Clinical Spectrum of KCNA1 Mutations: New Insights into Episodic Ataxia and Epilepsy Comorbidity


Abstract
:1. Introduction
2. Episodic Ataxia
3. KCNA1 Structure and Regulation
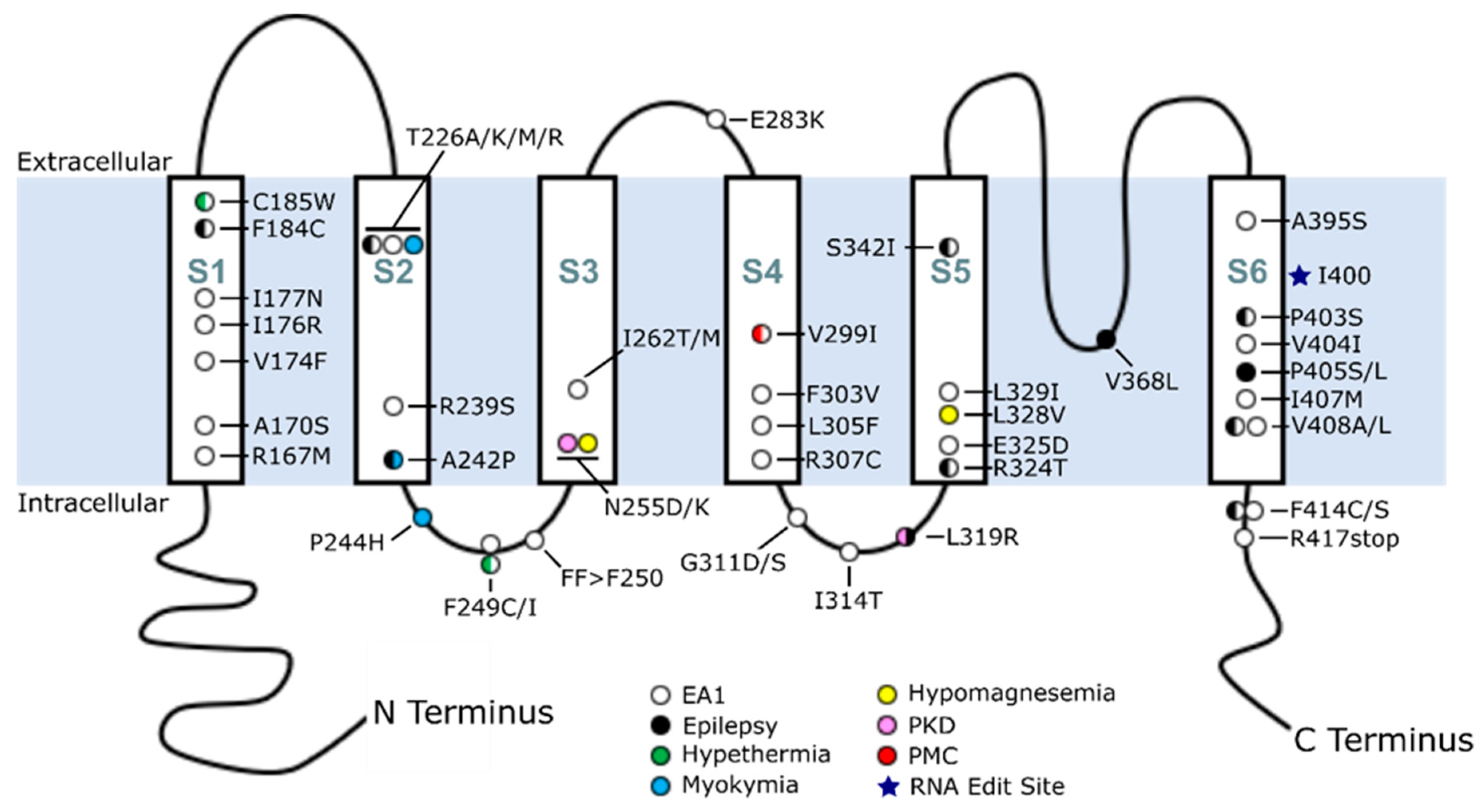
4. Overview of KCNA1 Mutations in EA1 and Disease
5. Phenotypic Variability Associated with KCNA1 Mutations
5.1. EA1
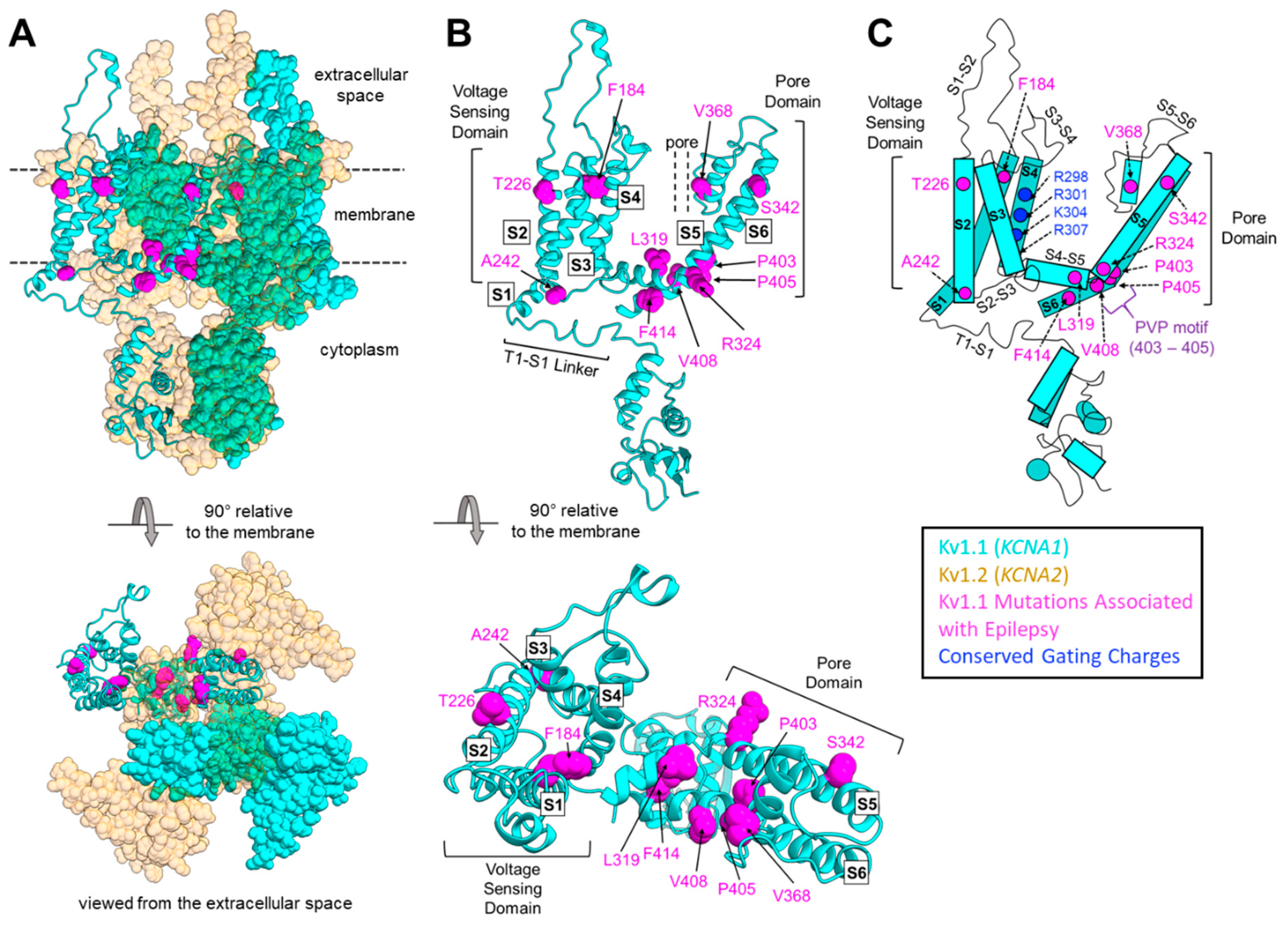
5.2. Epilepsy, Seizures, and Epileptic Encephalopathies
5.3. Hypomagnesemia
5.4. Paroxysmal Kinesigenic Dyskinesia
6. Understanding KCNA1 Mutations Using Animal Models
6.1. V408A/+ Mouse Model
6.2. Megencephaly Mouse Model
6.3. Kcna1 Global Knockout Mouse Model
6.4. Kcna1 Neuron-Specific Conditional Knockout Mouse Model
6.5. S309T Rat Model
7. Genetic Modifiers Add to Genotype–Phenotype Complexity
8. Non-Coding and Benign KCNA1 Mutations
9. Unanswered Questions and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EA1 | Episodic Ataxia Type 1 |
| SNP | Single nucleotide polymorphism |
| TM | Transmembrane |
| CNS | Central nervous system |
| aa | Amino acid |
| vdW | van der Waals |
| PVP | Proline-Valine-Proline motif |
| IL | Intracellular Linker |
| EL | Extracellular Linker |
| PKD | Paroxysmal Kinesigenic Dyskinesia |
| EE | Epileptic Encephalopathy |
| PMC | Paradoxical Myotonic Congenita |
| DD | Developmental Delay |
| ID | Intellectual Disability |
| PDD | Pervasive Developmental Disorder |
| CNV | Copy number variant |
| SMEI | Severe myoclonic epilepsy of infancy |
| TE | Tonic extension |
References
- Choi, K.-D.; Choi, J.-H. Episodic Ataxias: Clinical and Genetic Features. J. Mov. Disord. 2016, 9, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Xu, Z.; Wu, Y.; Tan, E.-K. Paroxysmal movement disorders: Recent advances and proposal of a classification system. Park. Relat. Disord. 2019, 59, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Méneret, A.; Roze, E. Paroxysmal movement disorders: An update. Mov. Disord. 2016, 172, 433–445. [Google Scholar] [CrossRef]
- Sanger, T.D.; Delgado, M.R.; Gaebler-Spira, D.; Hallett, M.; Mink, J.W. Classification and Definition of Disorders Causing Hypertonia in Childhood. Pediatrics 2003, 111, e89–e97. [Google Scholar] [CrossRef] [Green Version]
- Damak, M.; Riant, F.; Boukobza, M.; Tournier-Lasserve, E.; Bousser, M.G.; Vahedi, K. Late onset hereditary episodic ataxia. J. Neurol. Neurosurg. Psychiatry 2009, 80, 566–568. [Google Scholar] [CrossRef]
- Escayg, A.; De Waard, M.; Lee, D.D.; Bichet, D.; Wolf, P.; Mayer, T.; Johnston, J.; Baloh, R.; Sander, T.; Meisler, M.H. Coding and Noncoding Variation of the Human Calcium-Channel b4-Subunit Gene CACNB4 in Patients with Idiopathic Generalized Epilepsy and Episodic Ataxia. Am. J. Hum. Genet. 2000, 66, 1531–1539. [Google Scholar] [CrossRef] [Green Version]
- Graves, T.D.; Rajakulendran, S.; Zuberi, S.M.; Morris, F.H.R.; Schorge, F.S.; Hanna, M.G.; Kullmann, F.D.M. Nongenetic factors influence severity of episodic ataxia type 1 in monozygotic twins. Neurology 2010, 75, 367–372. [Google Scholar] [CrossRef]
- Tomlinson, S.E.; Rajakulendran, S.; Tan, S.V.; Graves, T.D.; Bamiou, D.-E.; Labrum, R.W.; Burke, D.; Sue, C.M.; Giunti, P.; Schorge, S.; et al. Clinical, genetic, neurophysiological and functional study of new mutations in episodic ataxia type 1. Mov. Disord. 2013, 84, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Coutelier, M.; Coarelli, G.; Monin, M.-L.; Konop, J.; Davoine, C.-S.; Tesson, C.; Valter, R.; Anheim, M.; Behin, A.; Castelnovo, G.; et al. A panel study on patients with dominant cerebellar ataxia highlights the frequency of channelopathies. Brain 2017, 140, 1579–1594. [Google Scholar] [CrossRef]
- Brunt, E.R.P.; Van Weerden, T.W. Familial paroxysmal kinesigenic ataxia and continuous myokymia. Brain 1990, 113, 1361–1382. [Google Scholar] [CrossRef]
- Browne, D.L.; Gancher, S.T.; Nutt, J.G.; Brunt, E.R.P.; Smith, E.A.; Kramer, P.; Litt, M. Episodic ataxia/myokymia syndrome is associated with point mutations in the human potassium channel gene, KCNA1. Nat. Genet. 1994, 8, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, H.; Mol, G.J.; Van Der Vlies, P.; Stulp, R.; Verlind, E.; Mantel, G.; Averyanov, Y.; Hofstra, R.M.; Buys, C.H.C. Three novel KCNA1 mutations in episodic ataxia type I families. Hum. Genet. 1998, 102, 464–466. [Google Scholar] [CrossRef] [PubMed]
- Imbrici, P.; D’Adamo, M.C.; Kullmann, D.M.; Pessia, M. Episodic ataxia type 1 mutations in the KCNA1 gene impair the fast inactivation properties of the human potassium channels Kv1.4-1.1/Kvβ1.1 and Kv1.4-1.1/Kvβ1.2. Eur. J. Neurosci. 2006, 24, 3073–3083. [Google Scholar] [CrossRef] [PubMed]
- Van Dyke, D. Hereditary Myokymia and Periodic Ataxia. J. Neurol. Sci. 1975, 25, 109–118. [Google Scholar] [CrossRef]
- D’adamo, M.C.; Gallenmuller, C.; Servettini, I.; Hartl, E.; Tucker, S.J.; Arning, L.; Biskup, S.; Grottesi, A.; Guglielmi, L.; Imbrici, P.; et al. Novel phenotype associated with a mutation in the KCNA1(Kv1.1) gene. Front. Physiol. 2015, 5, 525. [Google Scholar] [CrossRef] [Green Version]
- Graves, T.D.; Cha, Y.-H.; Hahn, A.F.; Barohn, R.; Salajegheh, M.K.; Griggs, R.C.; Bundy, B.N.; Jen, J.C.; Baloh, R.W.; Hanna, M.G. Episodic ataxia type 1: Clinical characterization, quality of life and genotype-phenotype correlation. Brain 2014, 137, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Comu, S.; Giuliani, M.; Narayanan, V. Episodic Ataxia and A New Mutation of Myokymia Syndrome: Potassium Channel Gene Kv1. 1. Ann. Neurol. 1996, 40, 684–687. [Google Scholar] [CrossRef]
- Chen, H.; Von Hehn, C.; Kaczmarek, L.K.; Ment, L.R.; Pober, B.R.; Hisama, F.M. Functional analysis of a novel potassium channel (KCNA1) mutation in hereditary myokymia. Neurogenetics 2007, 8, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Brownstein, C.A.; Beggs, A.H.; Rodan, L.; Shi, J.; Towne, M.C.; Pelletier, R.; Cao, S.; Rosenberg, P.A.; Urion, D.K.; Picker, J.; et al. Clinical heterogeneity associated with KCNA1 mutations include cataplexy and nonataxic presentations. Neurogenetics 2016, 17, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Zuberi, S.M.; Eunson, L.H.; Spauschus, A.; De Silva, R.; Tolmie, J.; Wood, N.W.; Mcwilliam, R.C.; Stephenson, J.P.B.; Kullmann, D.M.; Hanna, M.G. A novel mutation in the human voltage-gated potassium channel gene (Kv1.1) associates with episodic ataxia type 1 and sometimes with partial epilepsy. Brain 1999, 122, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Eunson, L.H.; Rea, R.; Zuberi, S.M.; Youroukos, S.; Panayiotopoulos, C.P.; Liguori, R.; Avoni, P.; Mcwilliam, R.C.; Stephenson, J.B.P.; Hanna, M.G.; et al. Clinical, Genetic, and Expression Studies of Mutations in the Potassium Channel Gene KCNA1 Reveal New Phenotypic Variability. Ann. Neurol. 2000, 48, 647–656. [Google Scholar] [CrossRef]
- Mestre, T.A.; Manole, A.; Macdonald, H.; Riazi, S.; Kraeva, N.; Hanna, M.G.; Lang, A.E.; Männikkö, R.; Yoon, G. A novel KCNA1 mutation in a family with episodic ataxia and malignant hyperthermia. Neurogenetics 2016, 17, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Shook, S.J.; Mamsa, H.; Jen, J.C.; Baloh, R.W.; Zhou, L. Novel mutation in KCNA1 causes episodic ataxia with paroxysmal dyspnea. Muscle Nerve 2008, 37, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Zima, L.; Ceulemans, S.; Reiner, G.; Galosi, S.; Chen, D.; Sahagian, M.; Haas, R.H.; Hyland, K.; Friedman, J. Paroxysmal motor disorders: Expanding phenotypes lead to coalescing genotypes. Ann. Clin. Transl. Neurol. 2018, 5, 996–1010. [Google Scholar] [CrossRef]
- Van der Wijst, J.; Glaudemans, B.; Venselaar, H.; Nair, A.V.; Forst, A.-L.; Hoenderop, J.G.J.; Bindels, R.J.M. Functional Analysis of the Kv1.1 N255D Mutation Associated with Autosomal Dominant Hypomagnesemia. J. Biol. Chem. 2010, 285, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Glaudemans, B.; Van Der Wijst, J.; Scola, R.H.; Lorenzoni, P.J.; Heister, A.; Van Der Kemp, A.W.; Knoers, N.V.; Hoenderop, J.G.; Bindels, R.J. A missense mutation in the Kv1.1 voltage-gated potassium channel-encoding gene KCNA1 is linked to human autosomal dominant hypomagnesemia. J. Clin. Investig. 2009, 119, 936–942. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.-M.; Lin, J.-H.; Cao, L.; Zhang, T.-M.; Zeng, S.; Zhang, K.-L.; Tian, W.-T.; Hu, Z.-M.; Li, N.; Wang, J.-L.; et al. Familial paroxysmal kinesigenic dyskinesia is associated with mutations in the KCNA1 gene. Hum. Mol. Genet. 2018, 27, 625–637. [Google Scholar] [CrossRef]
- Zhu, J.; Alsaber, R.; Zhao, J.; Ribeiro-Hurley, E.; Thornhill, W.B. Characterization of the Kv1.1 I262T and S342I mutations associated with episodic ataxia 1 with distinct phenotypes. Arch. Biochem. Biophys. 2012, 524, 99–105. [Google Scholar] [CrossRef]
- Klein, A.; Boltshauser, E.; Jen, J.; Baloh, R. Episodic ataxia type 1 with distal weakness: A novel manifestation of a potassium channelopathy. Neuropediatrics 2004, 35, 147–149. [Google Scholar]
- Lassche, S.; Lainez, S.; Bloem, B.R.; van de Warrenburg, B.P.C.; Hofmeijer, J.; Lemmink, H.H.; Hoenderop, J.G.J.; Bindels, R.J.M.; Drost, G. A novel KCNA1 mutation causing episodic ataxia type I. Muscle Nerve 2014, 50, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Imbrici, P.; Altamura, C.; Gualandi, F.; Mangiatordi, G.F.; Neri, M.; De Maria, G.; Ferlini, A.; Padovani, A.; D’Adamo, M.C.; Nicolotti, O.; et al. A novel KCNA1 mutation in a patient with paroxysmal ataxia, myokymia, painful contractures and metabolic dysfunctions. Mol. Cell. Neurosci. 2017, 83, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Rajakulendran, S.; Tan, S.; Matthews, E.; Tomlinson, S.; Labrum, R.; Sud, R.; Kullmann, D.M.; Schorge, S.; Hanna, M.G. A patient with episodic ataxia and paramyotonia congenita due to mutations in KCNA1 and SCNA4. Neurology 2009, 73, 993–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.; Bove, C.; Silvestri, G.; Mantuano, E.; Modoni, A.; Veneziano, L.; Macchioni, L.; Hunter, T.; Hunter, G.; Pessia, M.; et al. A channelopathy mutation in the voltage-sensor discloses contributions of a conserved phenylalanine to gating properties of Kv1.1 channels and ataxia. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poujois, A.; Antoine, J.-C.; Combes, A.; Laurian Touraine, R. Chronic neuromyotonia as a phenotypic variation associated with a new mutation in the KCNA1 gene. J. Neurol. 2006, 253, 957–959. [Google Scholar] [CrossRef]
- Karalok, Z.S.; Megaro, A.; Cenciarini, M.; Guven, A.; Hasan, S.M.; Taskin, B.D.; Imbrici, P.; Ceylaner, S.; Pessia, M.; D’Adamo, M.C. Identification of a new de novo mutation underlying regressive episodic ataxia type I. Front. Neurol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Zerr, P.; Adelman, J.P.; Maylie, J. Characterization of three episodic ataxia mutations in the human Kv1.1 potassium channel. FEBS J. 1998, 431, 461–464. [Google Scholar] [CrossRef] [Green Version]
- Tristán-Clavijo, E.; Scholl, F.G.; Macaya, A.; Iglesias, G.; Rojas, A.M.; Lucas, M.; Castellano, A.; Martinez-Mir, A. Dominant-negative mutation p.Arg324Thr in KCNA1 impairs Kv1.1 channel function in episodic ataxia. Mov. Disord. 2016, 31, 1743–1748. [Google Scholar] [CrossRef]
- Jan Lubbers, W.; Brunt, E.R.P.; Scheffer, H.; Litt, M.; Stulp, R.; Browne, D.L.; van Weerden, T.W. Hereditary myokymia and paroxysmal ataxia linked to chromosome 12 is responsive to acetazolamide. J. Neurol. Neurosurg. Psychiatry 1995, 59, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Van Der Wijst, J.; Konrad, M.; Verkaart, S.A.J.; Tkaczyk, M.; Latta, F.; Altmüller, J.; Thiele, H.; Beck, B.; Schlingmann, K.P.; De Baaij, J.H.F. A de novo KCNA1 Mutation in a Patient with Tetany and Hypomagnesemia. Nephron 2018, 139, 359–366. [Google Scholar] [CrossRef]
- Knight, M.A.; Storey, E.; Mckinlay Gardner, R.J.; Hand, P.; Forrest, S.M. Identification of a novel missense mutation L329I in the episodic ataxia type 1 gene KCNA1—A challenging problem. Hum. Mutat. 2000, 16, 157. [Google Scholar] [CrossRef]
- Lee, H.; Wang, H.; Jen, J.C.; Sabatti, C.; Baloh, R.W.; Nelson, S.F. A novel mutation in KCNA1 causes episodic ataxia without myokymia. Hum. Mutat. 2004, 24, 536. [Google Scholar] [CrossRef] [PubMed]
- Verdura, E.; Fons, C.; Schlüter, A.; Ruiz, M.; Fourcade, S.; Casasnovas, C.; Castellano, A.; Pujol, A. Complete loss of KCNA1 activity causes neonatal epileptic encephalopathy and dyskinesia. J. Med. Genet. 2019, 57, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.; Golumbek, P.; Cellini, E.; Doccini, V.; Guerrini, R.; Wallgren-Pettersson, C.; Thuresson, A.C.; Gurnett, C.A. De novo KCNA1 variants in the PVP motif cause infantile epileptic encephalopathy and cognitive impairment similar to recurrent KCNA2 variants. Am. J. Med. Genet. 2018, 176A, 1748–1752. [Google Scholar] [CrossRef] [PubMed]
- Tacik, P.; Guthrie, K.J.; Strongosky, A.J.; Broderick, D.F.; Riegert-Johnson, D.L.; Tang, S.; El-Khechen, D.; Parker, A.S.; Ross, O.A.; Wszolek, Z.K. Whole-exome sequencing as a diagnostic tool in a family with episodic ataxia type 1. Mayo Clin. Proc. 2015, 90, 366–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrini, E.; Marini, C.; Mei, D.; Galuppi, A.; Cellini, E.; Pucatti, D.; Chiti, L.; Rutigliano, D.; Bianchini, C.; Virdò, S.; et al. Diagnostic Targeted Resequencing in 349 Patients with Drug-Resistant Pediatric Epilepsies Identifies Causative Mutations in 30 Different Genes. Hum. Mutat. 2017, 38, 216–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demos, M.K.; Macri, V.; Farrell, K.; Nelson, T.N.; Chapman, K.; Accili, E.; Armstrong, L. A Novel KCNA1 Mutation Associated with Global Delay and Persistent Cerebellar Dysfunction. Mov. Disord. 2009, 24, 778–782. [Google Scholar] [CrossRef]
- Imbrici, P.; Gualandi, F.; D’Adamo, M.C.; Masieri, M.T.; Cudia, P.; De Grandis, D.; Mannucci, R.; Nicoletti, I.; Tucker, S.J.; Ferlini, A.; et al. A novel KCNA1 mutation identified in an Italian family affected by episodic ataxia type 1. Neuroscience 2008, 157, 577–587. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- D’Adamo, M.C.; Hasan, S.; Guglielmi, L.; Servettini, I.; Cenciarini, M.; Catacuzzeno, L.; Franciolini, F. New insights into the pathogenesis and therapeutics of episodic ataxia type 1. Front. Cell. Neurosci. 2015, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, R.; Logette, E.; Marani, M.; Herzog, M.; Tache, V.; Scantamburlo, E.; Buchillier, V.; Markram, H. A Kinetic Map of the Homomeric Voltage-Gated Potassium Channel (Kv) Family. Front. Cell. Neurosci. 2019, 13, 358. [Google Scholar] [CrossRef] [Green Version]
- Jan, L.Y.; Jan, Y.N. Voltage-gated potassium channels and the diversity of electrical signalling. J. Physiol. 2012, 590, 2591–2599. [Google Scholar] [CrossRef] [PubMed]
- Ovsepian, S.V.; Leberre, M.; Steuber, V.; O’Leary, V.B.; Leibold, C.; Oliver Dolly, J. Distinctive role of KV1.1 subunit in the biology and functions of low threshold K+ channels with implications for neurological disease. Pharmacol. Ther. 2016, 159, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, V.E.S.; Muniz, Z.M.; Sewing, S.; Lichtinghagen, R.; Parcej, D.N.; Pongs, O.; Dolly, J.O. Antibodies Specific for Distinct Kv Subunits Unveil a HeteroOligomeric Basis for Subtypes of α-Dendrotoxin-Sensitive K+ Channels in Bovine Brain. Biochemistry 1994, 33, 1617–1623. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kunkelt, D.D.; Martin, T.M.; Schwartzkroin, P.A.; Tempel, B.L. Heteromultimeric K+ channels in terminal and juxtaparanodal regions of neurons. Lett. Nat. 1993, 365, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Parcej, D.N.; Scott, V.E.; Dolly, J.O. Oligomeric properties of alpha-dendrotoxin-sensitive potassium ion channels purified from bovine brain. Biochemistry 1992, 31, 11084–11088. [Google Scholar] [CrossRef]
- Pongs, O.; Schwarz, J.R. Ancillary Subunits Associated With Voltage-Dependent K Channels. Physiol. Rev. 2010, 90, 755–796. [Google Scholar] [CrossRef] [Green Version]
- Coleman, S.K.; Newcombe, J.; Pryke, J.; Oliver Dolly, J. Subunit Composition of Kv1 Channels in Human CNS. J. Neurochem. 1999, 73, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Robbins, C.A.; Tempel, B.L. Kv1.1 and Kv1.2: Similar channels, different seizure models. Epilepsia 2012, 53, 134–141. [Google Scholar] [CrossRef]
- Miceli, F.; Soldovieri, M.V.; Ambrosino, P.; De Maria, M.; Manocchio, L.; Medoro, A.; Taglialatela, M. Molecular pathophysiology and pharmacology of the voltage-sensing module of neuronal ion channels. Front. Cell. Neurosci. 2015, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Bezanilla, F. The Voltage Sensor in Voltage-Dependent Ion Channels. Physiol. Rev. 2000, 80, 555–592. [Google Scholar] [CrossRef]
- Bhuyan, R.; Seal, A. Conformational Dynamics of Shaker-Type Kv1.1 Ion Channel in Open, Closed, and Two Mutated States. J. Membr. Biol 2015, 248, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Rea, R.; Spauschus, A.; Eunson, L.H.; Hanna, M.G.; Kullman, D.M. Variable K+ channel subunit dysfunction in inherited mutations of KCNA1. J. Physiol. 2002, 538, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, T.; Zagotta, W.; Aldrich, R. Biophysical and molecular mechanisms of Shaker potassium channel inactivation. Science 1990, 250, 533–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Jan, Y.; Jan, L. Specification of subunit assembly by the hydrophilic amino-terminal domain of the Shaker potassium channel. Science 1992, 257, 1225–1230. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, T.; Rosenthal, J.J.C.; Holmgren, M.; Reenan, R. Control of human potassium channel inactivation by editing of a small mRNA hairpin. Nat. Struct. Mol. Biol. 2004, 11, 950–956. [Google Scholar] [CrossRef]
- Ferrick-Kiddie, E.A.; Rosenthal, J.J.C.; Ayers, G.D.; Emeson, R.B. Mutations underlying Episodic Ataxia type-1 antagonize Kv1.1 RNA editing. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Imbrici, P.; D’adamo, M.C.; Grottesi, A.; Biscarini, A.; Pessia, M. Episodic ataxia type 1 mutations affect fast inactivation of K channels by a reduction in either subunit surface expression or affinity for inactivation domain. Am. J. Physiol. Cell Physiol. 2011, 300, 1314–1322. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-H.; Fu, S.-J.; Huang, J.-J.; Tang, C.-Y. The episodic ataxia type 1 mutation I262T alters voltage-dependent gating and disrupts protein biosynthesis of human Kv1.1 potassium channels. Sci. Rep. 2015, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Kunkel, D.D.; Schwartzkroin, P.A.; Tempel, B.L. Localization of Kv1.1 and Kv1.2, Two K Channel Proteins, to Synaptic Terminals, Somata, and Dendrites in the Mouse Brain. J. Neurosci. 1994, 14, 4588–4599. [Google Scholar] [CrossRef]
- Dodson, P.D.; Forsythe, I.D. Presynaptic K+ channels: Electrifying regulators of synaptic terminal excitability. Trends Neurosci. 2004, 27, 210–217. [Google Scholar] [CrossRef]
- Chiu, S.; Zhou, L.; Zhang, C.L.; Messing, A. Analysis of potassium channel functions in mammalian axons by gene knockouts. J. Neurocytol. 1999, 28, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Smart, S.L.; Lopantsev, V.; Zhang, C.L.; Robbins, C.A.; Wang, H.; Chiu, S.Y.; Schwartzkroin, P.A.; Messing, A.; Tempel, B.L. Deletion of the K(v)1.1 Potassium channel causes epilepsy in mice. Neuron 1998, 20, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Glasscock, E.; Qian, J.; Yoo, J.W.; Noebels, J.L. Masking epilepsy by combining two epilepsy genes. Nat. Neurosci. 2007, 10, 1554–1558. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Q.; Ni, F.; Ma, J. Structure of the full-length Shaker potassium channel Kv1.2 by normal-mode-based X-ray crystallographic refinement. PNAS 2010, 107, 11352–11357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.; Hunter, T.; Hunter, G.; Pessia, M.; D’Adamo, M.C. Commentary: A channelopathy mutation in the voltage-sensor discloses contributions of a conserved phenylalanine to gating properties of Kv1.1 channels and ataxia. Front. Cell. Neurosci. 2018, 12, 174. [Google Scholar] [CrossRef]
- Petitjean, D.; Kalstrup, T.; Zhao, J.; Blunck, X.R. A Disease Mutation Causing Episodic Ataxia Type I in the S1 Links Directly to the Voltage Sensor and the Selectivity Filter in Kv Channels. J. Neurosci. 2015, 35, 12198–12206. [Google Scholar] [CrossRef] [Green Version]
- Liguori, R.; Avoni, P.; Baruzzi, A.; Stasi, D.I.; Montagna, P. Familial Continuous Motor Unit Activity and Epilepsy. Muscle Nerve 2001, 24, 630–633. [Google Scholar] [CrossRef]
- Heeroma, J.H.; Henneberger, C.; Rajakulendran, S.; Hanna, M.G.; Schorge, S.; Kullmann, D.M. Episodic ataxia type 1 mutations differentially affect neuronal excitability and transmitter release. Dis. Model. Mech. 2009, 2, 612–619. [Google Scholar] [CrossRef] [Green Version]
- Tao, X.; Lee, A.; Limapichat, W.; Dougherty, D.A.; Mackinnon, R. A Gating Charge Transfer Center in Voltage Sensors. Science 2010, 328, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, J.J.; Bezanilla, F. Control of a final gating charge transition by a hydrophobic residue in the S2 segment of a K þ channel voltage sensor. PNAS 2011, 108, 6444–6449. [Google Scholar] [CrossRef] [Green Version]
- Schwaiger, C.S.; Liin, S.I.; Elinder, F.; Lindahl, E. The conserved phenylalanine in the K+ channel voltage-sensor domain creates a barrier with unidirectional effects. Biophys. J. 2013, 104, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, K.B.; Harvey, A.S.; Archer, J.S. Epileptic encephalopathy: Use and misuse of a clinically and conceptually important concept. Epilepsia 2016, 57, 343–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbrici, P.; Grottesi, A.; D’adamo, M.C.; Mannucci, R.; Tucker, S.J.; Pessia, M. Channels Contribution of the central hydrophobic residue in the PXP motif of voltage-dependent K + channels to S6 flexibility and gating properties. Channels 2009, 3, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, H.; Kimura, Y.; Enomoto, Y.; Tsurusaki, Y.; Akahira-Azuma, M.; Kuroda, Y.; Tsuji, M.; Goto, T.; Kurosawa, K. Discordant phenotype caused by CASK mutation in siblings with NF1. Hum. Genome Var. 2019, 6, 20. [Google Scholar] [CrossRef]
- Syrbe, S.; Hedrich, U.B.; Riesch, E.; Djemie, T.; Muller, S.; Moller, R.S.; Maher, B.; Hernandez-Hernandez, L.; Synofzik, M.; Caglayan, H.S.; et al. De novo loss- or gain-of-function mutations in KCNA2 cause epileptic encephalopathy. Nat. Genet. 2015, 8, 393–399. [Google Scholar] [CrossRef]
- Liu, H.-L.; Lin, J.-C. A Set of Homology Models of Pore Loop Domain of Six Eukaryotic Voltage-Gated Potassium Channels Kv1.1-Kv1.6. Proteins Struct. Funct. Genet. 2004, 55, 558–567. [Google Scholar] [CrossRef]
- Masnada, S.; Hedrich, U.B.S.; Gardella, E.; Schubert, J.; Kaiwar, C.; Klee, E.W.; Lanpher, B.C.; Gavrilova, R.H.; Synofzik, M.; Bast, T.; et al. Clinical spectrum and genotype-phenotype associations of KCNA2-related encephalopathies. Brain 2017, 140, 2337–2354. [Google Scholar] [CrossRef] [Green Version]
- Klassen, T.L.; Bomben, V.C.; Patel, A.; Drabek, J.; Chen, T.T.; Gu, W.; Zhang, F.; Chapman, K.; Lupski, J.R.; Noebels, J.L.; et al. High-resolution molecular genomic autopsy reveals complex sudden unexpected death in epilepsy risk profile. Epilepsia 2014, 55, e6–e12. [Google Scholar] [CrossRef] [Green Version]
- Harkin, L.A.; McMahon, J.M.; Iona, X.; Dibbens, L.; Pelekanos, J.T.; Zuberi, S.M.; Sadleir, L.G.; Andermann, E.; Gill, D.; Farrell, K.; et al. The spectrum of SCN1A-related infantile epileptic encephalopathies. Brain 2007, 130, 843–852. [Google Scholar] [CrossRef]
- Skluzacek, J.V.; Watts, K.P.; Parsy, O.; Wical, B.; Camfield, P. Dravet syndrome and parent associations: The IDEA League experience with comorbid conditions, mortality, management, adaptation, and grief. Epilepsia 2011, 52, 95–101. [Google Scholar] [CrossRef]
- Streit, A.K.; Derst, C.; Wegner, S.; Heinemann, U.; Zahn, R.K.; Decher, N. RNA editing of Kv1.1 channels may account for reduced ictogenic potential of 4-aminopyridine in chronic epileptic rats. Epilepsia 2011, 52, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Ritchie, G.; Kerstan, D.; Sub Kang, H.; Cole, D.E.C.; Quamme, G.A. Magnesium Transport in the Renal Distal Convoluted Tubule. Physiol. Rev. 2001, 81, 51–84. [Google Scholar] [CrossRef] [PubMed]
- Voets, T.; Nilius, B.; Hoefs, S.; Van Der Kemp, A.W.C.M.; Droogmans, G.; Bindels, R.J.M.; Hoenderop, J.G.J. TRPM6 Forms the Mg 2 Influx Channel Involved in Intestinal and Renal Mg 2 Absorption. J. Biol. Chem. 2004, 279, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, Q.; Purhonen, P.; Hebert, H. Structure of potassium channels. Cell. Mol. Life Sci. 2015, 72, 3677–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herson, P.S.; Virk, M.; Rustay, N.R.; Bond, C.T.; Crabbe, J.C.; Adelman, J.P.; Maylie, J. A mouse model of episodic ataxia type-1. Nat. Neurosci. 2003, 6, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, O.; Imbrici, P.; Botti, F.M.; Pettorossi, V.E.; D’Adamo, M.C.; Valentino, M.; Zammit, C.; Mora, M.; Gibertini, S.; Di Giovanni, G.; et al. Kv1.1 knock-in ataxic mice exhibit spontaneous myokymic activity exacerbated by fatigue, ischemia and low temperature. Neurobiol. Dis. 2012, 47, 310–321. [Google Scholar] [CrossRef]
- Persson, A.-S.; Klement, G.; Almgren, M.; Sahlholm, K.; Nilsson, J.; Petersson, S.; Århem, P.; Schalling, M.; Lavebratt, C. A truncated Kv1.1 protein in the brain of the megencephaly mouse: Expression and interaction. BMC Neurosci. 2005, 6, 65. [Google Scholar] [CrossRef] [Green Version]
- Petersson, S.; Persson, A.-S.; Johansen, J.E.; Ingvar, M.; Nilsson, J.; Ran Klement, G.È.; Rhem, P.A.Ê.; Schalling, M.; Lavebratt, C. Truncation of the Shaker-like voltage-gated potassium channel, Kv1.1, causes megencephaly. Eur. J. Neurosci. 2003, 18, 3231–3240. [Google Scholar] [CrossRef]
- Rae Donahue, L.; Cook, S.A.; Johnson, K.R.; Bronson, R.T.; Davissen, M.T. Megencephaly: A new mouse mutation on chromosome 6 that causes hypertrophy of the brain. Mamm. Genome 1996, 7, 871–876. [Google Scholar] [CrossRef]
- Dhaibar, H.; Gautier, N.M.; Chernyshev, O.Y.; Dominic, P.; Glasscock, E. Cardiorespiratory profiling reveals primary breathing dysfunction in Kcna1-null mice: Implications for sudden unexpected death in epilepsy. Neurobiol. Dis. 2019, 127, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Glasscock, E.; Yoo, J.W.; Chen, T.T.; Klassen, T.L.; Noebels, J.L. Kv1.1 Potassium Channel Deficiency Reveals Brain-Driven Cardiac Dysfunction as a Candidate Mechanism for Sudden Unexplained Death in Epilepsy. J. Neurosci. 2010, 30, 5167–5175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, M.; Trosclair, K.; Hamilton, K.A.; Glasscock, E. Genetic ablation or pharmacological inhibition of Kv1.1 potassium channel subunits impairs atrial repolarization in mice. Am. J. Physiol. Cell Physiol. 2019, 316, 154–161. [Google Scholar] [CrossRef]
- Moore, B.M.; Jou, C.J.; Tatalovic, M.; Kaufman, E.S.; Kline, D.D.; Kunze, D.L. The Kv1.1 null mouse, a model of sudden unexpected death in epilepsy (SUDEP). Epilepsia 2014, 55, 1808–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeone, K.A.; Matthews, S.A.; Rho, J.M.; Simeone, T.A. Ketogenic Diet Treatment Increases Longevity in Kcna1-null Mice, a Model of Sudden Unexpected Death in Epilepsy. Epilepsia 2016, 57, e178–e182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trosclair, K.; Dhaibar, H.A.; Gautier, N.M.; Mishra, V.; Glasscock, E. Neuron-specific Kv1.1 deficiency is sufficient to cause epilepsy, premature death, and cardiorespiratory dysregulation. Neurobiol. Dis. 2020, 137, 104759. [Google Scholar] [CrossRef]
- Ishida, S.; Sakamoto, Y.; Nishio, T.; Baulac, S.; Kuwamura, M.; Ohno, Y.; Takizawa, A.; Kaneko, S.; Serikawa, T.; Mashimo, T. Kcna1-mutant rats dominantly display myokymia, neuromyotonia and spontaneous epileptic seizures. Brain Res. 2012, 1435, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Diez, M.; Schweinhardt, P.; Petersson, S.; Wang, F.-H.; Lavebratt, C.; Schalling, M.; Èkfelt, T.H.; Spenger, C. MRI and in situ hybridization reveal early disturbances in brain size and gene expression in the megencephalic (mceph/mceph) mouse. Eur. J. Neurosci. 2003, 18, 3218–3230. [Google Scholar] [CrossRef]
- Simeone, K.A.; Hallgren, J.; Bockman, C.S.; Aggarwal, A.; Kansal, V.; Netzel, L.; Iyer, S.H.; Matthews, S.A.; Deodhar, M.; Oldenburg, P.J.; et al. Respiratory dysfunction progresses with age in Kcna1-null mice, a model of sudden unexpected death in epilepsy. Epilepsia 2018, 59, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Roundtree, H.M.; Simeone, T.A.; Johnson, C.; Matthews, S.A.; Samson, K.K.; Simeone, K.A. Orexin Receptor Antagonism Improves Sleep and Reduces Seizures in Kcna1-null Mice. Sleep 2016, 39, 352–368. [Google Scholar] [CrossRef] [Green Version]
- Fenoglio-Simeone, K.A.; Wilke, J.C.; Milligan, H.L.; Allen, C.N.; Rho, J.M.; Maganti, R.K. Ketogenic diet treatment abolishes seizure periodicity and improves diurnal rhythmicity in epileptic Kcna1-null mice. Epilepsia 2009, 50, 2027–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasscock, E.; Voigt, N.; Mccauley, M.D.; Sun, Q.; Li, N.; Chiang, D.Y.; Zhou, X.-B.; Molina, C.E.; Thomas, D.; Schmidt, C.; et al. Expression and function of Kv1.1 potassium channels in human atria from patients with atrial fibrillation. Basic Res. Cardiol. 2015, 110, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brew, H.M.; Gittelman, J.X.; Silverstein, R.S.; Hanks, T.D.; Demas, V.P.; Robinson, L.C.; Robbins, C.A.; McKee-Johnson, J.; Shing, Y.C.; Messing, A.; et al. Seizures and reduced life span in mice lacking the potassium channel subunit Kv1.2, but hypoexcitability and enlarged Kv1 currents in auditory neurons. J. Neurophysiol. 2007, 98, 1501–1525. [Google Scholar] [CrossRef] [Green Version]
- Gautier, N.M.; Glasscock, E. Spontaneous seizures in Kcna 1-null mice lacking voltage-gated Kv1.1 channels activate Fos expression in select limbic circuits. J. Neurochem. 2015, 135, 157–164. [Google Scholar] [CrossRef]
- Douglas, C.L.; Vyazovskiy, V.; Southard, T.; Chiu, S.Y.; Messing, A.; Tononi, G.; Cirelli, C. Sleep in Kcna2 knockout mice. BMC Biol. 2007, 5, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noebels, J.L.; Sidman, R.L. Inherited Epilepsy: Spike-Wave and Focal Motor Seizures in the Mutant Mouse Tottering. Science 1979, 204, 1334–1336. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, C.F.; Lutz, C.M.; Hawkes, R.; Frankel, W.N.; Copeland, N.G.; Jenkins, N.A. Absence Epilepsy in Tottering Mutant Mice Is Associated with Calcium Channel Defects. Cell 1996, 87, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Holth, J.K.; Bomben, V.C.; Graham Reed, J.; Inoue, T.; Younkin, L.; Younkin, S.G.; Pautler, R.G.; Botas, J.; Noebels, J.L. Tau loss attenuates neuronal network hyperexcitability in mouse and drosophila genetic models of epilepsy. J. Neurosci. 2013, 33, 1651–1659. [Google Scholar] [CrossRef]
- Wang, J.Z.; Liu, F. Microtubule-associated protein tau in development, degeneration and protection of neurons. Prog. Neurobiol. 2008, 85, 148–175. [Google Scholar] [CrossRef]
- Dixit, R.; Ross, J.L.; Goldman, Y.E.; Holzbaur, E.L.F. Differential regulation of dynein and kinesin motor proteins by tau. Science 2008, 319, 1086–1089. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.; Burnham, V.; Tedoldi, M.; Danial, N.N.; Yellen, G. BAD knockout provides metabolic seizure resistance in a genetic model of epilepsy with sudden unexplained death in epilepsy. Epilepsia 2018, 59, e1–e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, V.; Karumuri, B.K.; Gautier, N.M.; Liu, R.; Hutson, T.N.; Vanhoof-Villalba, S.L.; Vlachos, I.; Iasemidis, L.; Glasscock, E. Scn2a deletion improves survival and brain-heart dynamics in the Kcna1-null mouse model of sudden unexpected death in epilepsy (SUDEP). Hum. Mol. Genet. 2017, 26, 2091–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Mutation | Protein Domain | Clinical Diagnoses | Other Clinical Observations | Reference |
|---|---|---|---|---|
| R167M | S1 | EA1 | [8] | |
| A170S | S1 | EA1 | [9] | |
| V174F | S1 | EA1 | [10,11] | |
| I176R | S1 | EA1 | [12] | |
| I177N | S1 | EA1 | [13] | |
| F184C | S1 | EA1 + Seizures | [14] | |
| C185W | S1 | EA1 + Hyperthermia | Sleep a | [8,15,16] |
| T226A | S2 | EA1 | [12] | |
| T226M | S2 | EA1 | [17] | |
| T226K | S2 | Myokymia | [18] | |
| T226R | S2 | EA1 + Epilepsy | Respiratory b, Sleep c, DD | [19,20] |
| R239S | S2 | EA1 | [11] | |
| A242P | S2 | Neuromyotonia + Seizures | [8,21] | |
| P244H | S2–S3 IL | Myokymia | [21] | |
| F249C | S2–S3 IL | EA1 + Hyperthermia | [22] | |
| F249I | S2–S3 IL | EA1 | [11] | |
| FF>F250 | S2–S3 IL | EA1 | Respiratory d | [23,24] |
| N255D | S3 | Hypomagnesemia | [25,26] | |
| N255K | S3 | PKD | [27] | |
| I262T | S3 | EA1 | [28,29] | |
| I262M | S3 | EA1 | [30] | |
| E283K | S3–S4 EL | EA1 | [31] | |
| V299I | S4 | EA1 + PMC | [32] | |
| F303V | S4 | EA1 | [33] | |
| L305F | S4 | EA1 | [34] | |
| R307C | S4 | EA1 | [7] | |
| G311D | S4–S5 IL | EA1 | [35] | |
| G311S | S4–S5 IL | EA1 | [36] | |
| I314T | S4–S5 IL | EA1 | [19] | |
| L319R | S4–S5 IL | PKD + Seizures | [27] | |
| R324T | S5 | EA1 + Epilepsy | [37] | |
| E325D | S5 | EA1 | [38] | |
| L328V | S5 | Hypomagnesemia | [39] | |
| L329I | S5 | EA1 | [40] | |
| S342I | S5 | EA1 + Seizures | [28,41] | |
| V368L | S5–S6 pore loop | EE | Severe ID | [42] |
| A395S | S6 | EA1 | * | |
| P403S | S6 (PVP) | EA1 + Epilepsy | Respiratory e, DD, Moderate ID | [43] |
| V404I | S6 (PVP) | EA1 | Mild ID | [12,21,44] |
| P405S | S6 (PVP) | EE | DD, Macrocephaly f | [43] |
| P405L | S6 (PVP) | EE | PDD g | [43,45] |
| I407M | S6 | EA1 | [8] | |
| V408A | S6 | EA1 | [11] | |
| V408L | S6 | EA1 + Seizures | Global DD | [46] |
| F414C | C Terminus | EA1 | [47] | |
| F414S | C Terminus | EA1 + Epilepsy | [16] | |
| R417stop | C Terminus | EA1 | [21] |
| Species | Mutation | Phenotype | Reference |
|---|---|---|---|
| Mouse | V408A/+ | Stress-induced motor incoordination, acetazolamide responsive | [96] |
| Mouse | Truncation at aa 230 | Epilepsy, megalencephaly, unsteady gait | [98,99,100] |
| Mouse | Global gene knockout | Epilepsy, sleep deficits, cardiorespiratory abnormalities, sudden death | [72,73,101,102,103,104,105] |
| Mouse | Neuron-specific conditional gene knockout | Epilepsy, cardiorespiratory abnormalities, sudden death | [106] |
| Rat | S309T | Myokymia, neuromyotonia, seizures, premature death | [107] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulhus, K.; Ammerman, L.; Glasscock, E. Clinical Spectrum of KCNA1 Mutations: New Insights into Episodic Ataxia and Epilepsy Comorbidity. Int. J. Mol. Sci. 2020, 21, 2802. https://doi.org/10.3390/ijms21082802
Paulhus K, Ammerman L, Glasscock E. Clinical Spectrum of KCNA1 Mutations: New Insights into Episodic Ataxia and Epilepsy Comorbidity. International Journal of Molecular Sciences. 2020; 21(8):2802. https://doi.org/10.3390/ijms21082802
Chicago/Turabian StylePaulhus, Kelsey, Lauren Ammerman, and Edward Glasscock. 2020. "Clinical Spectrum of KCNA1 Mutations: New Insights into Episodic Ataxia and Epilepsy Comorbidity" International Journal of Molecular Sciences 21, no. 8: 2802. https://doi.org/10.3390/ijms21082802
APA StylePaulhus, K., Ammerman, L., & Glasscock, E. (2020). Clinical Spectrum of KCNA1 Mutations: New Insights into Episodic Ataxia and Epilepsy Comorbidity. International Journal of Molecular Sciences, 21(8), 2802. https://doi.org/10.3390/ijms21082802