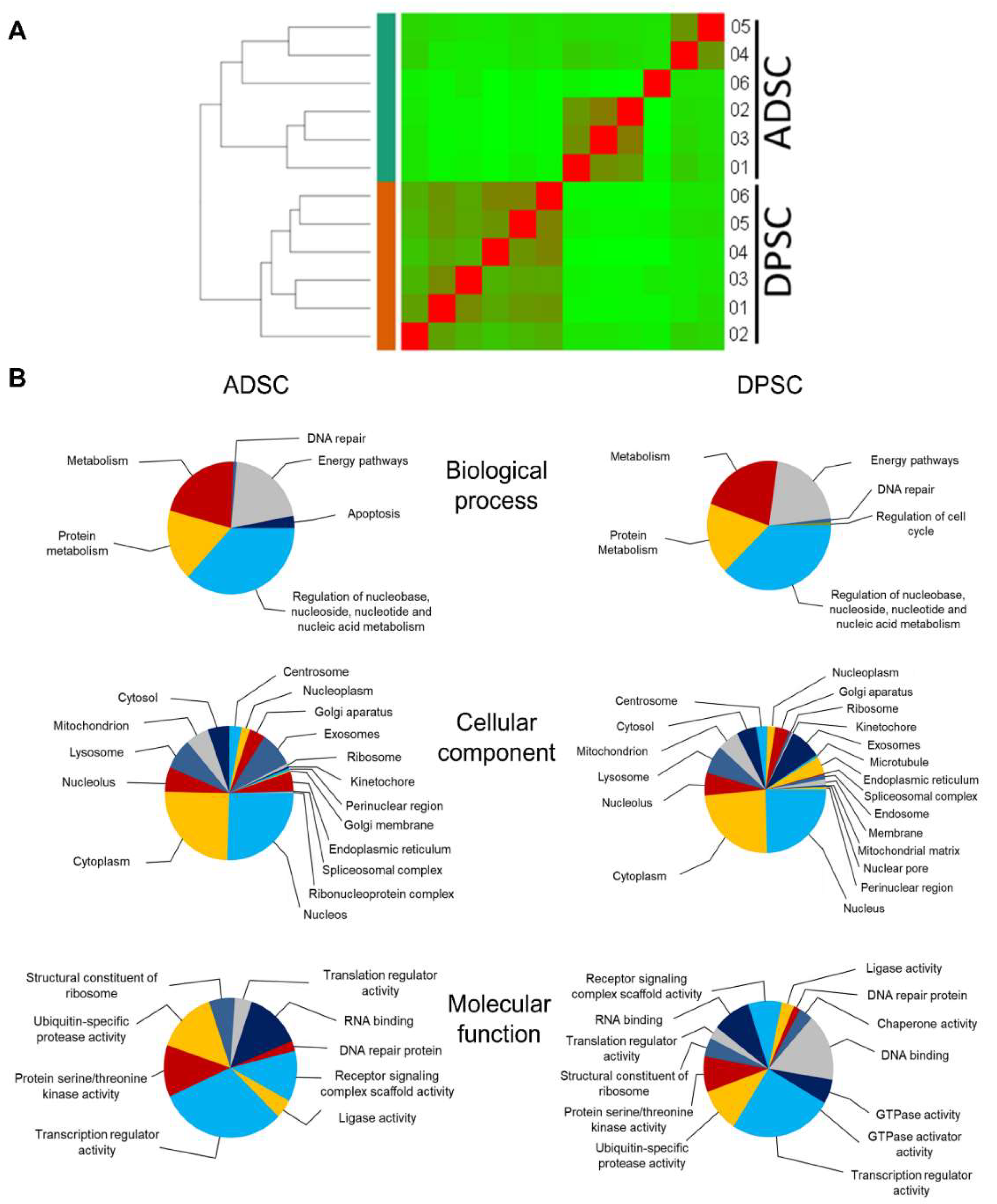
2.3. Transcriptomic Analyses Revealed Basal Differences between ADSCs and DPSCs and the Inability of DPSCs to Undergo Adipogenic Differentiation
3. Materials and Methods
3.1. Isolation and Cell Cultivation
3.2. Immunophenotypic Characterisation
3.3. Osteogenic Differentiation
3.4. Chondrogenic Differentiation
3.5. Adipogenic Cell Differentiation
3.6. Total RNA ExStraction and Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
3.7. Total RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
3.8. RNA Sequencing
3.9. Data Availability
3.10. Bioinformatics Analysis of RNA-Seq Data
3.11. Gene Ontology Annotation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal Stromal Cells |
| ADSCs | Adipose Tissue-Derived Stromal Cells |
| DPSCs | Dental Pulp-Derived Stromal Cells |
| PPARG | Peroxisome Proliferator-Activated Receptor Gamma |
| LPL | Lipoprotein Lipase |
| BMP | Bone Morphogenetic Protein |
| ISCT | International Society for Cellular Therapy |
| FABP4 | Fatty Acid Binding Protein 4 |
| M1 | Commercial Culture Medium |
| M2 | Custom Culture Medium |
| CEBPA | CCAAT/Enhancer-Binding Protein Alpha |
| PC | Positive Control |
| CTL | Control |
| GAPDH | Glyceraldehyde-3-phosphate Dehydrogenase |
| SEM | Standard Error of the Mean |
| P | Passage |
| RNA | Ribonucleic Acid |
| mRNA | Messenger RNA |
| DNA | Deoxyribonucleic Acid |
| GO | Gene Ontology |
| DEGs | Differentially Expressed Genes |
| FDR | False Discovery Rate |
| DKK1 | Dickkopf WNT Signalling Pathway Inhibitor 1 |
| sFRP4 | Frizzled-Related Protein 4 |
| TCF4 | Transcription Factor 4 |
| WNT10B | Wingless-Type, Member 10B |
| TGF-β | Transforming Growth Factor Beta |
| NOG | Noggin |
| ID | DNA-binding |
| MSX2 | Msh-Like 2 Homeobox |
| RUNX2 | Runt-Related Transcription Factor 2 |
| PBS | Phosphate-Buffered Saline Solution |
| IMDM | Iscove’s Modified Dulbecco’s Media |
| FBS | Foetal Bovine Serum |
| DMEM-F12 | Dulbecco’s Modified Eagle Medium: Nutrient Mixture F-12 |
| 7-AAD | 7-Aminoactinomycin D |
| hMSC | Human Mesenchymal Stromal Cells |
| RT-PCR | Reverse Transcription-Polymerase Chain Reaction |
| cDNA | Complementary DNA |
| PCR | Polymerase Chain Reaction |
| qRT-PCR | Quantitative Real-Time Polymerase Chain Reaction |
References
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Caplan, A.I. Why are MSCs therapeutic? New data: New insight. J. Pathol. 2009, 217, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytom. Part A 2018, 93, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Ide, R.; Saiki, C.; Kumazawa, Y.; Okamura, H. Human Dental Pulp Cells Differentiate toward Neuronal Cells and Promote Neuroregeneration in Adult Organotypic Hippocampal Slices In Vitro. Int. J. Mol. Sci. 2017, 18, 1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.; Mazor, Y.; Rinon, A.; Blumenthal, J.; Golan, Y.; Buzhor, E.; Livnat, I.; Ben-Ari, S.; Lieder, I.; Shitrit, A.; et al. LifeMap Discovery: The embryonic development, stem cells, and regenerative medicine research portal. PLoS ONE 2013, 8, e66629. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [Green Version]
- Tamaki, Y.; Nakahara, T.; Ishikawa, H.; Sato, S. In vitro analysis of mesenchymal stem cells derived from human teeth and bone marrow. Odontology 2013, 101, 121–132. [Google Scholar] [CrossRef]
- Isobe, Y.; Koyama, N.; Nakao, K.; Osawa, K.; Ikeno, M.; Yamanaka, S.; Okubo, Y.; Fujimura, K.; Bessho, K. Comparison of human mesenchymal stem cells derived from bone marrow, synovial fluid, adult dental pulp, and exfoliated deciduous tooth pulp. Int. J. Oral Maxillofac. Surg. 2016, 45, 124–131. [Google Scholar] [CrossRef]
- Alsulaimani, R.S.; Ajlan, S.A.; Aldahmash, A.M.; Alnabaheen, M.S.; Ashri, N.Y. Isolation of dental pulp stem cells from a single donor and characterization of their ability to differentiate after 2 years of cryopreservation. Saudi Med. J. 2016, 37, 551–560. [Google Scholar] [CrossRef]
- Ullah, I.; Subbarao, R.B.; Kim, E.J.; Bharti, D.; Jang, S.J.; Park, J.S.; Shivakumar, S.B.; Lee, S.L.; Kang, D.; Byun, J.H.; et al. In vitro comparative analysis of human dental stem cells from a single donor and its neuronal differentiation potential evaluated by electrophysiology. Life Sci. 2016, 154, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Berry, D.C.; Stenesen, D.; Zeve, D.; Graff, J.M. The developmental origins of adipose tissue. Development 2013, 140, 3939–3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From stem cell to adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christodoulides, C.; Lagathu, C.; Sethi, J.K.; Vidal-Puig, A. Adipogenesis and WNT signalling. Trends Endocrinol. Metab. TEM 2009, 20, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Yin, C.; Zhao, F.; Ali, A.; Ma, J.; Qian, A. Mesenchymal Stem Cells: Cell Fate Decision to Osteoblast or Adipocyte and Application in Osteoporosis Treatment. Int. J. Mol. Sci. 2018, 19, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, N. Characterisation of dental pulp stem cells: A new horizon for tissue regeneration? Arch. Oral Biol. 2012, 57, 1439–1458. [Google Scholar] [CrossRef] [PubMed]
- Salehi, H.; Amirpour, N.; Niapour, A.; Razavi, S. An Overview of Neural Differentiation Potential of Human Adipose Derived Stem Cells. Stem. Cell Rev. 2016, 12, 26–41. [Google Scholar] [CrossRef]
- Wang, X.; Ma, Y.; Gao, Z.; Yang, J. Human adipose-derived stem cells inhibit bioactivity of keloid fibroblasts. Stem. Cell Res. Ther. 2018, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Agha-Hosseini, F.; Jahani, M.A.; Jahani, M.; Mirzaii-Dizgah, I.; Ali-Moghaddam, K. In vitro isolation of stem cells derived from human dental pulp. Clin. Transpl. 2010, 24, E23–E28. [Google Scholar] [CrossRef]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.G.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell. Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef]
- Odabas, S.; Elçin, A.E.; Elçin, Y.M. Isolation and Characterization of Mesenchymal Stem Cells. In Bone Marrow and Stem Cell Transplantation; Beksaç, M., Ed.; Springer: New York, NY, USA, 2014; pp. 47–63. [Google Scholar] [CrossRef]
- Rebelatto, C.K.; Aguiar, A.M.; Moretao, M.P.; Senegaglia, A.C.; Hansen, P.; Barchiki, F.; Oliveira, J.; Martins, J.; Kuligovski, C.; Mansur, F.; et al. Dissimilar differentiation of mesenchymal stem cells from bone marrow, umbilical cord blood, and adipose tissue. Exp. Biol. Med. 2008, 233, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bhattacharyya, S.; Rattan, V. Effect of uncontrolled freezing on biological characteristics of human dental pulp stem cells. Cell Tissue Bank 2015, 16, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Struys, T.; Moreels, M.; Martens, W.; Donders, R.; Wolfs, E.; Lambrichts, I. Ultrastructural and immunocytochemical analysis of multilineage differentiated human dental pulp- and umbilical cord-derived mesenchymal stem cells. Cells Tissues Organs 2011, 193, 366–378. [Google Scholar] [CrossRef]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Oktar, P.A.; Yildirim, S.; Balci, D.; Can, A. Continual expression throughout the cell cycle and downregulation upon adipogenic differentiation makes nucleostemin a vital human MSC proliferation marker. Stem Cell Rev. 2011, 7, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.; Feisst, V.; Ranjard, L.; Loomes, K.; Damani, T.; Jackson-Patel, V.; Locke, M.; Sheppard, H.; Narayan, P.; Dunbar, P.R. Visualization and Quantification of Mesenchymal Cell Adipogenic Differentiation Potential with a Lineage Specific Marker. J. Vis. Exp. JoVE 2018. [Google Scholar] [CrossRef] [Green Version]
- Bowers, R.R.; Kim, J.W.; Otto, T.C.; Lane, M.D. Stable stem cell commitment to the adipocyte lineage by inhibition of DNA methylation: Role of the BMP-4 gene. Proc. Natl. Acad. Sci. USA 2006, 103, 13022–13027. [Google Scholar] [CrossRef] [Green Version]
- Abdelwahab, S.A.; Owada, Y.; Kitanaka, N.; Adida, A.; Sakagami, H.; Ono, M.; Watanabe, M.; Spener, F.; Kondo, H. Enhanced expression of adipocyte-type fatty acid binding protein in murine lymphocytes in response to dexamethasone treatment. Mol. Cell. Biochem. 2007, 299, 99–107. [Google Scholar] [CrossRef]
- Lee, D.; Wada, K.; Taniguchi, Y.; Al-Shareef, H.; Masuda, T.; Usami, Y.; Aikawa, T.; Okura, M.; Kamisaki, Y.; Kogo, M. Expression of fatty acid binding protein 4 is involved in the cell growth of oral squamous cell carcinoma. Oncol. Rep. 2014, 31, 1116–1120. [Google Scholar] [CrossRef] [Green Version]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar]
- Wafer, R.; Tandon, P.; Minchin, J.E.N. The Role of Peroxisome Proliferator-Activated Receptor Gamma (PPARG) in Adipogenesis: Applying Knowledge from the Fish Aquaculture Industry to Biomedical Research. Front. Endocrinol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.; Ong, J.M.; Clemens, T.L.; Kern, P.A. 1,25-Dihydroxyvitamin D induces lipoprotein lipase expression in 3T3-L1 cells in association with adipocyte differentiation. Endocrinology 1996, 137, 1540–1544. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, X.; Sun, L.; Guo, W.; Tian, W. Potential of human dental stem cells in repairing the complete transection of rat spinal cord. J. Neural Eng. 2017, 14, 026005. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, B.; Kumar, N.; Sreenivasa Rao, D.P.; Baadhe, R.R. Role of signaling pathways in mesenchymal stem cell differentiation. Curr. Stem. Cell. Res. Ther. 2014, 9, 508–512. [Google Scholar] [CrossRef]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of adipogenesis by Wnt signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef]
- Hu, F.; Xu, P.; Sun, B.; Teng, G.; Xiao, Z. Deep sequencing reveals complex mechanisms of microRNA regulation during retinoic acid-induced neuronal differentiation of mesenchymal stem cells. Genomics 2017. [Google Scholar] [CrossRef]
- Ichida, F.; Nishimura, R.; Hata, K.; Matsubara, T.; Ikeda, F.; Hisada, K.; Yatani, H.; Cao, X.; Komori, T.; Yamaguchi, A.; et al. Reciprocal roles of MSX2 in regulation of osteoblast and adipocyte differentiation. J. Biol. Chem. 2004, 279, 34015–34022. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, M.F. Adipogenic Differentiation of Human Mesenchymal Stem Cells. U.S. Patent No. 5,827,740, 27 October 1998. [Google Scholar]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. Roy. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pathan, M.; Keerthikumar, S.; Ang, C.S.; Gangoda, L.; Quek, C.Y.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef] [PubMed]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. g: Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic Acids Res. 2016, 44, W83–W89. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Population | Passage | Cell Culture Media | n | Mean (%) | Median (%) | SD (%) | Minimum Value (%) | Maximum Value (%) |
|---|---|---|---|---|---|---|---|---|
| DPSCs | P4 | M1 | 40 | 0 | 0 | 0 | 0 | 0 |
| M2 | 40 | 0 | 0 | 0 | 0 | 0 | ||
| ADSCs | P4 | M1 | 40 | 49.2 | 50 | 18.3 | 20 | 100 |
| M2 | 40 | 48.5 | 47.4 | 11.9 | 24 | 78.3 |
| Gene | Expression in ADSCs | Expression in DPSCs | log2-fold-change | p-Value | padj. | |
|---|---|---|---|---|---|---|
| WNT Pathway | JUN | 12,495.79 | 28,397.18 | 1.14 | 0.020 | 0.08 |
| CCND1 | 30,864.96 | 106,032.86 | 1.70 | 0.001 | 7.5 × 10−3 | |
| DKK1 | 8539.62 | 2376.18 | −1.68 | 0.019 | 0.08 | |
| TCF4 | 9071.10 | 2716.28 | −1.68 | 2.09 × 10−4 | 0.002 | |
| MMP7 | 0.52 | 6.15 | 2.63 | 0.015 | 0.07 | |
| WNT10B | 15.34 | 44.36 | 1.42 | 0.028 | 0.10 | |
| BMP Pathway | ID1 | 13,706.57 | 4008.96 | −1.68 | 0.002 | 0.02 |
| ID2 | 21,168.96 | 2332.81 | −2.98 | 1.11 × 10−6 | 2.67 × 10−5 | |
| ID3 | 36,749.25 | 4789.64 | −2.82 | 1.35 × 10−8 | 5.42 × 10−7 | |
| MSX2 | 348.26 | 4816.06 | 3.40 | 6.04 × 10−6 | 1.2 × 10−4 | |
| NOG | 101.84 | 530.68 | 2.31 | 9.24 × 10−8 | 2.94 × 10−6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fracaro, L.; Senegaglia, A.C.; Herai, R.H.; Leitolis, A.; Boldrini-Leite, L.M.; Rebelatto, C.L.K.; Travers, P.J.; Brofman, P.R.S.; Correa, A. The Expression Profile of Dental Pulp-Derived Stromal Cells Supports Their Limited Capacity to Differentiate into Adipogenic Cells. Int. J. Mol. Sci. 2020, 21, 2753. https://doi.org/10.3390/ijms21082753
Fracaro L, Senegaglia AC, Herai RH, Leitolis A, Boldrini-Leite LM, Rebelatto CLK, Travers PJ, Brofman PRS, Correa A. The Expression Profile of Dental Pulp-Derived Stromal Cells Supports Their Limited Capacity to Differentiate into Adipogenic Cells. International Journal of Molecular Sciences. 2020; 21(8):2753. https://doi.org/10.3390/ijms21082753
Chicago/Turabian StyleFracaro, Letícia, Alexandra C. Senegaglia, Roberto H. Herai, Amanda Leitolis, Lidiane M. Boldrini-Leite, Carmen L. K. Rebelatto, Paul J. Travers, Paulo R. S. Brofman, and Alejandro Correa. 2020. "The Expression Profile of Dental Pulp-Derived Stromal Cells Supports Their Limited Capacity to Differentiate into Adipogenic Cells" International Journal of Molecular Sciences 21, no. 8: 2753. https://doi.org/10.3390/ijms21082753
APA StyleFracaro, L., Senegaglia, A. C., Herai, R. H., Leitolis, A., Boldrini-Leite, L. M., Rebelatto, C. L. K., Travers, P. J., Brofman, P. R. S., & Correa, A. (2020). The Expression Profile of Dental Pulp-Derived Stromal Cells Supports Their Limited Capacity to Differentiate into Adipogenic Cells. International Journal of Molecular Sciences, 21(8), 2753. https://doi.org/10.3390/ijms21082753