Force-Generating Mechanism of Axonemal Dynein in Solo and Ensemble



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
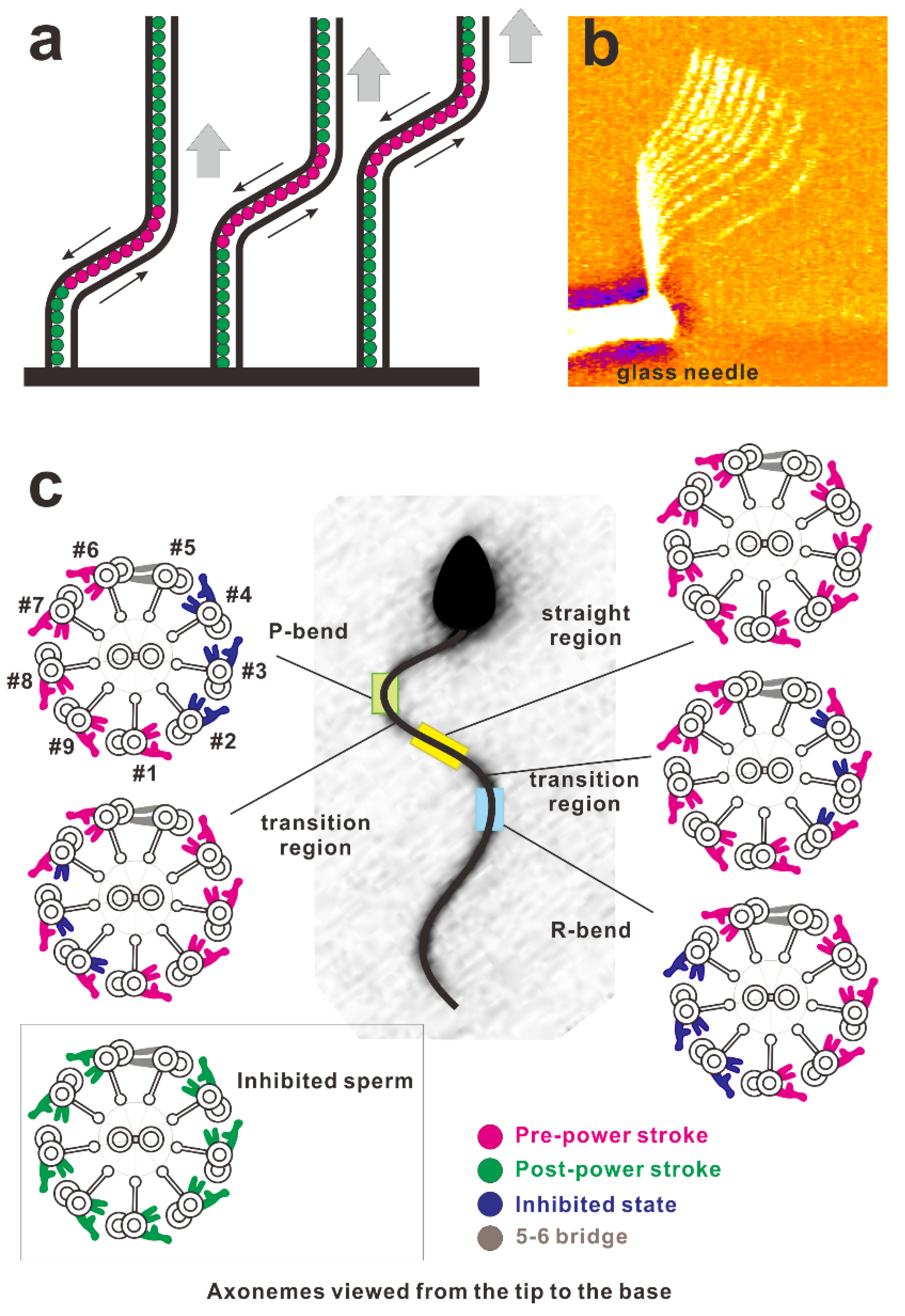
:1. Introduction
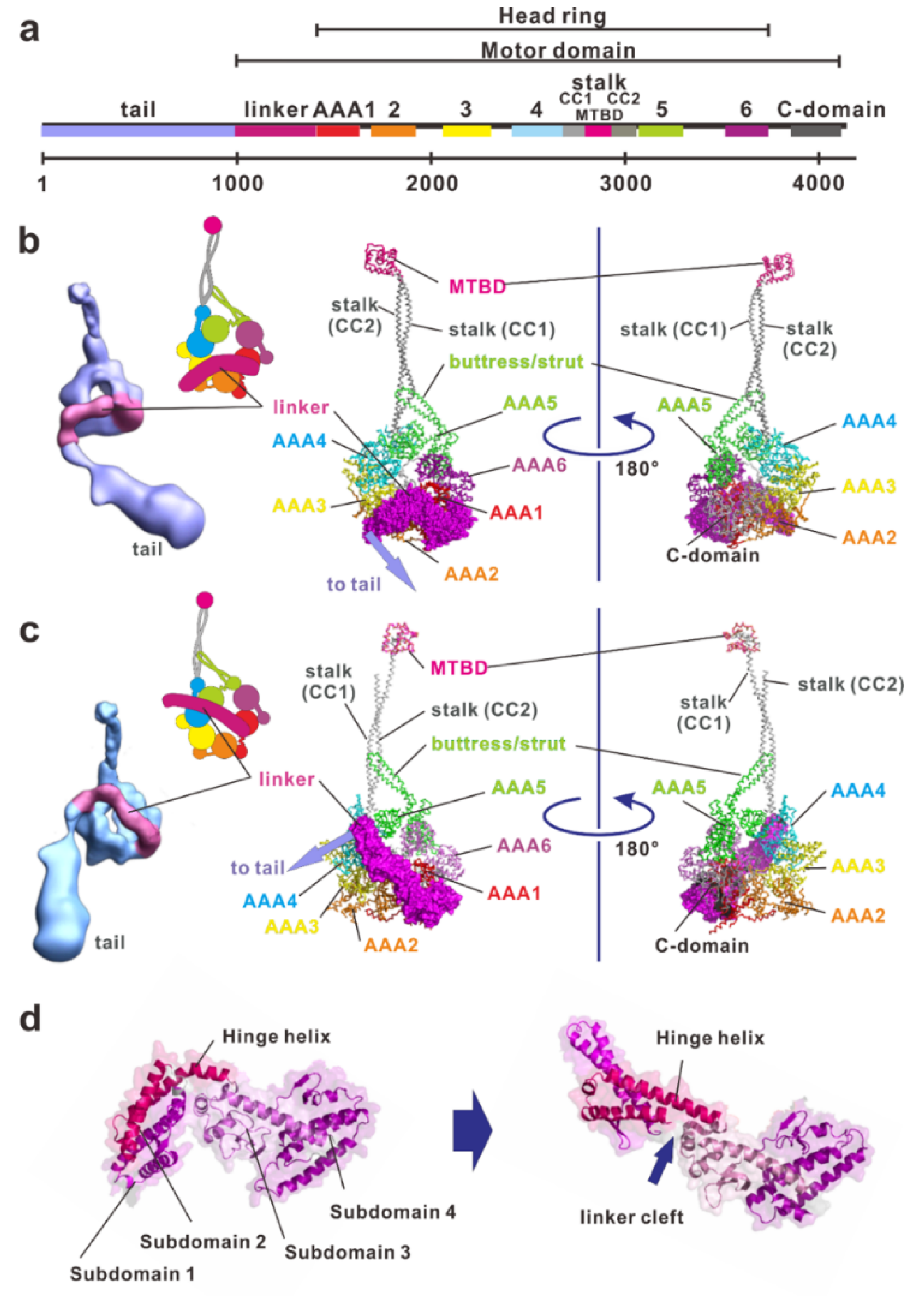
2. Overview of the Dynein Molecule Architecture
2.1. Linker
2.2. Stalk and Microtubule-Binding Domain
2.3. Buttress/Strut
2.4. Tail
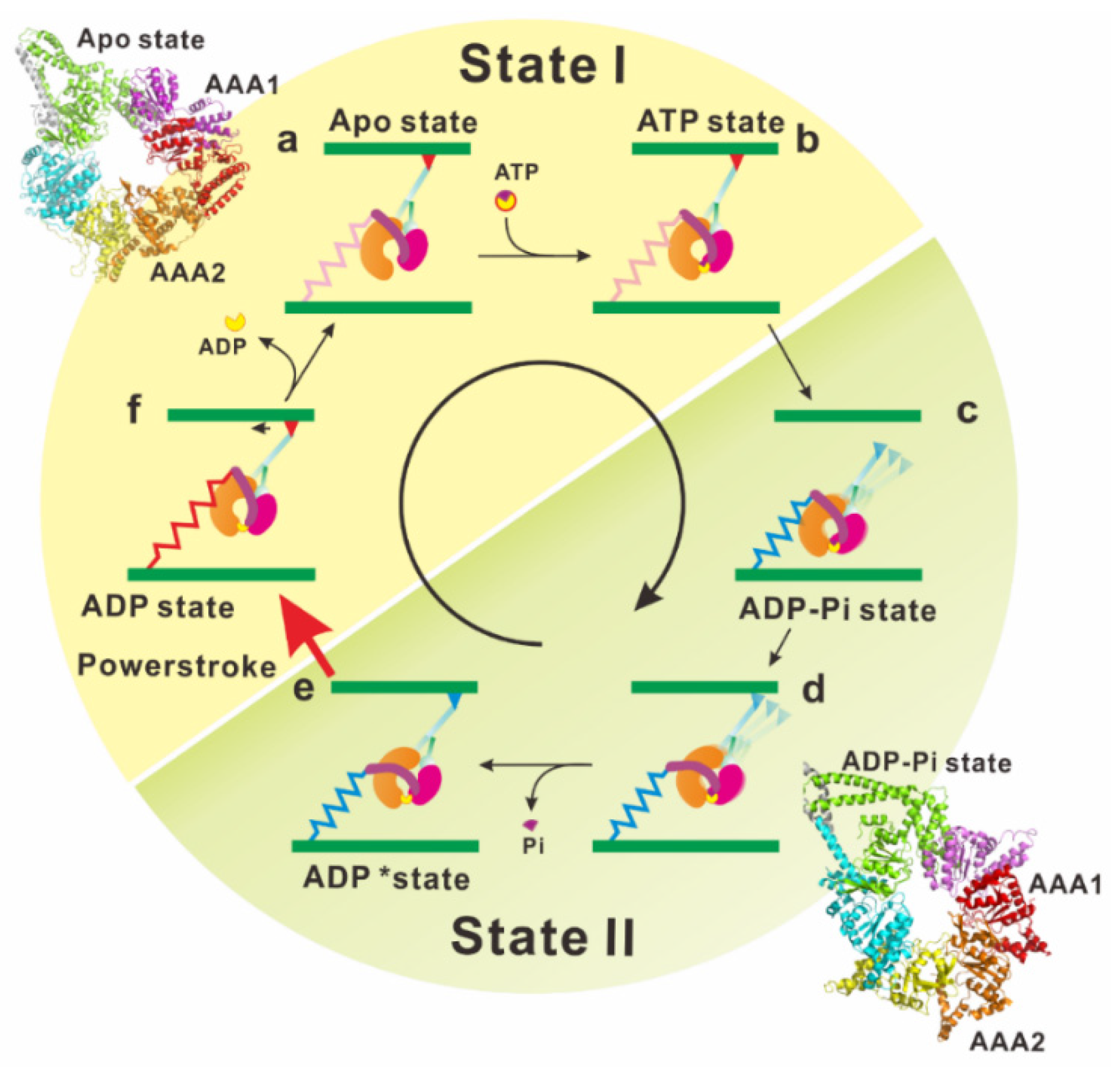
3. Force Generating Mechanism of Dynein
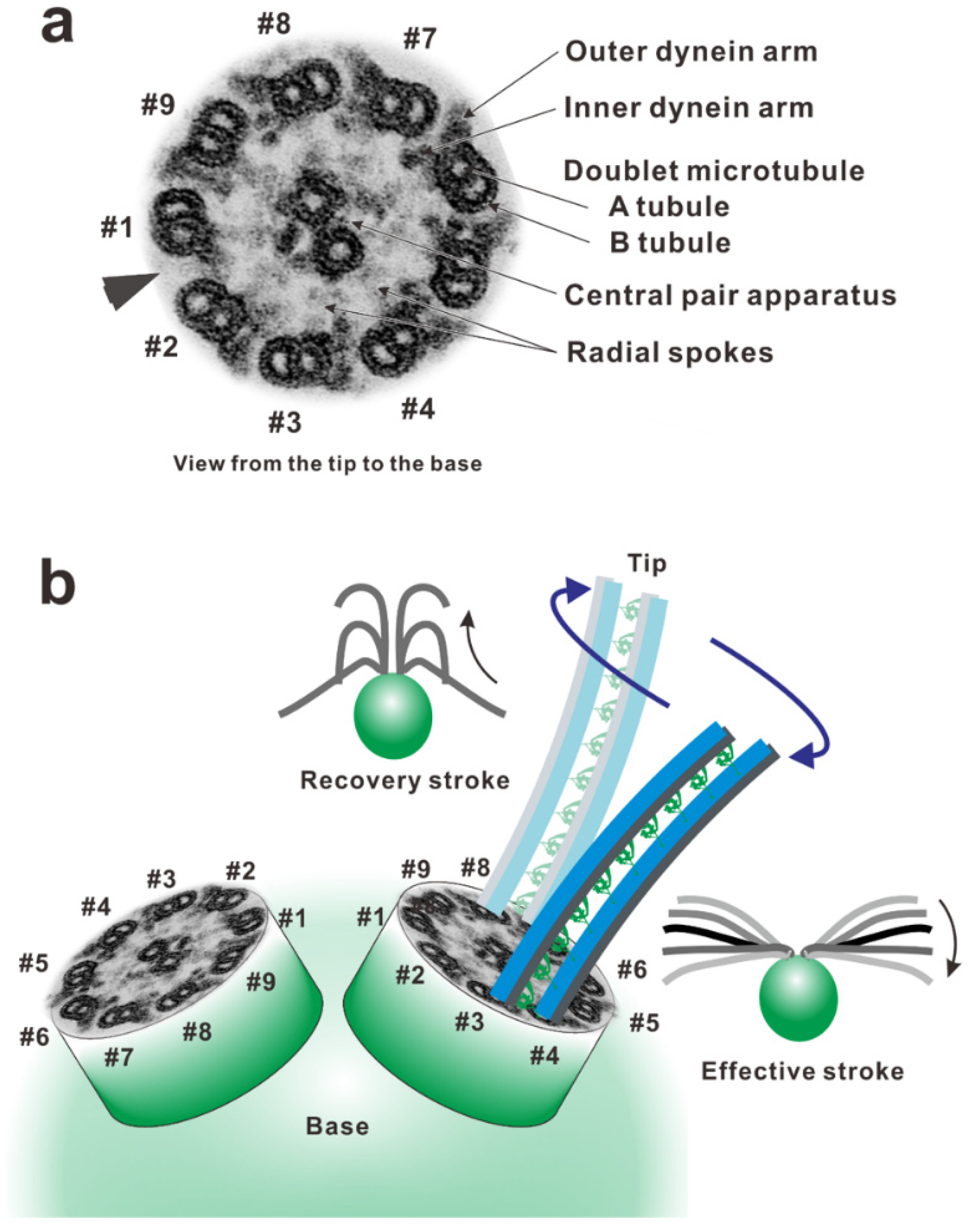
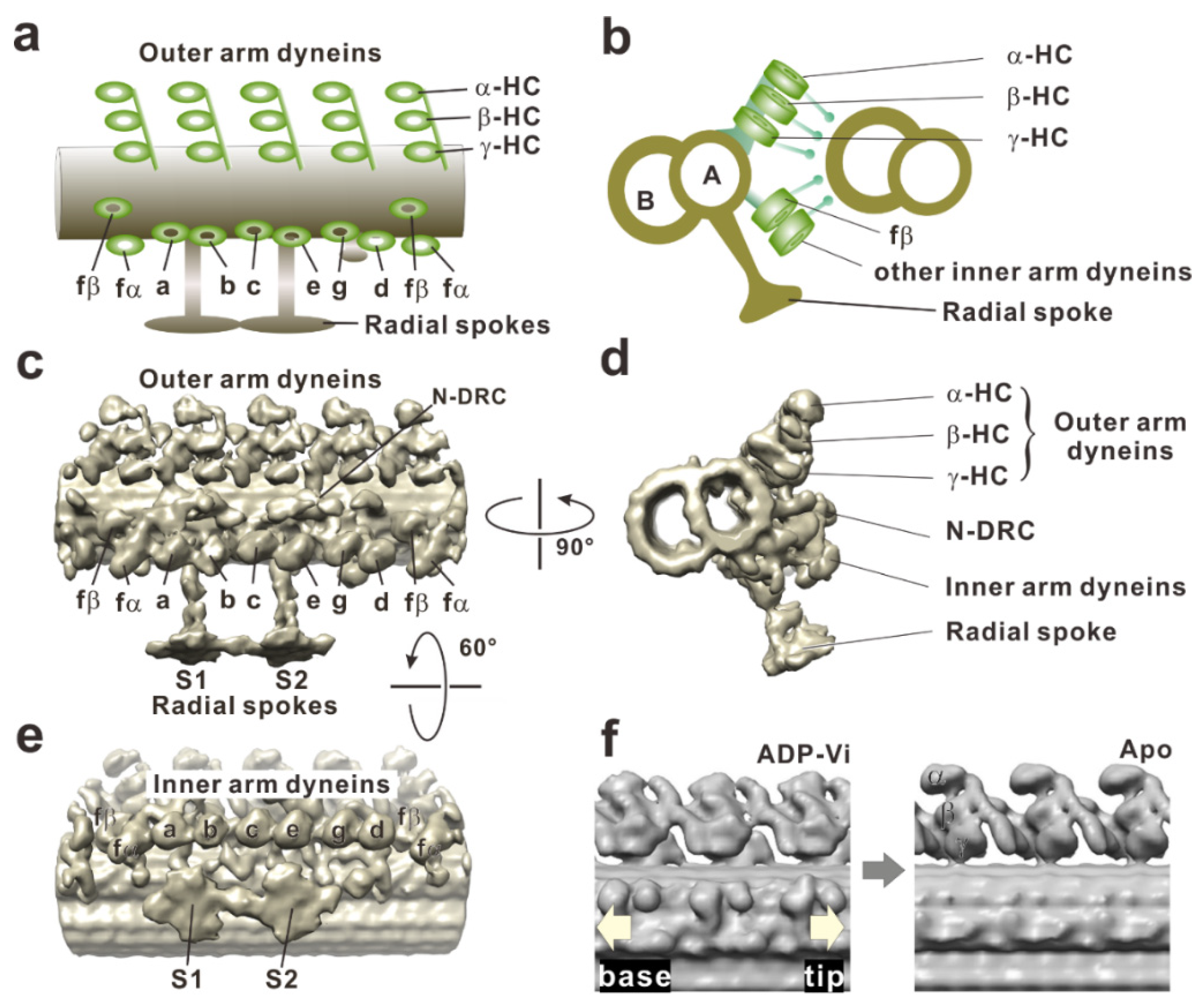
4. Force Generation of Dynein in the Axoneme
5. In the Axoneme: Conversion of the Shear between Doublets into the Bend
6. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gibbons, I.R. Studies on the protein components of cilia from tetrahymena pyriformis. Proc. Natl. Acad. Sci. USA 1963, 50, 1002–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbons, I.R.; Rowe, A.J. Dynein: A protein with adenosine triphosphatase activity from cilia. Science 1965, 149, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Afzelius, B. Electron microscopy of the sperm tail; results obtained with a new fixative. J. Biophys. Biochem. Cytol. 1959, 5, 269–278. [Google Scholar] [CrossRef] [PubMed]
- King, S.M. Axonemal dynein arms. Cold Spring Harb. Perspect. Biol. 2016, 8, a028100. [Google Scholar] [CrossRef] [PubMed]
- Kollmar, M. Fine-tuning motile cilia and flagella: Evolution of the dynein motor proteins from plants to humans at high resolution. Mol. Biol. Evol. 2016, 33, 3249–3267. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, I.R. Cilia and Flagella of Eukaryotes. J. Cell Biol. 1981, 91, 107s–124s. [Google Scholar] [CrossRef]
- Inaba, K. Molecular basis of sperm flagellar axonemes: Structural and evolutionary aspects. Ann. NY Acad. Sci. 2007, 1101, 506–526. [Google Scholar] [CrossRef]
- Mohri, H.; Inaba, K.; Ishijima, S.; Baba, S.A. Tubulin-dynein system in flagellar and ciliary movement. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 397–415. [Google Scholar] [CrossRef] [Green Version]
- Inaba, K. Calcium sensors of ciliary outer arm dynein: Functions and phylogenetic considerations for eukaryotic evolution. Cilia 2015, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Toyoshima, Y.Y. Chymotryptic digestion of Tetrahymena ciliary dynein. II. Pathway of the degradation of 22S dynein heavy chains. J. Cell Biol. 1987, 105, 897–901. [Google Scholar] [CrossRef] [Green Version]
- Pfister, K.K.; Haley, B.E.; Witman, G.B. The photoaffinity probe 8-azidoadenosine 5′-triphosphate selectively labels the heavy chain of Chlamydomonas 12S dynein. J. Biol. Chem. 1984, 259, 8499–8504. [Google Scholar] [PubMed]
- Kagami, O.; Kamiya, R. Translocation and rotation of microtubules caused by multiple species of Chlamydomonas inner-arm dynein. J. Cell Sci. 1992, 103, 653–664. [Google Scholar]
- Piperno, G.; Ramanis, Z.; Smith, E.F.; Sale, W.S. Three distinct inner dynein arms in Chlamydomonas flagella: Molecular composition and location in the axoneme. J. Cell Biol. 1990, 110, 379–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodenough, U.W.; Gebhart, B.; Mermall, V.; Mitchell, D.R.; Heuser, J.E. High-pressure liquid chromatography fractionation of Chlamydomonas dynein extracts and characterization of inner-arm dynein subunits. J. Mol. Biol. 1987, 194, 481–494. [Google Scholar] [CrossRef]
- Sale, W.S.; Fox, L.A. Isolated β-heavy chain subunit of dynein translocates microtubules in vitro. J. Cell Biol. 1988, 107, 1793–1797. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, H.; Nakayama, H. Translocation of microtubules caused by the αβ, β and γ outer arm dynein subparticles of Chlamydomonas. J. Cell Sci. 1998, 111, 1155–1164. [Google Scholar] [PubMed]
- Kotani, N.; Sakakibara, H.; Burgess, S.A.; Kojima, H.; Oiwa, K. Mechanical properties of inner-arm dynein-f (dynein I1) studied with in vitro motility assays. Biophys. J. 2007, 93, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, H.; Kojima, H.; Sakai, Y.; Katayama, E.; Oiwa, K. Inner-arm dynein c of Chlamydomonas flagella is a single-headed processive motor. Nature 1999, 400, 586–590. [Google Scholar] [CrossRef]
- Shimizu, Y.; Sakakibara, H.; Kojima, H.; Oiwa, K. Slow axonemal dynein e facilitates the motility of faster dynein c. Biophys. J. 2014, 106, 2157–2165. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, R. Functional diversity of axonemal dyneins as studied in Chlamydomonas mutants. Int. Rev. Cytol. 2002, 219, 115–155. [Google Scholar] [CrossRef]
- Yagi, T.; Minoura, I.; Fujiwara, A.; Saito, R.; Yasunaga, T.; Hirono, M.; Kamiya, R. An axonemal dynein particularly important for flagellar movement at high viscosity. Implications from a new Chlamydomonas mutant deficient in the dynein heavy chain gene DHC9. J. Biol. Chem. 2005, 280, 41412–41420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vale, R.D.; Toyoshima, Y.Y. Rotation and translocation of microtubules in vitro induced by dyneins from tetrahymena cilia. Cell 1988, 52, 459–469. [Google Scholar] [CrossRef]
- Kikushima, K.; Kamiya, R. Clockwise translocation of microtubules by flagellar inner-arm dyneins in vitro. Biophys. J. 2008, 94, 4014–4019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shingyoji, C.; Higuchi, H.; Yoshimura, M.; Katayama, E.; Yanagida, T. Dynein arms are oscillating force generators. Nature 1998, 393, 711–714. [Google Scholar] [CrossRef]
- Wais-Steider, J.; Satir, P. Effect of vanadate on gill cilia: Switching mechanism in ciliary beat. J. Supramol. Struct. 1979, 11, 339–347. [Google Scholar] [CrossRef]
- Satir, P.; Matsuoka, T. Splitting the ciliary axoneme: Implications for a “switch-point” model of dynein arm activity in ciliary motion. Cell Motil. Cytoskeleton 1989, 14, 345–358. [Google Scholar] [CrossRef]
- Woolley, D.M. Evidence for “twisted plane” undulations in golden hamster sperm tails. J. Cell Biol. 1977, 75, 851–865. [Google Scholar] [CrossRef]
- Mohri, H.; Mohri, T.; Okuno, M. Topographical relationship between the axonemal arrangement and the bend direction in starfish sperm flagella. Cell Motil. Cytoskeleton 1987, 8, 76–84. [Google Scholar] [CrossRef]
- Hoops, H.J.; Witman, G.B. Outer doublet heterogeneity reveals structural polarity related to beat direction in Chlamydomonas flagella. J. Cell Biol. 1983, 97, 902–908. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Shingyoji, C. Bending-induced switching of dynein activity in elastase-treated axonemes of sea urchin sperm--roles of Ca2+ and ADP. Cell Motil. Cytoskeleton 2009, 66, 292–301. [Google Scholar] [CrossRef]
- Hayashi, S.; Shingyoji, C. Mechanism of flagellar oscillation-bending-induced switching of dynein activity in elastase-treated axonemes of sea urchin sperm. J. Cell Sci. 2008, 121, 2833–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, R.; Shingyoji, C. Induction of beating by imposed bending or mechanical pulse in demembranated, motionless sea urchin sperm flagella at very low ATP concentrations. Cell Struct. Funct. 2007, 32, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, Y.; Shingyoji, C. Effects of imposed bending on microtubule sliding in sperm flagella. Curr. Biol. 2004, 14, 2113–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindemann, C.B.; Lesich, K.A. Flagellar and ciliary beating: The proven and the possible. J. Cell Sci. 2010, 123, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, A.P.; Cho, C.; Jin, L.; Vale, R.D. Crystal structure of the dynein motor domain. Science 2011, 331, 1159–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, A.P.; Garbarino, J.E.; Wilson-Kubalek, E.M.; Shipley, W.E.; Cho, C.; Milligan, R.A.; Vale, R.D.; Gibbons, I.R. Structure and functional role of dynein’s microtubule-binding domain. Science 2008, 322, 1691–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Gleave, E.S.; Carter, A.P. Insights into dynein motor domain function from a 3.3-Åcrystal structure. Nat. Struct. Mol. Biol. 2012, 19, 492–497. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.; Zalyte, R.; Urnavicius, L.; Carter, A.P. Structure of human cytoplasmic dynein-2 primed for its power stroke. Nature 2015, 518, 435–438. [Google Scholar] [CrossRef] [Green Version]
- Kon, T.; Oyama, T.; Shimo-Kon, R.; Imamula, K.; Shima, T.; Sutoh, K.; Kurisu, G. The 2.8 Å crystal structure of the dynein motor domain. Nature 2012, 484, 345–350. [Google Scholar] [CrossRef]
- Kon, T.; Sutoh, K.; Kurisu, G. X-ray structure of a functional full-length dynein motor domain. Nat. Struct. Mol. Biol. 2011, 18, 638–642. [Google Scholar] [CrossRef]
- Goodenough, U.W.; Heuser, J.E. Substructure of the outer dynein arm. J. Cell. Biol. 1982, 95, 798–815. [Google Scholar] [CrossRef] [PubMed]
- Goodenough, U.W.; Heuser, J.E. Outer and inner dynein arms of cilia and flagella. Cell 1985, 41, 341–342. [Google Scholar] [CrossRef]
- Goodenough, U.W.; Heuser, J.E. Substructure of inner dynein arms, radial spokes, and the central pair/projection complex of cilia and flagella. J. Cell Biol. 1985, 100, 2008–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, K.H.; Sakakibara, H.; Movassagh, T.; Oiwa, K.; Ishikawa, T. Asymmetry of inner dynein arms and inter-doublet links in Chlamydomonas flagella. J. Cell Biol. 2009, 186, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Bui, K.H.; Sakakibara, H.; Movassagh, T.; Oiwa, K.; Ishikawa, T. Molecular architecture of inner dynein arms in situ in Chlamydomonas reinhardtii flagella. J. Cell Biol. 2008, 183, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Sakakibara, H.; Oiwa, K. The architecture of outer dynein arms in situ. J. Mol. Biol. 2007, 368, 1249–1258. [Google Scholar] [CrossRef]
- Movassagh, T.; Bui, K.H.; Sakakibara, H.; Oiwa, K.; Ishikawa, T. Nucleotide-induced global conformational changes of flagellar dynein arms revealed by in situ analysis. Nat. Struct. Mol. Biol. 2010, 17, 761–767. [Google Scholar] [CrossRef]
- Lin, J.; Nicastro, D. Asymmetric distribution and spatial switching of dynein activity generates ciliary motility. Science 2018, 360, eaar1968. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Okada, K.; Raytchev, M.; Smith, M.C.; Nicastro, D. Structural mechanism of the dynein power stroke. Nat. Cell Biol. 2014, 16, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Nicastro, D.; McIntosh, J.R.; Baumeister, W. 3D structure of eukaryotic flagella in a quiescent state revealed by cryo-electron tomography. Proc. Natl. Acad. Sci. USA 2005, 102, 15889–15894. [Google Scholar] [CrossRef] [Green Version]
- Nicastro, D.; Schwartz, C.; Pierson, J.; Gaudette, R.; Porter, M.E.; McIntosh, J.R. The molecular architecture of axonemes revealed by cryoelectron tomography. Science 2006, 313, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.A.; Walker, M.L.; Sakakibara, H.; Knight, P.J.; Oiwa, K. Dynein structure and power stroke. Nature 2003, 421, 715–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.J.; Kon, T.; Knight, P.J.; Sutoh, K.; Burgess, S.A. Functions and mechanics of dynein motor proteins. Nat. Rev. Mol. Cell Biol. 2013, 14, 713–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.J.; Malkova, B.; Walker, M.L.; Sakakibara, H.; Numata, N.; Kon, T.; Ohkura, R.; Edwards, T.A.; Knight, P.J.; Sutoh, K.; et al. ATP-driven remodeling of the linker domain in the dynein motor. Structure 2012, 20, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.J.; Numata, N.; Walker, M.L.; Kato, Y.S.; Malkova, B.; Kon, T.; Ohkura, R.; Arisaka, F.; Knight, P.J.; Sutoh, K.; et al. AAA+ ring and linker swing mechanism in the dynein motor. Cell 2009, 136, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Wickstead, B.; Gull, K. Dyneins across eukaryotes: A comparative genomic analysis. Traffic 2007, 8, 1708–1721. [Google Scholar] [CrossRef] [Green Version]
- Ogura, T.; Wilkinson, A.J. AAA+ superfamily ATPases: Common structure--diverse function. Genes Cells 2001, 6, 575–597. [Google Scholar] [CrossRef]
- Asai, D.J.; Koonce, M.P. The dynein heavy chain: Structure, mechanics and evolution. Trends Cell Biol. 2001, 11, 196–202. [Google Scholar] [CrossRef]
- Gibbons, I.R.; Gibbons, B.H.; Mocz, G.; Asai, D.J. Multiple nucleotide-binding sites in the sequence of dynein β heavy chain. Nature 1991, 352, 640–643. [Google Scholar] [CrossRef]
- Ogawa, K. Four ATP-binding sites in the midregion of the β heavy chain of dynein. Nature 1991, 352, 643–645. [Google Scholar] [CrossRef]
- Koonce, M.P.; Grissom, P.M.; McIntosh, J.R. Dynein from Dictyostelium: Primary structure comparisons between a cytoplasmic motor enzyme and flagellar dynein. J. Cell Biol. 1992, 119, 1597–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikami, A.; Paschal, B.M.; Mazumdar, M.; Vallee, R.B. Molecular cloning of the retrograde transport motor cytoplasmic dynein (MAP 1C). Neuron 1993, 10, 787–796. [Google Scholar] [CrossRef]
- Kon, T.; Nishiura, M.; Ohkura, R.; Toyoshima, Y.Y.; Sutoh, K. Distinct functions of nucleotide-binding/hydrolysis sites in the four AAA modules of cytoplasmic dynein. Biochemistry 2004, 43, 11266–11274. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.; Reck-Peterson, S.L.; Vale, R.D. Regulatory ATPase sites of cytoplasmic dynein affect processivity and force generation. J. Biol. Chem. 2008, 283, 25839–25845. [Google Scholar] [CrossRef] [Green Version]
- Silvanovich, A.; Li, M.G.; Serr, M.; Mische, S.; Hays, T.S. The third P-loop domain in cytoplasmic dynein heavy chain is essential for dynein motor function and ATP-sensitive microtubule binding. Mol. Biol. Cell 2003, 14, 1355–1365. [Google Scholar] [CrossRef]
- Yagi, T. ADP-dependent microtubule translocation by flagellar inner-arm dyneins. Cell Struct. Funct. 2000, 25, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Kikushima, K.; Yagi, T.; Kamiya, R. Slow ADP-dependent acceleration of microtubule translocation produced by an axonemal dynein. FEBS Letters 2004, 563, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Shiroguchi, K.; Toyoshima, Y.Y. Regulation of monomeric dynein activity by ATP and ADP concentrations. Cell Motil. Cytoskeleton 2001, 49, 189–199. [Google Scholar] [CrossRef]
- Mocz, G.; Helms, M.K.; Jameson, D.M.; Gibbons, I.R. Probing the nucleotide binding sites of axonemal dynein with the fluorescent nucleotide analogue 2′(3′)-O-(-N-methylanthraniloyl)-adenosine 5′-triphosphate. Biochemistry 1998, 37, 9862–9869. [Google Scholar] [CrossRef]
- Kon, T.; Imamula, K.; Roberts, A.J.; Ohkura, R.; Knight, P.J.; Gibbons, I.R.; Burgess, S.A.; Sutoh, K. Helix sliding in the stalk coiled coil of dynein couples ATPase and microtubule binding. Nat. Struct Mol. Biol. 2009, 16, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, I.R.; Garbarino, J.E.; Tan, C.E.; Reck-Peterson, S.L.; Vale, R.D.; Carter, A.P. The affinity of the dynein microtubule-binding domain is modulated by the conformation of its coiled-coil stalk. J. Biol. Chem. 2005, 280, 23960–23965. [Google Scholar] [CrossRef] [Green Version]
- Lacey, S.E.; He, S.; Scheres, S.H.W.; Carter, A.P. Cryo-EM of dynein microtubule-binding domains shows how an axonemal dynein distorts the microtubule. eLife 2019, 8, e47145. [Google Scholar] [CrossRef]
- Kardon, J.R.; Vale, R.D. Regulators of the cytoplasmic dynein motor. Nat. Rev. Mol. Cell Biol. 2009, 10, 854–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reck-Peterson, S.L.; Yildiz, A.; Carter, A.P.; Gennerich, A.; Zhang, N.; Vale, R.D. Single-molecule analysis of dynein processivity and stepping behavior. Cell 2006, 126, 335–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torisawa, T.; Ichikawa, M.; Furuta, A.; Saito, K.; Oiwa, K.; Kojima, H.; Toyoshima, Y.Y.; Furuta, K. Autoinhibition and cooperative activation mechanisms of cytoplasmic dynein. Nat. Cell Biol. 2014, 16, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Toropova, K.; Zalyte, R.; Mukhopadhyay, A.G.; Mladenov, M.; Carter, A.P.; Roberts, A.J. Structure of the dynein-2 complex and its assembly with intraflagellar transport trains. Nat. Struct Mol. Biol. 2019, 26, 823–829. [Google Scholar] [CrossRef]
- Toropova, K.; Mladenov, M.; Roberts, A.J. Intraflagellar transport dynein is autoinhibited by trapping of its mechanical and track-binding elements. Nat. Struct Mol. Biol. 2017, 24, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, H.A.; Kamiya, R. Association between actin and light chains in Chlamydomonas flagellar inner-arm dyneins. Biochem. Biophys. Res. Commun. 2001, 288, 443–447. [Google Scholar] [CrossRef]
- LeDizet, M.; Piperno, G. The light chain p28 associates with a subset of inner dynein arm heavy chains in Chlamydomonas axonemes. Mol. Biol. Cell 1995, 6, 697–711. [Google Scholar] [CrossRef] [Green Version]
- Piperno, G.; Mead, K.; Shestak, W. The inner dynein arms I2 interact with a “dynein regulatory complex” in Chlamydomonas flagella. J. Cell Biol. 1992, 118, 1455–1463. [Google Scholar] [CrossRef]
- Takada, S.; Wilkerson, C.G.; Wakabayashi, K.; Kamiya, R.; Witman, G.B. The outer dynein arm-docking complex: Composition and characterization of a subunit (oda1) necessary for outer arm assembly. Mol. Biol. Cell 2002, 13, 1015–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, A.P. Crystal clear insights into how the dynein motor moves. J. Cell Sci. 2013, 126, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Carter, A.P. Review: Structure and mechanism of the dynein motor ATPase. Biopolymers 2016, 105, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Kon, T.; Mogami, T.; Ohkura, R.; Nishiura, M.; Sutoh, K. ATP hydrolysis cycle-dependent tail motions in cytoplasmic dynein. Nat. Struct Mol. Biol. 2005, 12, 513–519. [Google Scholar] [CrossRef]
- Oiwa, K.; Sakakibara, H.; Furuta, K. Electron microscopy of isolated dynein complexes and the power stroke mechanism. In Dyneins: Dynein Mechanics, Dysfunction, and Disease, 2nd ed.; King, S.M., Ed.; Academic Press: London, UK, 2018; pp. 2–35. ISBN 9780128094709. [Google Scholar]
- Allen, R.D. A reinvestigation of cross-sections of cilia. J Cell Biol 1968, 37, 825–831. [Google Scholar] [CrossRef]
- Avolio, J.; Lebduska, S.; Satir, P. Dynein arm substructure and the orientation of arm-microtubule attachments. J. Mol. Biol. 1984, 173, 389–401. [Google Scholar] [CrossRef]
- Warner, F.D.; Mitchell, D.R.; Perkins, C.R. Structural conformation of the ciliary ATPase dynein. J. Mol. Biol. 1977, 114, 367–384. [Google Scholar] [CrossRef]
- Witman, G.B.; Minervini, N. Dynein arm conformation and mechanochemical transduction in the eukaryotic flagellum. Symp. Soc. Exp. Biol. 1982, 35, 203–223. [Google Scholar] [PubMed]
- Bui, K.H.; Pigino, G.; Ishikawa, T. Three-dimensional structural analysis of eukaryotic flagella/cilia by electron cryo-tomography. J. Synchrotron Radiat. 2011, 18, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T. Axoneme structure from motile cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028076. [Google Scholar] [CrossRef] [Green Version]
- Ueno, H.; Bui, K.H.; Ishikawa, T.; Imai, Y.; Yamaguchi, T.; Ishikawa, T. Structure of dimeric axonemal dynein in cilia suggests an alternative mechanism of force generation. Cytoskeleton (Hoboken) 2014, 71, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.A.; Knight, P.J. Is the dynein motor a winch? Curr. Opin. Struct. Biol. 2004, 14, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.; Yasunaga, T.; Shingyoji, C.; Hirose, K. Dynein pulls microtubules without rotating its stalk. Proc. Natl. Acad. Sci. USA 2008, 105, 19702–19707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, H.; Shima, T.; Sutoh, K.; Walker, M.L.; Knight, P.J.; Kon, T.; Burgess, S.A. Direct observation shows superposition and large scale flexibility within cytoplasmic dynein motors moving along microtubules. Nat. Commun. 2015, 6, 8179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shingyoji, C.; Murakami, A.; Takahashi, K. Local reactivation of triton-extracted flagella by iontophoretic application of ATP. Nature 1977, 265, 269–270. [Google Scholar] [CrossRef] [PubMed]
- Okuno, M.; Hiramoto, Y. Mechanical stimulation of starfish sperm flagella. J. Exp. Biol. 1976, 65, 401–413. [Google Scholar] [PubMed]
- Lindemann, C.B.; Rikmenspoel, R. Sperm flagella: Autonomous oscillations of the contractile system. Science 1972, 175, 337–338. [Google Scholar] [CrossRef]
- Hayashibe, K.; Shingyoji, C.; Kamiya, R. Induction of temporary beating in paralyzed flagella of Chlamydomonas mutants by application of external force. Cell Motil. Cytoskeleton. 1997, 37, 232–239. [Google Scholar] [CrossRef]
- Hastie, A.T. Purification and characterization of dynein from pig tracheal cilia. Methods Enzymol. 1991, 196, 223–234. [Google Scholar] [CrossRef]
- King, S.M. Turning dyneins off bends cilia. Cytoskeleton (Hoboken) 2018, 75, 372–381. [Google Scholar] [CrossRef]
- Porter, M.E.; Sale, W.S. The 9 + 2 axoneme anchors multiple inner arm dyneins and a network of kinases and phosphatases that control motility. J. Cell Biol. 2000, 151, F37–F42. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.F.; Sale, W.S. Regulation of dynein-driven microtubule sliding by the radial spokes in flagella. Science 1992, 257, 1557–1559. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Yanagisawa, H.; Yagi, T.; Kikkawa, M. Mechanosignaling between central apparatus and radial spokes controls axonemal dynein activity. J. Cell Biol. 2014, 204, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Yagi, T.; Nishiyama, M. High hydrostatic pressure induces vigorous flagellar beating in Chlamydomonas non-motile mutants lacking the central apparatus. Sci. Rep. 2020, 10, 2072. [Google Scholar] [CrossRef] [PubMed]
- Brokaw, C.J. Molecular mechanism for oscillation in flagella and muscle. Proc. Natl. Acad. Sci. USA 1975, 72, 3102–3106. [Google Scholar] [CrossRef] [Green Version]
- Jülicher, F.; Prost, J. Cooperative molecular motors. Phys. Rev. Lett. 1995, 75, 2618–2621. [Google Scholar] [CrossRef]
- Jülicher, F.; Prost, J. Spontaneous oscillations of collective molecular motors. Phys. Rev. Lett. 1997, 78, 4510–4513. [Google Scholar] [CrossRef] [Green Version]
- Riedel-Kruse, I.H.; Hilfinger, A.; Howard, J.; Jülicher, F. How molecular motors shape the flagellar beat. HFSP Journal 2007, 1, 192–208. [Google Scholar] [CrossRef] [Green Version]
- Brokaw, C.J. Flagellar movement: A sliding filament model. Science 1972, 178, 455–462. [Google Scholar] [CrossRef]
- Brokaw, C.J. Computer simulation of flagellar movement. I. Demonstration of stable bend propagation and bend initiation by the sliding filament model. Biophys. J. 1972, 12, 564–586. [Google Scholar] [CrossRef] [Green Version]
- Sartori, P.; Geyer, V.F.; Howard, J.; Jülicher, F. Curvature regulation of the ciliary beat through axonemal twist. Phys. Rev. E. 2016, 94, 042426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindemann, C.B. A model of flagellar and ciliary functioning which uses the forces transverse to the axoneme as the regulator of dynein activation. Cell Motil. Cytoskeleton 1994, 29, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, C.B. A geometric clutch hypothesis to explain oscillations of the axoneme of cilia and flagella. J. Theor. Biol. 1994, 168, 175–189. [Google Scholar] [CrossRef]
- Lindemann, C.B. Geometric clutch model version 3: The role of the inner and outer arm dyneins in the ciliary beat. Cell Motil. Cytoskeleton 2002, 52, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Huang, J.; Yang, J.; He, D.; Wang, C.; Qi, X.; Taylor, I.A.; Liu, J.; Peng, Y.L. Structure based function-annotation of hypothetical protein MGG_01005 from Magnaporthe oryzae reveals it is the dynein light chain orthologue of dynlt1/3. Sci. Rep. 2018, 8, 3952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibusuki, R.; Shiraga, M.; Furuta, A.; Yoshio, M.; Kojima, H.; Oiwa, K.; Furuta, K. Collective motility of dynein linear arrays built on DNA nanotubes. Biochem. Biophys. Res. Commun. 2020, 523, 1014–1019. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishibashi, K.; Sakakibara, H.; Oiwa, K. Force-Generating Mechanism of Axonemal Dynein in Solo and Ensemble. Int. J. Mol. Sci. 2020, 21, 2843. https://doi.org/10.3390/ijms21082843
Ishibashi K, Sakakibara H, Oiwa K. Force-Generating Mechanism of Axonemal Dynein in Solo and Ensemble. International Journal of Molecular Sciences. 2020; 21(8):2843. https://doi.org/10.3390/ijms21082843
Chicago/Turabian StyleIshibashi, Kenta, Hitoshi Sakakibara, and Kazuhiro Oiwa. 2020. "Force-Generating Mechanism of Axonemal Dynein in Solo and Ensemble" International Journal of Molecular Sciences 21, no. 8: 2843. https://doi.org/10.3390/ijms21082843
APA StyleIshibashi, K., Sakakibara, H., & Oiwa, K. (2020). Force-Generating Mechanism of Axonemal Dynein in Solo and Ensemble. International Journal of Molecular Sciences, 21(8), 2843. https://doi.org/10.3390/ijms21082843